
沉水植物对淡水养殖池塘底泥重金属Cu污染的修复研究
2019-05-21 08:46:24原居林张雷鸣顾志敏
淡水渔业 2019年3期
刘 梅,原居林,倪 蒙,张雷鸣,顾志敏,
(1.农业部淡水渔业健康养殖重点实验室,浙江省淡水水产遗传育种重点实验室,浙江省淡水水产研究所,浙江湖州 313001;2.上海海洋大学水产与生命科学学院,上海 201306)
近年来,淡水养殖池塘重金属超标问题相关报道研究越来越多[1-4],重金属污染问题由于具有持久性、生物富集和放大作用已成为我国最重要的环境和食品安全问题之一[5-7]。Cu作为鱼类生长必需元素之一,在水产饲料中过度添加的现象非常普遍,而残饵在池塘底部堆积,造成其在表层沉积物中的含量不断升高[8-9];另外养殖过程中渔药(如硫酸铜等)的滥用,进一步导致养殖水体中重金属Cu含量不断增加。由于重金属极易吸附在水体的悬浮颗粒物中,且不能降解,因此通过各种途径进入养殖水体的重金属最终通过吸附、沉降、埋藏等过程进入到底泥中。重金属在底泥中的大量积累会形成巨大的潜在危害和风险[2,10]。许多研究显示,我国淡水池塘底泥重金属Cu含量远高于我国土壤环境质量Ⅰ级标准(35 mg/kg)几倍甚至几十倍,如贾成霞等[1]调查发现北京地区养殖塘底泥Cu含量为18~155 mg/kg,而施沁璇等[3]2014年调查发现湖州市养殖塘底泥Cu含量为10.50~98.20 mg/kg,因此池塘底泥重金属Cu超标问题不容忽视。
目前对土壤重金属污染的治理研究大多以植物修复为主,通过植物对重金属的吸收、累积、转移来达到治理目的[11-12]。沉水植物因全株都生活在水体中,茎、叶的表皮都与根一样具有吸收作用,这种结构对重金属元素的浓缩富集都有很强的作用。而且植物生长过程中需要少量的Cu,Cu是其正常生命活动所必需的微量元素之一,是一些酶(如多酚氧化酶、抗坏血酸氧化酶、细胞色素氧化酶)的组成成分,可以影响氧化还原过程,还存在于叶绿体的质体蓝素中,参与光合作用的电子传递体系[13],对植物的生长发育起着十分重要的作用。另外,养殖塘中种植沉水植物能够改善水质、抑藻控藻、提高池塘生态多样性以及增强养殖水环境稳定性,所以用沉水植物来修复养殖塘底泥重金属Cu污染能够带来较高的生态经济效益[12,14]。
苦草(Vallisnerianatans)、轮叶黑藻(Hydrillaverticillata)和伊乐藻(Elodeanuttallii)是我国淡水养殖塘常见的多年生沉水草本植物,生物量大且生命力强,但是过量的Cu同样会对这些植物生长造成不良影响,会使植物生长发育受阻、生理代谢过程发生絮乱等危害[15-17]。因此,本研究选择这三种沉水植物作为试验材料,通过综合分析其对底泥重金属Cu的富集效果及相关机理,筛选最佳修复淡水养殖塘底泥重金属Cu污染的沉水植物,利用底泥-植物系统的吸附、转化及自我调控功能,定期收获地上部分并达到一定程度上对底泥Cu污染的修复,为今后深入修复重金属Cu污染底泥提供一定的研究依据。
1 材料与方法
1.1 供试底泥
供试底泥采自湖州市菱湖镇加州鲈鱼养殖塘,自然风干后,先初步剔除其中粗杂质,过100目筛,测得Cu的背景值为28.34 mg/kg,pH为7.2左右。用分析纯的Cu(NO3)2溶液,拌入一部分风干的原始底泥中并充分混匀,静置钝化两周风干后作为试验底泥,测得配制后底泥中Cu含量为112.42 mg/kg,Cu污染程度为土壤环境质量Ⅲ级标准,达到JENSEN底泥污染评价重污染级别。
1.2 供试植物
供试沉水植物苦草、轮叶黑藻和伊乐藻均采自浙江省湖州市养殖池塘。选取高度为15 cm左右、生长状态良好、个体均匀带根的植株作为试验材料,移栽到无污染底泥的生态缸中,在1/10霍格兰营养液水培适应3 d后用于盆栽Cu污染修复试验。
1.3 实验设计
本试验共设3个处理组,3个空白对照组,每个处理组和对照组各设3个重复。其中,HV:轮叶黑藻空白对照组,HV+Cu:轮叶黑藻Cu处理组,EN:伊乐藻空白对照组,EN+Cu:伊乐藻Cu处理组,VN:苦草空白对照组,VN+Cu:苦草Cu处理组。试验采用透明的玻璃生态缸(长30 cm×宽20 cm×高50 cm)静态模拟自然状况下小型水体,将其置于通风良好的室内,其上30 cm处悬挂仿太阳灯,其光照度6 000 lx,光照时间为每天8∶00~20∶00。将原始底泥和配制好的底泥分别铺入18个生态缸内,厚度约15 cm。然后每个生态缸中加入25 L曝晒的自来水,生态缸内水深约40 cm,然后将初始鲜重200 g的三种沉水植物分别植入盛有空白对照组和Cu处理组底泥的生态缸中,经测定实验初期水体中总氮平均浓度为1.2 mg/L,硝态氮为 0.71 mg/L,全磷为0.16 mg/L,其中Cu处理组水体中铜离子浓度为0.82 mg/L。培养试验从2017年8月2号开始至11月29号结束,整个试验周期为120 d,三种沉水植物基本完成一个生长周期,同时为了保证植物正常生长营养元素的需要,每14 d加入100 mL霍格兰营养液(铜离子浓度为0.025 mg/L)。每30 d收割一次植物地上样品(分为茎部和叶部)及植物根际土样,最后一次收集整株植物并分别取根茎叶三个部位。试验中蒸发的水分定期用曝晒的自来水补充。
1.4 样品处理
Cu含量的测定:取植物样先用自来水充分冲洗,再用0.1 mol/L的HCl洗净,最后用去离子水淋洗2~3次,吸干表面水分,将植物样品分为根部、茎部和叶部三部分,置于烘箱中105 ℃杀青30 min,70 ℃下烘干至恒重,称取各部位干重,研磨过60目尼龙筛待测;将土样自然风干后,研磨过100目尼龙筛待测。用HNO3-HClO4消解植物样,HCl-HNO3-HClO4消解土样。用电感耦合等离子体原子发射光谱法进行Cu含量的测定。
SOD和POD酶活的测定:取植株相同部位的鲜叶片,用蒸馏水洗净、揩干,用于测定各项生理指标。SOD、POD酶液制备时取材于预冷研钵中,加入pH 7.8磷酸缓冲液(0.05 mol/L),冰浴下研磨成匀浆,10 000 r/min低温(4 ℃)离心20 min,上清液即为所需要的酶液。SOD和POD酶活测定均采用南京建成生物工程研究所试剂盒及美国伯腾仪器有限公司的Microplate Reader Gen5测定。
土壤脱氢酶活性采用氯化三苯基四氮唑(TTC)法测定[18]。
1.5 数据处理与分析
(1)生长率:
GR=dw/dt
式中:GR为生长率;w为植物增加的生物量;t为植物生长的时间。
(2)去除率:
R=(C0-C)/C0×100%
式中:R为去除率;C0为试验开始时底泥中重金属含量;C为试验结束时底泥中重金属含量。
用Excel 2013、SPSS 18.0统计分析软件进行数据分析处理。图由Excel 2013绘制,表由Excel 2013生成,使用SPSS 18.0软件中的独立样本t检验对数据进行显著性差异比较,描述性统计使用平均值±标准差(Mean±SD)表示,P<0.05代表差异显著。
2 结果与分析
2.1 重金属铜对三种沉水植物生长特征的影响
表1显示,与移植时相比,试验结束时三种沉水植物收获的生物量均有明显增加,这表明浓度为112.42 mg/kg的Cu胁迫对三种沉水植物的生长并没有造成严重的影响。轮叶黑藻和伊乐藻在整个生长期间,Cu处理组收获总鲜重显著低于对照组,而苦草总鲜重虽然低于对照组,但差异不显著,说明苦草对Cu胁迫表现出较强的耐受性。
轮叶黑藻在前3次收割时,Cu处理组的茎部和叶部鲜重均显著低于不加Cu对照组,而在第四次收割时处理组之间均差异不显著;伊乐藻在前两次收割时,Cu处理组茎部鲜重低于对照组,但差异不显著,而叶部鲜重显著低于对照组,在第三、四次收割时,除第四次收割的根部鲜重显著低于对照组外,其他均差异不显著;苦草在第一次和第四次收割时,处理组间叶部鲜重存在显著差异,且最后一次收获的处理组根部鲜重显著低于对照组,其他均是Cu处理组根部和叶部低于对照组,但差异不显著。说明Cu胁迫条件下,沉水植物尽管能正常生长,但其根茎叶生长受到不同程度的影响,具体影响程度因品种、收获时间不同而存在一定差异。
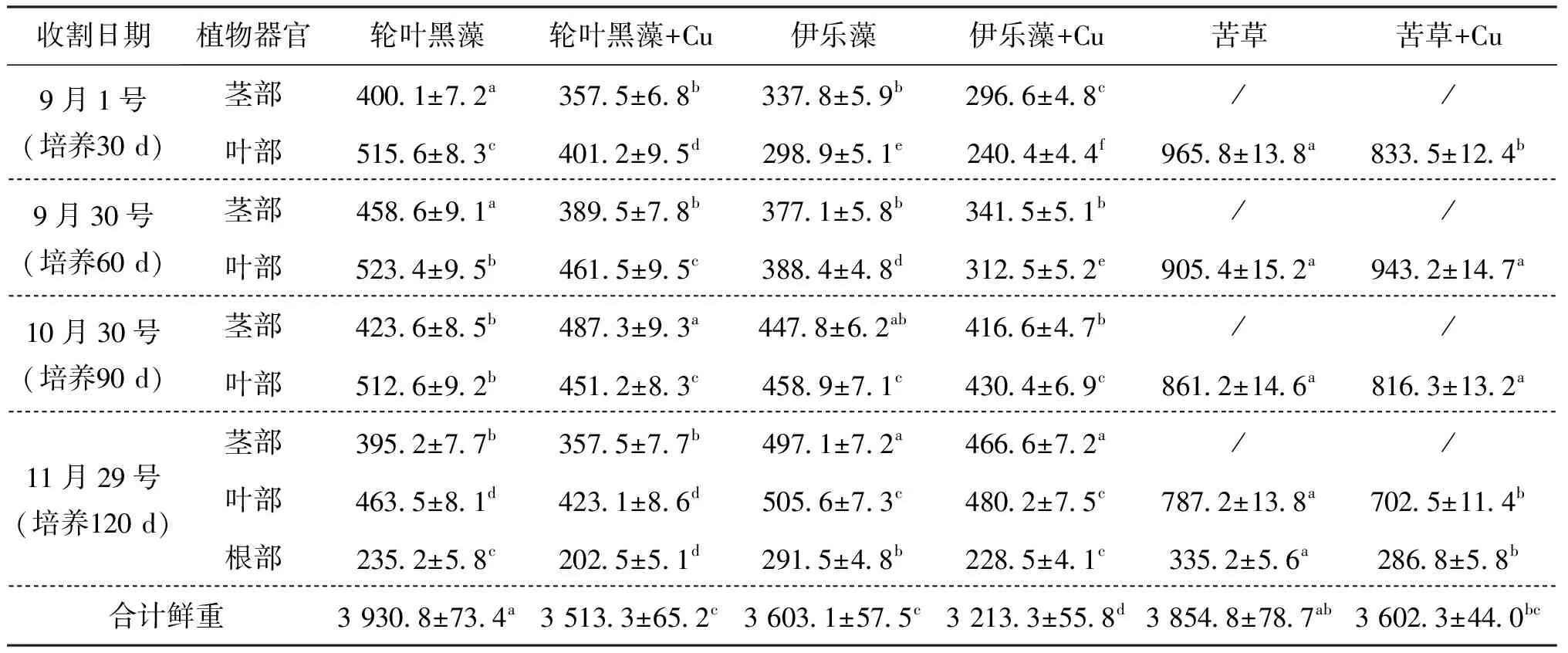
表1 三种沉水植物根茎叶收获鲜重变化特征
注:表中数据为平均值±标准差;同行不同小写字母表示不同处理之间在0.05水平存在显著差异(n=3),下同。
2.2 三种沉水植物不同部位对重金属铜的累积差异
从表2可以看出Cu处理组和对照组中每种沉水植物收获时根部含Cu量均显著大于叶部,而叶部含Cu量显著大于茎部。值得注意的是整个实验期间,大部分茎和叶被定期收割三次,根部则未被收割,而最终根部Cu含量只是茎叶含量的1.20~1.64倍,说明根部对Cu吸收富集作用存在一定阈值。通过比较各对照组之间三种沉水植物根茎叶Cu含量发现,其大致顺序依次为轮叶黑藻>苦草>伊乐藻,而Cu处理组中其大致顺序依次为苦草>轮叶黑藻>伊乐藻。结果表明苦草较其他两种沉水植物对Cu具有更强的耐受能力,可以作为受Cu污染养殖塘底泥植物修复的先锋植物,而轮叶黑藻可以作为未受Cu污染养殖塘底泥进一步生物净化的植物品种。
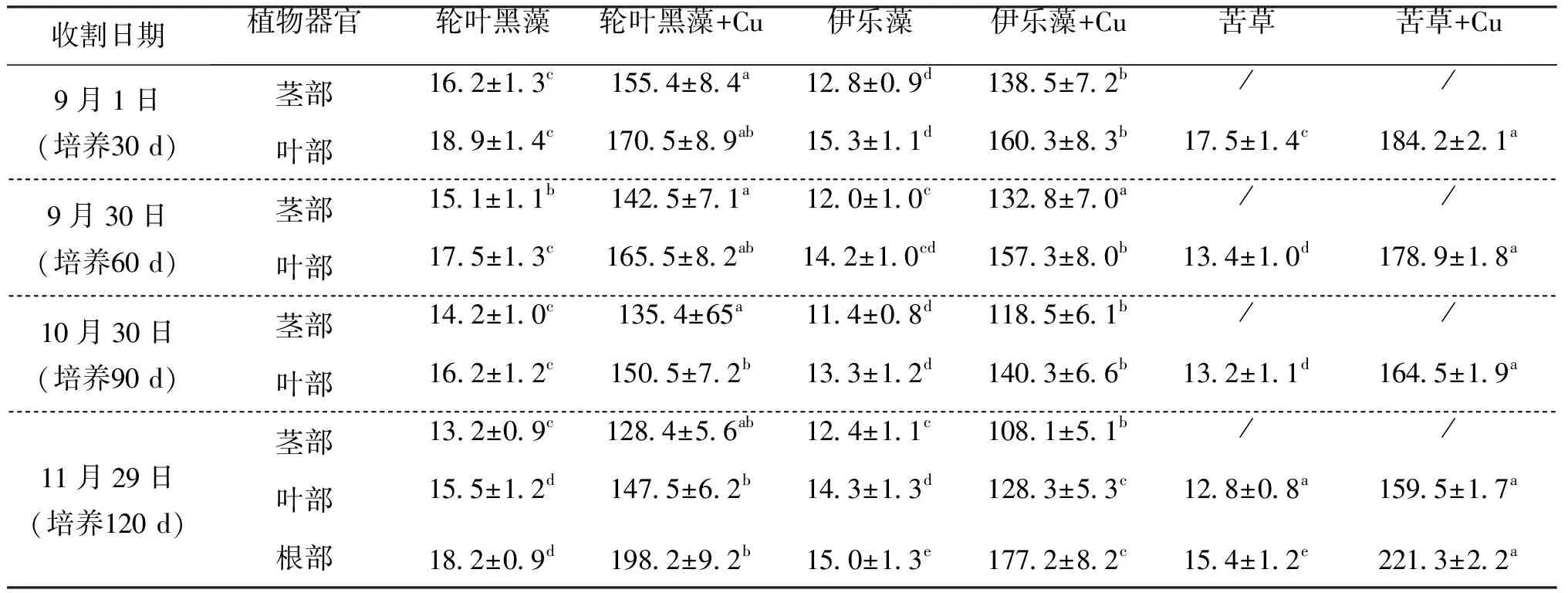
表2 三种沉水植物收货时根茎叶Cu含量变化特征
植物对重金属的吸收分布情况是耐性物种选择的重要指标之一,根据沉水植物不同部位含铜量和生物量计算了三种沉水植物的Cu累积量。从图2可以看出,各对照组之间,Cu累积量顺序依次为轮叶黑藻>苦草>伊乐藻,但无显著性差异;而对于Cu处理组,Cu累积量顺序依次为苦草>轮叶黑藻>伊乐藻,且差异显著,主要是由于处理组中苦草具有较高的生物量和各部位较高的Cu含量。另外轮叶黑藻各对照组和Cu胁迫处理组各部位Cu累积量大小顺序为叶部>茎部>根部,且各部位差异显著;而伊乐藻叶部和茎部Cu累积量差异不显著,但是均显著大于根部累积量;苦草的Cu累积量则主要集中在叶部,根部只占极少一部分。说明不同植物不同部位Cu累积量存在一定差异,对于养殖塘底泥重金属Cu污染修复来讲,叶部累积量较大的沉水植物较适宜种植,因可通过定期收割地上部分来不断降低底泥重金属含量直至达到安全水平。
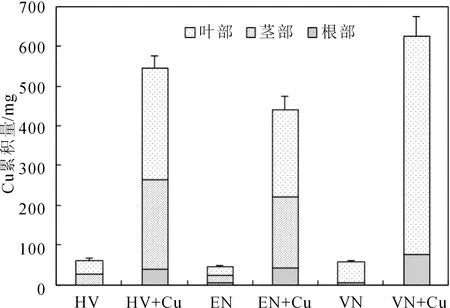
图2 三种沉水植物不同部位Cu累积量特征
2.3 底泥中重金属铜含量随时间的动态变化
随着沉水植物的生长和收割,Cu处理组底泥中Cu含量显著下降,至实验结束时,Cu处理组中由轮叶黑藻、伊乐藻和苦草修复的养殖塘底泥分别由初始(112.42±6.6) mg/kg下降至(49.15±4.4)、(59.65±4.2)和(45.62±2.8)mg/kg,三种沉水植物Cu去除率依次为(48.62±2.5)%、(39.27±2.1)%和(55.72±2.8)%,各处理组之间差异较显著;而对照组底泥中含Cu含量则稍有下降,但下降趋势较平缓且不显著,其对应去除率分别为(19.90±1.0)%、(21.88±1.2)%和(16.23±0.8)%(表3)。处理组中,经轮叶黑藻和苦草富集后的底泥间Cu含量显著低于伊乐藻,而轮叶黑藻和苦草组底泥Cu含量则差异不显著。试验结束时底泥中Cu含量与试验开始的污染底泥相比皆呈显著差异,说明底泥Cu污染有很大改善。
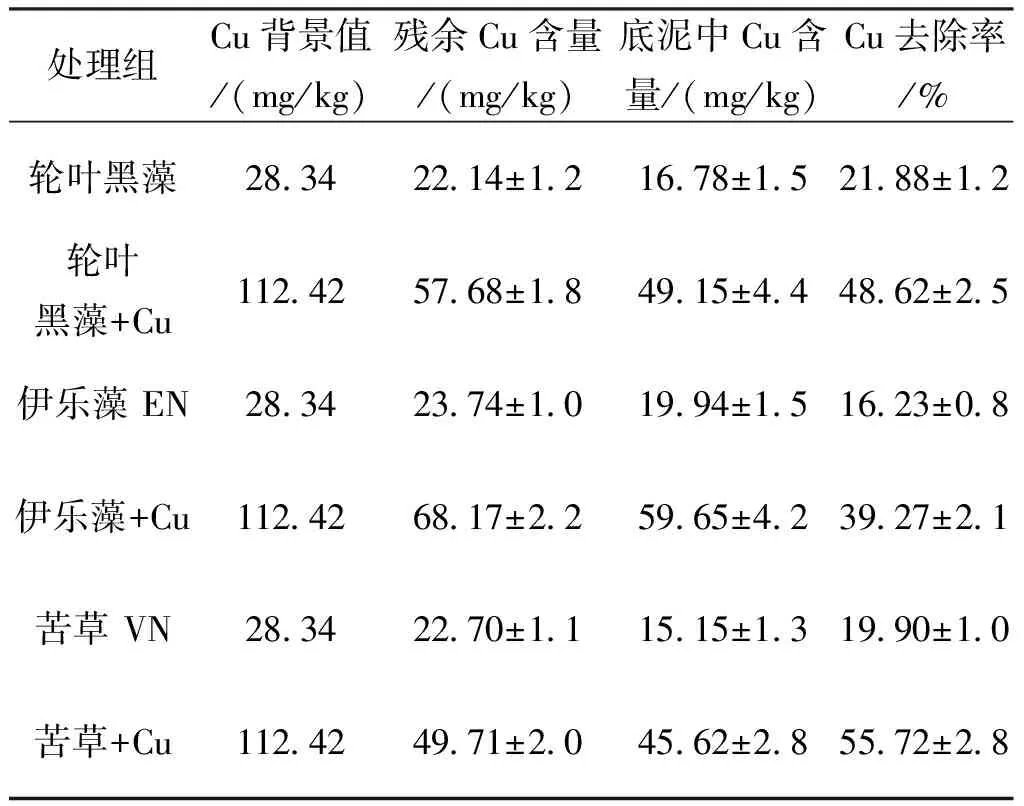
表3 三种沉水植物对底泥重金属Cu的去除率
2.4 三种沉水植物抗氧化酶活性变化特征
图3显示了三种沉水植物在Cu胁迫下抗氧化酶活性随时间变化特征。从图中可以看出,在第一次收割时Cu处理组SOD活性与初始值相比显著上升,随后不同程度下降;而与对照组相比则显著提高,在其后三次收割其提高并不明显。Cu处理组POD活性在第一次收割时其相对于初始值显著提高,此后其活性基本保持不变,且苦草稍大于轮叶黑藻,而两者则显著大于伊乐藻,另外Cu处理组的POD活性显著高于各自对照组,而对照组在整个试验周期内POD活性基本保持不变。
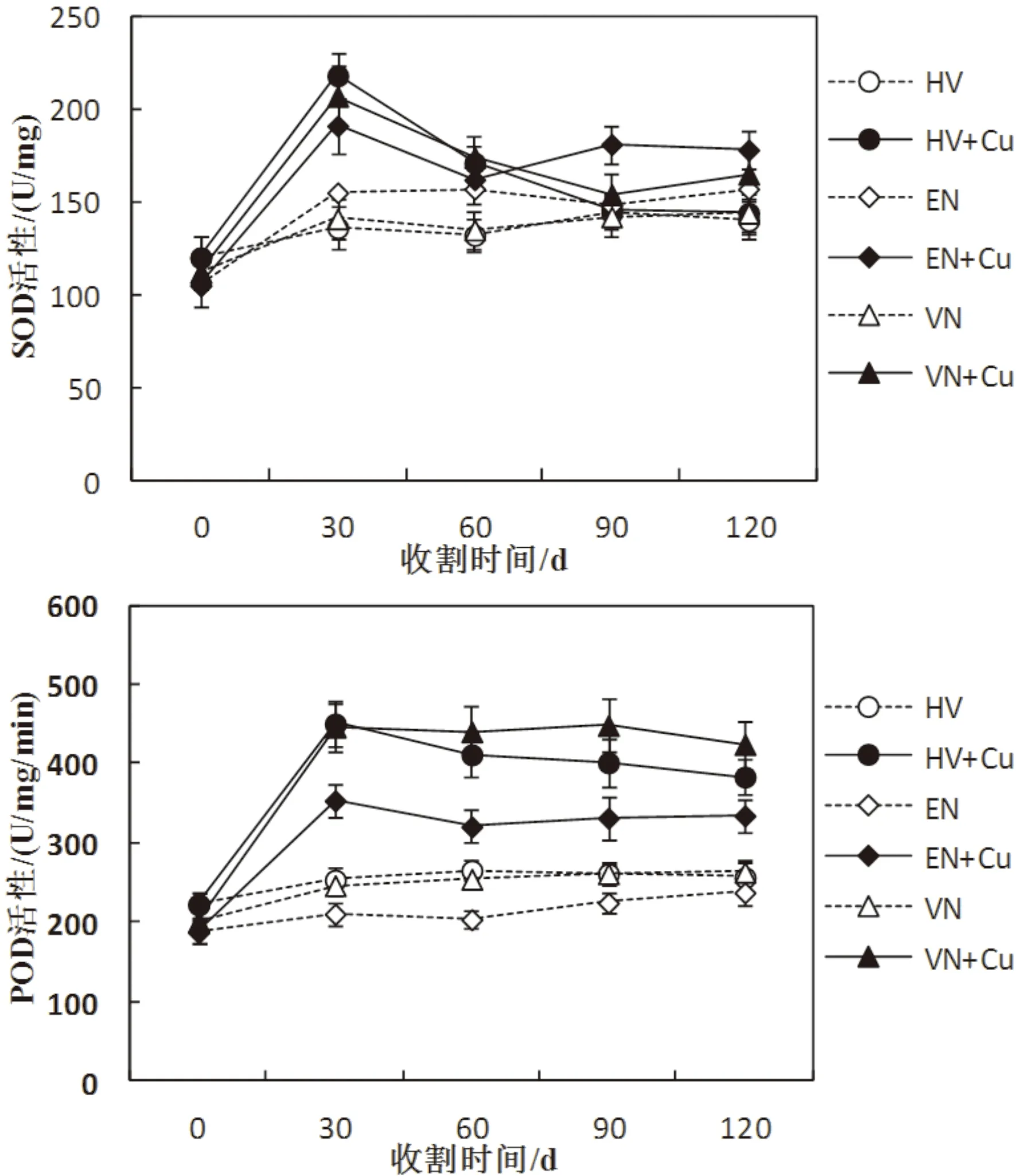
图3 三种沉水植物SOD和POD活性变化特征
2.5 三种沉水植物根际底泥脱氢酶活性变化情况
图4显示了三种沉水植物根际底泥脱氢酶活性的变化特征,可以看出,轮叶黑藻和苦草各对照组土壤脱氢酶活性在前两次收获(9月1日和30日)均快速增加,而自第三次收割(10月30日)后则开始缓慢下降,伊乐藻则整个生长周期逐渐增加至平稳状态,3个处理组脱氢酶活性均显著低于对照组。这主要是由于刚刚种上沉水植物,作物的根际不是很发达,土壤中微生物的数量也不是很多,所以脱氢酶的活性较低。随着植物的生长及根际环境的改变,脱氢酶活性的上升速度较快,之后随着有机物的降解、植物产量及微生物量的增加,脱氢酶活性缓慢增加,轮叶黑藻、苦草和伊乐藻在第三次收割时均达到最高水平。随着轮叶黑藻和苦草的生长、天气转冷脱氢酶活性逐渐降低,表明轮叶黑藻和苦草对沉积物中重金属Cu污染物的修复能力也逐渐降低。而伊乐藻具有耐低温生理特征,在低温条件下生长良好,尽管温度降低,但是根际活性较好,故脱氢酶活性仍缓慢增加至趋于平稳,因此可以选择伊乐藻在寒冷的冬季进行底泥重金属污染修复品种,为冬春季节或低温条件下水体重金属污染植物修复提供依据。而Cu胁迫下,各处理组脱氢酶活性均显著低于各对照组,且脱氢酶活性增加较缓慢,至后期逐渐趋于平稳。说明Cu会明显抑制底泥脱氢酶活性,且随着底泥Cu含量的逐渐降低,其抑制作用逐渐解除,活性越来越高。
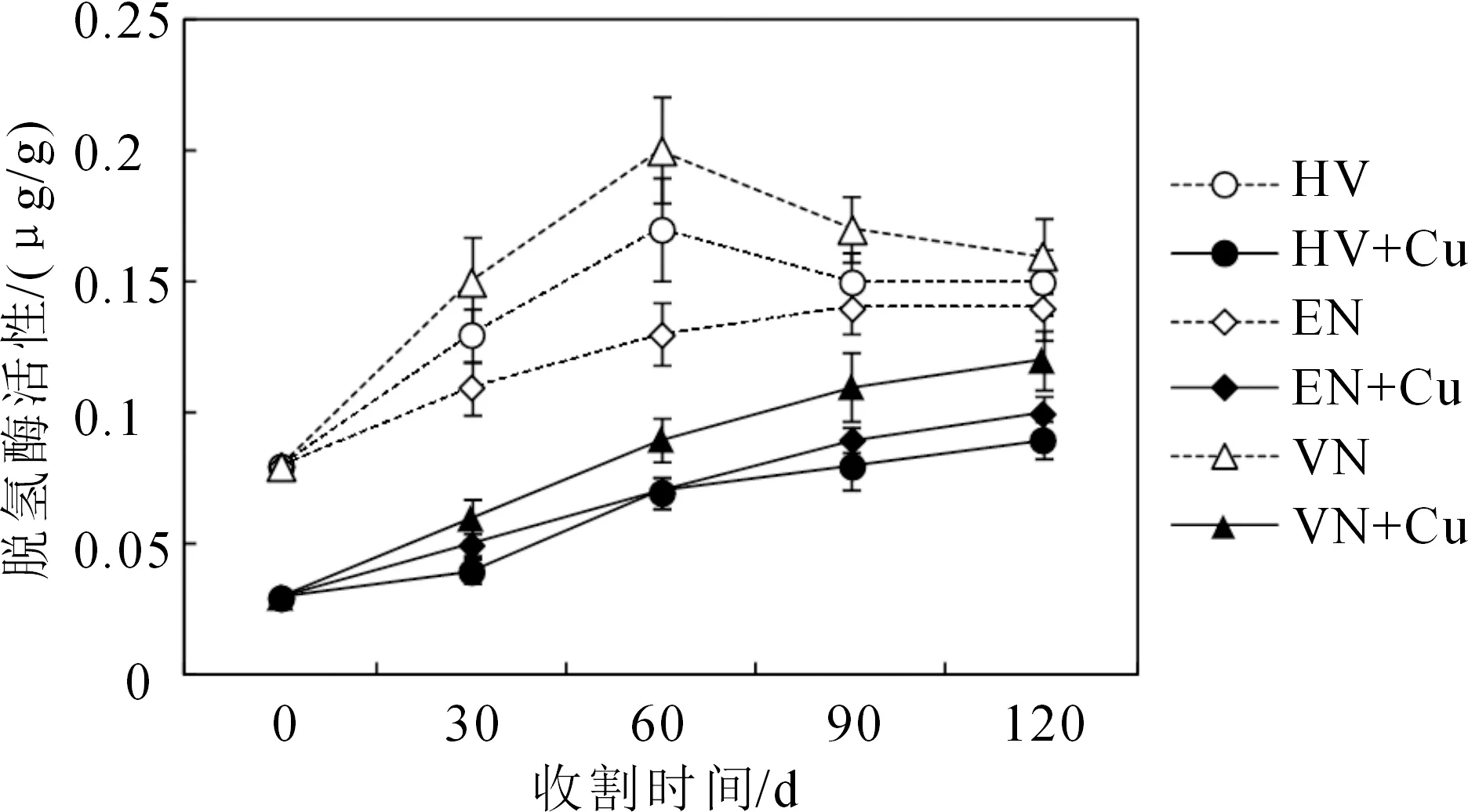
图4 三种沉水植物根际底泥脱氢酶活性变化特征
3 讨论
生物量的大小是衡量植物修复潜力大小的一个重要指标,测量植物生物量在重金属胁迫下的变化也能反映出植物对重金属的耐性大小[12,19]。试验期间,苦草、轮叶黑藻和伊乐藻并未发生失绿、萎蔫和死亡等现象,说明三种沉水植物对底泥相对较高的Cu浓度均具有一定的耐受性,并未超出其耐受临界值。收获生物量大小顺序依次为苦草>轮叶黑藻>伊乐藻,分析原因,主要是沉水植物生长除了与Cu抗胁迫性有关外,还与其自身生长环境密切相关,苦草和轮叶黑藻较适宜于在水温为20~30 ℃内生长,而伊乐藻较适宜低温(10~15 ℃)环境生长,而本试验期间,前三次收割时平均温度均在20 ℃以上,仅最后一个月平均温度约为15 ℃,同时由于轮叶黑藻和伊乐藻根部的退化,而苦草具有相对发达的根系,水生植物的根系越发达越利于营养物质的吸收。
另外,植株不同部位对重金属都有一定积累量,并且积累程度存在一定差异[20]。本研究中,一定Cu浓度下,根茎叶部富集Cu的能力是受限的,并不会随着时间的延长而提高,因此通过定期收割地上多余生物体以促进植物快速生长,从而达到快速富集底泥中重金属Cu的目的。本研究发现,三种沉水植物富集能力顺序依次为苦草>轮叶黑藻>伊乐藻,均可作为底泥Cu污染修复植物,且对重金属的吸收存在浓度效应,即随着环境中重金属离子浓度的升高,植物富集的重金属含量也会相应增加,这与许多研究结果相一致[10-12],如超累积植物蓖麻体内Cu含量随着Cu矿废弃地土壤含量的增高而增高[21]。因此,不同植物对重金属的耐性、吸收、转运能力相差较大,在应用沉水植物修复池塘底泥重金属污染时应根据植物特性和污染特征进行针对性修复。通过本试验结果可知,尤其是已知底泥有轻度Cu污染的淡水养殖塘,在不影响其正常生产,又能保证其生产效益的情况下,宜于春季种植苦草和轮叶黑藻,秋季宜种植伊乐藻,进而通过定期收割以移除底泥中多余的Cu,达到底泥中Cu生物净化或修复的效果。
植物超量累积重金属是基于多种防御机制[22]。许多研究认为,重金属对植物的伤害与自由基的产生有关,生物体主要通过抗氧化酶系统防御自由基损伤[23]。SOD、POD等是植物体内抗氧化酶系统中几种主要的保护酶,SOD作为超氧自由基清除剂,其活性高低与植物抗逆性大小有一定的相关性,在适度的逆境环境诱导下,SOD活性增加能提高植物的适应能力[24-25]。POD是植物体内分布较广的一类氧化还原酶,具有重要的生理防护功能,如木质素的形成伸展、蛋白的聚合、植物生长素的代谢、病毒的抵抗和创伤的愈合等[26-27]。在本实验中,SOD和POD的活性出现一定程度的升高,这是植物体的保护性反应,能有效地清除毒害所产生的活性氧,说明沉水植物对重金属Cu胁迫具有一定的抗逆性能和适应能力,根据SOD和POD活性可知适应能力依次为苦草>轮叶黑藻>伊乐藻,这与上述三种沉水植物鲜重以及累积量结果相一致。同时,植物的生长会不断激活土壤酶活性,土壤酶活性的升高又可促进植物的生长,从而促进植物对重金属的转化吸收,增强其富集能力[28-29]。本试验亦发现,随着三种沉水植物的不断生长,其根际土壤脱氢酶活性逐渐恢复,但重金属Cu对土壤脱氢酶的活性亦产生了不同程度抑制。对照组脱氢酶活性受温度的影响较大,随着温度的逐渐降低,苦草和轮叶黑藻生长受限,脱氢酶活性受到一定抑制,呈现不断降低趋势,而伊乐藻生物量继续增加,说明植物生长旺盛阶段对脱氢酶的激活作用要大于温度的抑制作用,因此其活性继续缓慢的上升至稳定状态。而Cu处理组中,尽管温度不断下降抑制了脱氢酶活性,但随着Cu含量的不断降低,对脱氢酶活性的抑制作用大大减弱,从而抵消了温度降低的抑制作用,提高了土壤脱氢酶的活性。因此,在利用沉水植物修复养殖塘底泥重金属时应尽量减弱外界环境因素对土壤酶的抑制作用,比如增加沉水植物的生长时间或增加其收割次数或选择适宜其生长的温度条件等以不断增强土壤酶的活性,促进植物生长进而促进吸收转移底泥中过量的重金属,又可不断减轻重金属对植物的胁迫作用。
猜你喜欢
农业灾害研究(2023年8期)2023-11-07 03:21:48
科学养鱼(2022年1期)2022-02-17 02:12:14
科学养鱼(2021年5期)2021-06-17 04:39:30
农业知识(2020年10期)2020-12-20 17:20:43
至爱(2020年10期)2020-11-20 02:41:06
当代水产(2019年10期)2019-12-18 05:28:56
农机化研究(2019年9期)2019-05-24 09:53:42
中南民族大学学报(自然科学版)(2018年4期)2018-12-29 01:04:30
农村百事通(2017年21期)2017-11-30 16:31:01
中国酿造(2017年4期)2017-04-27 07:06:28
