
天女木兰种子天冬氨酸蛋白酶基因的克隆与分析
2019-05-13 12:58:16张晓林陆秀君
中南林业科技大学学报 2019年5期
侯 哲,梅 梅,张晓林,陆秀君
(1.沈阳农业大学 林学院,辽宁 沈阳 110866;2.中国林业科学研究院 a.林业研究所;b.国家林业局森林培育重点实验室;c.林木遗传育种国家重点实验室,北京 100091)
天女木兰Magnolia sieboldiiK.Koch 是世界上稀有的珍贵树种之一,主要分布于中国北部地区,朝鲜、日本也有少量的分布。天女木兰具有广泛的开发利用价值,花色洁白素雅,异常芬芳,是一种庭院绿化及观赏树种,木质优良,树干通直,可做家具和雕刻用材。
天女木兰种子不同部位都存在不同程度的发芽抑制物质,种胚发育不完全造成了种子的深休眠[1]。种子在可收获的时候,胚乳中存在的不同浓度的植物激素,是致使种子难以萌发的又一个重要原因[2]。合理的浓度和浸种时间有利于提高种子质量,促进种子萌发[3]。生理休眠是天女木兰种子难以萌发的原因,种皮的机械束缚并不能造成种子深度休眠,其主要原因是种胚发育不完全,对种子进行150 d 的低温沙藏后高温15 d 催芽,种子可以达到90%的发芽率[4]。将GA3质量浓度设置为300 mg/L 后浸种,通过变温层积催芽效果较好,且在整个沙藏过程中温度起主导作用。而种胚的生长分化在采用GA3浸种变温层积处理后会有一定的促进作用,这是因为种子内各种促进种胚成熟物质的积累对种胚的生长产生有利的影响[5]。
种子打破休眠进而萌发是一个复杂的生理过程,可以通过蛋白质组学的研究来洞悉其分子机理[6]。木本植物在发芽期间的蛋白组变化的研究已有报道,包括山楂[6]、山毛榉[7]、槭树[8]、麻风树[9]、凤仙花[10]和南洋杉[10]等。本课题组在前期的研究中,对天女木兰种子进行变温层积处理,分别比较了种子在层积0、6、110 和150 d 的个体发育变化和蛋白组差异变化,以期确定与种子萌发相关的蛋白质,从而进一步了解种子萌发的分子机理[11]。
天女木兰层积催芽的种子在休眠解除过程中,差异表达蛋白天冬氨酸蛋白酶在层积过程中随种胚的伸长生长呈波动变化[12],表明天冬氨酸蛋白酶在种子萌发过程中发挥着重要作用。天冬氨酸蛋白酶是重要的蛋白水解酶,参与机体的新陈代谢及生物调控作用,广泛存在于植物种子、花及叶片等组织中。有研究表明,天冬氨酸蛋白酶在种子休眠解除过程中起主导作用,参与了种子萌发期间储藏蛋白的水解[12]。拟南芥PCS1基因编码的天冬氨酸蛋白酶参与调控胚胎发育,而PCS1突变体缺失该功能,导致发育胚细胞死亡和雌雄配子体退化[13]。
众多研究表明,天冬氨酸蛋白酶在种子休眠解除过程中起主导作用。本课题组在天女木兰种子休眠解除过程中种胚发育和相关蛋白的研究中指出,天冬氨酸蛋白酶在层积过程呈波动变化,在110 d 时达到高峰,说明该蛋白参与种子萌发期间储存蛋白的水解,在种子成熟胚的发育过程中发挥重要作用[11]。本研究以天女木兰种子为试验材料,在建立天女木兰总RNA 提取体系的基础上,利用同源克隆法RACE 技术从天女木兰种子中克隆得到天冬氨酸蛋白酶基因全长,采用生物信息学的方法进行分析,利用分子生物学技术对天女木兰种子休眠及其萌发机理进行深入研究,这对于天女木兰这一“活化石”树种的保护和开发利用具有非常重要的意义。
1 材料与方法
1.1 植物材料
试验用到的天女木兰种子采于沈阳农业大学植物园(40°49′N,123°34′E)内8年生的健康植株,于9月末采集种子等材料并迅速冷冻于液氮中,或者冷藏于-80℃的超低温冰箱中,直到RNA 提取。
1.2 RNA 提取及cDNA 合成
参照北京天根生化科技有限公司RNA 提取试剂盒(DP419)说明书进行RNA 的提取,按照PrimeScriptRT(TaKaRa,Japan)试剂盒说明书反转录合成cDNA。
1.3 天冬氨酸蛋白酶全长基因的克隆
1.3.1 基因中间片段的获得
1.3.1.1 设计天冬氨酸蛋白酶的简并引物
根据已公布的麻疯树、梅花、油棕、绿豆等天冬氨酸蛋白酶的基因序列,利用Primer5.0 设计简并引物(表1),交由沈阳汇佰生物科技有限公司合成。

表1 简并引物Table1 Degenerate primers
1.3.1.2 PCR 扩增目的片段
PCR 用的是promega 的Go Taq DNA 聚合酶,反应体系是20 μL 体系,温度梯度从48 ℃到65 ℃,扩增体系严格按照试剂说明书进行。
1.3.1.3 目的片段的回收
为了进一步纯化目的片段cDNA,使用全式金的EasyPure® Quick Gel Extractiom Kit 胶回收试剂盒进行目的片段的回收。
1.3.1.4 目的基因与pGEM®-TEasy 载体连接及转化
目的基因与pGEM®-TEasy 载体连接转化反应步骤参照pGEM®-TEasy 载体说明书。
1.3.1.5 阳性克隆载体的鉴定
筛选出只有白色菌落的部位,取出一部分稀释到10 μL ddH2O 中,取2 μL 菌液作为PCR 模板,进行阳性克隆载体的鉴定。
1.3.1.6 测序及序列分析将符合要求的菌液送至沈阳佰创生物技术有限公司进行测序。测完序后用BLAST 软件进行序列比对分析。
1.3.2 天冬氨酸蛋白酶基因的3′ RACE 扩增
根据已得到的天冬氨酸蛋白酶基因中间保守序列设计3′RACE特异性引物RC226-F1、RC226-F2,反转录引物、接头引物及特异性引物序列如表2所示。
以3′ RACE Adaptor 为反转引物的cDNA 为模板进行3′ RACE 巢式PCR 扩增(参照invitrogen 3′ race system manua 说明书进行),直到获得3′目的片段。将巢式PCR 产物回收纯化、连接载体、转化测序。
1.3.3 天冬氨酸蛋白酶基因的5′ RACE 扩增
5′ RACE 特异性引物、反转录引物、接头引物及特异性引物序列如表3所示。
以特异性引物RC226-RT1 为反转录引物,得到cDNA,经RNase H 和TdT 处理后,进行巢氏PCR(操作步骤见invitrogen 5′ race system manual),直到获得5′目的片段,然后将巢式PCR 产物回收纯化、连接载体、转化测序。

表2 3′RACE引物Table2 3′ RACE adaptor

表3 5′RACE引物Table3 5′ RACE adaptor
1.3.4 DNAMAN 拼接全长序列
将得到的天冬氨酸蛋白酶基因的保守序列、5′RACE序列及3′RACE序列通过DNAMAN软件进行拼接,进一步剪切掉重叠序列就可以得到天冬氨酸蛋白酶基因的全长cDNA 序列。
1.4 天冬氨酸蛋白酶基因的生物信息学分析
将得到的全长序列通过BLAST 中的nucleotide blast 进行基因的同源性分析,将全长cDNA 通过DNAMAN 软件转化为氨基酸序列后,输入Blastp软件进一步进行氨基酸的同源性分析,Clustalx 1.83 和MEGA 4.1 结合构建系统进化树进行亲缘性分析。目的基因翻译成蛋白后预测分析其理化性质,如相对分子量与等电点可以通过ProtParam(http://www.expasy.org/tools/protparam.html)进行分析预测;亲水性与疏水性可以用ProtScale 软件分析;蛋白的结构域、二级结构与三维结构可以通过软件Phyre2 预测分析;天冬氨酸蛋白酶基因的跨膜结构是否存在可以通过在线分析软件TMHMM Server-2.0 预测;利用Plant-mPLoc 软件对天女木兰天冬氨酸蛋白酶基因编码的蛋白进行亚细胞定位预测分析。
3 结果与分析
3.1 天女木兰种子总RNA 的提取
所得到的RNA 十分完整,28 S RNA 的亮度是18 S RNA 的2 倍,完全满足后续的分子试验要求(图1,表4)。
3.2 天冬氨酸蛋白酶基因全长的克隆
3.2.1 天冬氨酸蛋白酶基因中间序列的克隆
由简并引物克隆得到的序列回收纯化、连接载体后转入感受态细胞中,选取阳性克隆载体,从电泳检测结果(图2)可以看出1 和2 目的片段连接到载体上,可以进行测序分析。
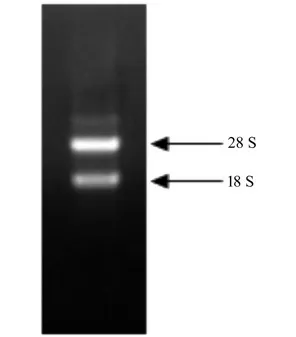
图1 天女木兰种子总RNA 电泳检测 Fig.1 Electrophoresis detection of total RNA of M.sieboldii seeds

表4 天女木兰总RNA质量浓度和产率的检测Table4 Detection of concentrations and productivities of M.sieboldii total RNA

图2 阳性克隆载体的鉴定Fig.2 Identification of positive cloning vector
3.2.2 天冬氨酸蛋白酶基因的3′ RACE 扩增
将3′ RACE 的PCR 产物回收纯化后测序得到了224 bp 的碱基序列(图3),该序列具有天然的poly(A)尾巴,且与已获得的中间序列可以通过DNAMAN 软件准确拼接,BLAST 后发现与其他植物中的天冬氨酸蛋白酶基因同源性较高,可以确定克隆到的该序列为天女木兰天冬氨酸蛋白酶基因的3′端。
3.2.3 天冬氨酸蛋白酶基因的5′ RACE 扩增

图3 3′ RACE 结果Fig.3 Results of 3′ RACE
将5′ RACE 的PCR 产物回收纯化后测序得到了265 bp 的碱基序列(图4),该序列与已获得的中间序列可以通过DNAMAN 软件准确拼接,BLAST 后发现其与其他植物中的天冬氨酸蛋白酶基因同源性较高,可以确定克隆到的该序列为天女木兰天冬氨酸蛋白酶基因的5′端。

图4 5′ RACE 结果Fig.4 Results of 5′ RACE
3.2.4 天冬氨酸蛋白酶基因ORF 的获得
将天冬氨酸蛋白酶基因的中间、3′端和5′端3 个片段经过DNAMAN 软件拼接后得到了1 798 bp 的碱基序列,全长序列与已知的天冬氨酸蛋白酶基因的同源性较高,进一步验证了该序列为天女木兰天冬氨酸蛋白酶基因(图5、图6)。

图5 全长cDNA 示意图Fig.5 Schematic diagram of the full length cDNA

图6 天冬氨酸蛋白酶的核苷酸序列及氨基酸序列Fig.6 Nucleotide sequences and amino acid residues sequences of aspartic acidase
3.3 天冬氨酸蛋白酶基因的生物信息学分析
3.3.1 天冬氨酸蛋白酶基因的相似性分析
通过NCBI 中的Blastx 数据库对获得的天女木兰天冬氨酸蛋白酶基因全长进行氨基酸同源性分析,比对结果显示目的序列推导的氨基酸序列与其它植物中的已知的天冬氨酸蛋白酶基因的氨基酸序列的同源性都在85%以上,其中与莲科的莲花Nelumbo nucifera(XP 010252464.1)以及大戟科的蓖麻Ricinus communis,(XP 002529926.1)等的相似性最高,达到了87%;与其他植物的同源性都在84%~87%之间,同源性非常高,进一步验证了所获得的全长序列是天女木兰的天冬氨酸蛋白酶基因全长,比对结果见表5。
通过MEGA 5.1 软件中Neighbouring-Joining聚类法对天女木兰的天冬氨酸蛋白酶的氨基酸序列与其它植物的天冬氨酸蛋白酶氨基酸序列构建进化树(图7)。从图7中可以看出,26 种植物的天冬氨酸蛋白合成酶基因由于亲缘关系被分成三大类,就聚类分析来看,与睡莲科的莲花首先聚为一类,这与BLAST 同源性分析结果一致,可能是因为二者均为开花类的观赏植物,属于直系同源蛋白,与豆科的芸豆与豇豆亲缘关系较远。本研究在木兰科及其亲缘关系较近的其他植物的分子水平研究方面奠定了一定的基础。
3.3.2 Ms Aspartic proteinase 基因编码的蛋白结构的分析
3.3.2.1 基本理化性质的分析
将Ms Aspartic proteinase 编码的氨基酸序列输入在线软件ProtParam,分析预测其基本的理化性质,通过软件分析可知,Ms Aspartic proteinase 编码的蛋白质的分子量为55.176 KDa,理论等电点为5.25,推断为含有酸性的蛋白;带有49 个负电荷残基(Asp+Glu),40 个正电荷残基(Arg+Lys);共含有7 714 个原子,其中C、H、N、O、S 的原子含量分别为2 452、3 839、651、748、24;不稳定系数计算后为37.31,归于稳定蛋白一类。
目的基因的亲水性与疏水性可以通过在线分析软件ProtScale 进行预测分析(图8),该蛋白的总平均亲水性数值为0.015,因此判定天女木兰天冬氨酸蛋白酶属于疏水性蛋白。

表5 天冬氨酸蛋白酶基因和其它25种植物天冬氨酸蛋白酶基因推导的氨基酸序列相似性比对Table5 Similarity comparison of aspartic proteinase and other 25 plants aspartic proteinase synthase andamino acid in GenBank

图7 26 种植物Aspartic proteinase 合成酶基因氨基酸序列构建的N-J 树Fig.7 Constructed N-J tree of amino acid sequences of aspartic proteinase from 26 plants
3.3.2.2 跨膜结构的预测
Ms Aspartic proteinase 编码的蛋白质跨膜结构可以通过在线分析软件TMHMM 2.0 进行分析预测,结果见图9。
结果显示,天女木兰的天冬氨酸蛋白属于跨膜蛋白。

图8 疏水性的预测分析Fig.8 Prediction analysis of hydrophobicity

图9 跨膜结构域的预测Fig.9 Prediction of transmembrane domain
3.3.2.3 信号肽位点的预测分析
在线分析软件SignalIP 4.0 可以预测分析天女木兰天冬氨酸蛋白的信号肽,结果如图10 所示。
由图10 可知,天女木兰天冬氨酸蛋白酶基因编码的蛋白在氨基酸的第26 位C 值最大,S 值陡峭,Y 值最高峰,为信号肽剪切位点,属于分泌蛋白。
3.3.2.4 三维结构的构建与分析
将天女木兰的天冬氨酸蛋白酶同Phyre2 中其他同源的天冬氨酸蛋白酶已知的蛋白结构作对比后创建模型(图11),该蛋白由二级结构和三维结构覆盖,覆盖率高达84%,可信度为100%,再一次证明了所克隆得到的基因是天女木兰天冬氨酸蛋白酶基因。

图10 信号肽的预测Fig.10 Prediction of signal peptide

图11 蛋白质三维结构建模Fig.11 Modeling of three-dimensional protein structure
4 结论与讨论
4.1 RACE 克隆全长
RACE 技术在获取各物种基因的全长方面得到了广泛的应用。本研究以RACE 技术为核心技术获取天女木兰天冬氨酸蛋白酶基因,在获取中间片段的基础上,通过设计特异性引物和RACE需要的5′ 与3′ 端接头引物,在优化的PCR 扩增条件以及为防止其他杂带的产生而采取的巢式扩增技术下,成功获得了天女木兰种子天冬氨酸蛋白酶基因的5′ 与3′ 端,具有5′ 与3′ 端应有的特征外,同时通过DNAMAN 软件可以正确的拼接,得到的全长序列与其他已公布的天冬氨酸蛋白酶基因具有非常高的同源性,证实了克隆的正确性与完整性。
虽然RACE 技术在植物全长cDNA 的克隆上取得了很大的进展,但是科研人员在实际的实验操作上依然存在着大量的问题与技术难题,如RACE 实验的基础是一段已知的基因片段才能够向两端延伸,而中间片段的获得也需要科研工作者大量的研究实验,具有一定的难度;RACE 对于RNA、cDNA、引物、扩增条件都具有相当高的要求,在实验的过程中每一步都需要注意操作的精细度以获得高质量的实验材料以及实验环境;RACE 的3′ 端扩增相对简单,而5′ 端扩增难度较大,需要的技术较为复杂,即使各部分酶反应顺利,也有可能会产生大量的非特异性条带。因此,需要针对不同的物种、不同的基因加以摸索研究,不断优化反应条件,才能成功地利用RACE 技术获取目的基因的全长cDNA。
4.2 天冬氨酸蛋白酶的生物信息学分析
天冬氨酸蛋白酶基因总长1 798 bp,序列中包含一段从109 bp ATG 起始到1 653 bpTAA 终止的完整的ORF 区域,总长1 545 bp,可以编码514个氨基酸残基。天女木兰天冬氨酸蛋白酶基因是否存在基因家族并不明确,该基因在天女木兰的时空表达模式在本研究中没有体现,下一步还需要进行深入的研究。
利用分子生物学技术对天女木兰种子休眠及其解除机理深入研究,利用同源克隆法RACE 技术克隆得到天冬氨酸蛋白酶基因全长,结合生物分析技术来对天冬氨酸蛋白酶的结构与功能以及亚细胞定位,有利于后续遗传转化中掌握天冬氨酸蛋白酶基因在天女木兰植物体中的作用,有助于全面地研究天女木兰种子休眠解除机理,为系统地探索天女木兰种子休眠机制提供了理论基础,找到了新的窗口与靶点。
猜你喜欢
河北省科学院学报(2023年4期)2023-08-29 13:10:04
教学月刊(中学版)(2022年12期)2022-12-27 00:11:27
青年生活(2020年6期)2020-03-28 01:25:37
幽默大师(2019年5期)2019-05-14 05:39:58
出版人(2019年2期)2019-03-16 04:31:34
山东化工(2017年22期)2017-12-20 02:43:37
花卉(2017年3期)2017-07-20 11:05:54
丝路艺术(2017年6期)2017-04-18 13:58:26
化工设计通讯(2017年10期)2017-03-02 03:24:02
听力学及言语疾病杂志(2015年5期)2015-12-24 01:47:05
