硝酸镓对耐碳青霉烯肺炎克雷伯菌生长及其生物膜的抑制作用
2019-05-05 03:00顾毅刘周徐方明盛男赵亚运孙凯莉刘艳艳叶英李家斌
中国抗生素杂志 2019年4期
顾毅 刘周 徐方明 盛男 赵亚运 孙凯莉 刘艳艳 叶英 李家斌
(安徽医科大学第一附属医院感染病科,安徽省细菌耐药监测中心,合肥 230023)
肺炎克雷伯菌(Klebsiella pneumoniae,KP)属于肠杆菌科克雷伯菌属、为革兰阴性菌,是临床重要的条件致病菌,可感染人体全身尤其是呼吸和泌尿系统,引发肺炎、脑膜炎、肝脓肿和败血症等疾病。随着抗生素的不恰当使用,其耐药率逐年上升。其中耐碳青霉烯肺炎克雷伯菌(carbapenem-resistantKlebsiella pneumoniae,CRKP)在全球广泛分布并呈不断增加的态势[1]。CRKP常携带大量耐药基因,对临床常用的抗菌药物呈现多重耐药,仅对多黏菌素、替加环素敏感率较高[2]。研究表明,对替加环素耐药的CRKP及携带mcr-1基因对多黏菌素耐药CRKP已经出现,给临床治疗带来了巨大挑战[3-6]。在自然界,超过90%的细菌均以生物被膜(biofilm,BF)的形式存在,当细菌BF形成后,其耐药性相比于浮游菌会提高10~1000倍[7],这使耐药菌的被清除率进一步降低,人体一旦被耐药菌感染便成为一件岌岌可危的事情。因此,寻找能有效抑制细菌及其BF形成的新型抗菌药物已成为防治KP的重要研究内容。庆大霉素(gentamicin,GEN)作为治疗KP导致的感染性疾病的常用抗菌药物,近年来已有文献报道,GEN对铜绿假单胞菌、葡萄球菌等病原微生物的BF形成有着抑制作用[8-10]。镓作为一种半金属元素,临床上主要用于肿瘤的显像检查和治疗[11-12],近年来其抑菌作用被研究人员关注。相关研究表明,硝酸镓(gallium nitrate,GaN)对鲍曼不动杆菌、大肠埃希菌的生长及生物膜形成有较好的抑制或清除作用[13-14]。本实验以GEN为对照组,研究GaN对CRKP生长及其生物膜形成的影响,为有效治疗耐药菌引起的感染提供有效的方法。
1 材料与方法
1.1 材料
1.1.1 菌株
本研究的82株CRKP均分离于安徽省细菌耐药监测中心监测网所属34家医院2015—2016年度的住院和门诊患者的各类临床标本。其中来源于痰标本63株,尿液标本11株,血液标本7株,分泌物标本1株。标准菌株肺炎克雷伯菌ATCC BAA-1705、肺炎克雷伯菌ATCC BAA-1706为质控菌株(保存于安徽省细菌耐药监控中心)。
1.1.2 仪器与试剂
VITEK-2 Compact细菌鉴定仪及配套GN鉴定卡(法国Biomerieux公司);ZEISS LSM 510 META型激光共聚焦扫描显微镜(德国Chua公司);硝酸镓(美国Sigma公司);M9CA肉汤培养基、结晶紫染色液(北京陆桥股份有限公司);96孔、24孔细胞培养板(美国Corning公司);细胞爬片(无锡耐思生物科技有限公司);细菌培养电热恒温培养箱(上海精宏公司);电子天平(上海名桥精密科学仪器公司)、移液器(德国Eppendorf公司)。抗菌药物标准品:亚胺培南(imipenem,IPM)、美罗培南(meropenem,MEM)、氨曲南(aztreonam,AZM)等均购自于中国食品药品检定研究院。
1.2 方法
1.2.1 菌株鉴定与药敏试验
所有菌株应用VITEK-2 Compact全自动微生物分析系统进行鉴定。所检测的抗菌药物包括氨苄西林、头孢哌酮/舒巴坦、哌拉西林/三唑巴坦、头孢吡肟、头孢曲松、头孢西丁、头孢他啶、头孢唑肟、氨曲南、亚胺培南、美罗培南、阿米卡星、庆大霉素、环丙沙星、左氧氟沙星、加替沙星及氯霉素应用琼脂倍比稀释法进行药敏试验。
1.2.2 GaN对CRKP的最小抑菌浓度(minimal inhibitory concentrations,MIC)测定
应用微量肉汤稀释法检测:GaN对CRKP的MIC值,具体步骤如下:各实验菌接种于琼脂平板上,37℃过夜,取单个菌落置生理盐水配制约1×108~2×108CFU/mL。GaN用M9培养基配制成浓度为16μmol/mL溶液,过滤除菌,加入到无菌的96孔板内,每排第1孔加260μL,第2~10孔每孔分别加入130μL培养基,然后从第一个孔移出130μL到第二个孔,以此类推成倍比稀释,每株菌设4个复孔。同时设无菌液的培养基为阴性对照和不含药液的菌液为阳性对照。将培养板置于37℃培养24h,以实验孔中无细菌生长的最低药物浓度为MIC,实验重复3次,结果取重复次数最多的值。
1.2.3 BF强阳性筛选
采用结晶紫染色法[15]。用琼脂培养基过夜培养待测菌株,菌液稀释为2×108CFU/mL,按1:100取菌液转移至96孔板中的130μL M9培养基中,将82株CRKP放于37℃培养箱静置24h,吸取菌液,用无菌PBS清洗游离细菌,待完全干燥,每孔加入0.2%结晶紫染色液140μL,染色30min,用缓慢持续的流水将多余的染料冲洗干净,待完全干燥后每孔加入95%乙醇150μL,使其与生物膜结合的结晶紫完全溶解,用酶标仪检测吸光度值(A570),基于临界值Ac,菌株BF可分为弱(Ac
1.2.4 BF抑制检测
根据生物膜筛选及其相应的MIC结果,选择1/2MIC的GaN作用于强阳性BF菌株。按“1.2.3”项的方法,检测31株BF强阳性的CRKP在1/2MIC药液浓度下生物膜形成情况。37℃静置培养24h,然后染色,检测A570吸光度值。实验重复4次,结果取平均值。
1.2.5 GaN与GEN分别对CRKP生物膜抑制率的检测
从31株BF强阳性的CRKP中随机挑选出20株菌,应用微量肉汤稀释法检测GEN对CRKP的MIC值。按“1.2.3”项的方法,检测20株菌分别在1/2MIC的GaN及1/2MIC的GEN药物浓度下生物膜形成情况。实验重复4次,结果取平均值。以不加药物处理的含菌的M9培养基为阳性对照,以单纯M9培养基为空白对照。生物膜抑制率=(1-处理组A570/对照组A570)×100%[14]。
1.2.6 激光共聚焦扫描显微镜(confocal laser scanning microscope,CLSM)三维生物膜结构成像
将12mm无菌细胞爬片放入标准24孔微量板中,使孔板中含菌液的培养基的GaN终浓度分别为0、1MIC和1/2MIC,37℃静置培养24h。吸取菌液,3.7%的甲醛固定,无菌PBS清洗游离细菌3次,0.1%吖啶橙避光染色5min[13],再用无菌蒸馏水清洗多余染料,自然干燥。利用CLSM Zeiss LSM880观测BF形成情况。参数设置如下:吖啶橙激发光波长为488nm,发射光波长512~654nm,物镜20倍。利用Zeiss ZEM 2012软件包获取共聚焦图片,通过Imaris 7.0软件制作三维重组的BF成像图片。实验重复3次。
1.2.7 统计学处理
CRKP对抗菌药物的耐药率应用WHONET 5.6 软件进行统计分析,折点解读参照CLSI 2016标准[16],生物膜以(平均值±标准差,±s)的形式表示,生物膜抑制率以百分比(%)的形式表示,SPSS 17.0软件进行两独立样本均数的t检验,以P<0.05为差异有统计学意义。
2 结果
2.1 药敏结果
安徽地区82株临床分离的CRKP对β-内酰胺类抗生素(氨苄西林、头孢曲松)和单环酰胺类药物(氨曲南)的耐药率均为100.0%,对头霉素类药物(头孢西丁)和β-内酰胺+酶抑制剂类药物(头孢哌酮/舒巴坦、哌拉西林/三唑巴坦)的耐药率较高,分别为95.1%、96.3%和97.6%,对喹诺酮类药物的耐药率为82.9%~98.8%,仅对氨基糖苷类药物中阿米卡星的耐药率较低,为59.8%(表1)。
2.2 GaN对CRKP的MIC值测定
应用微量肉汤稀释法检测GaN对82株CRKP的抑菌情况,其MIC值范围为0.0625~2μmol/mL,其中MIC50和MIC90分别为0.25和0.5μmol/mL。菌株MIC值分布情况见图1。
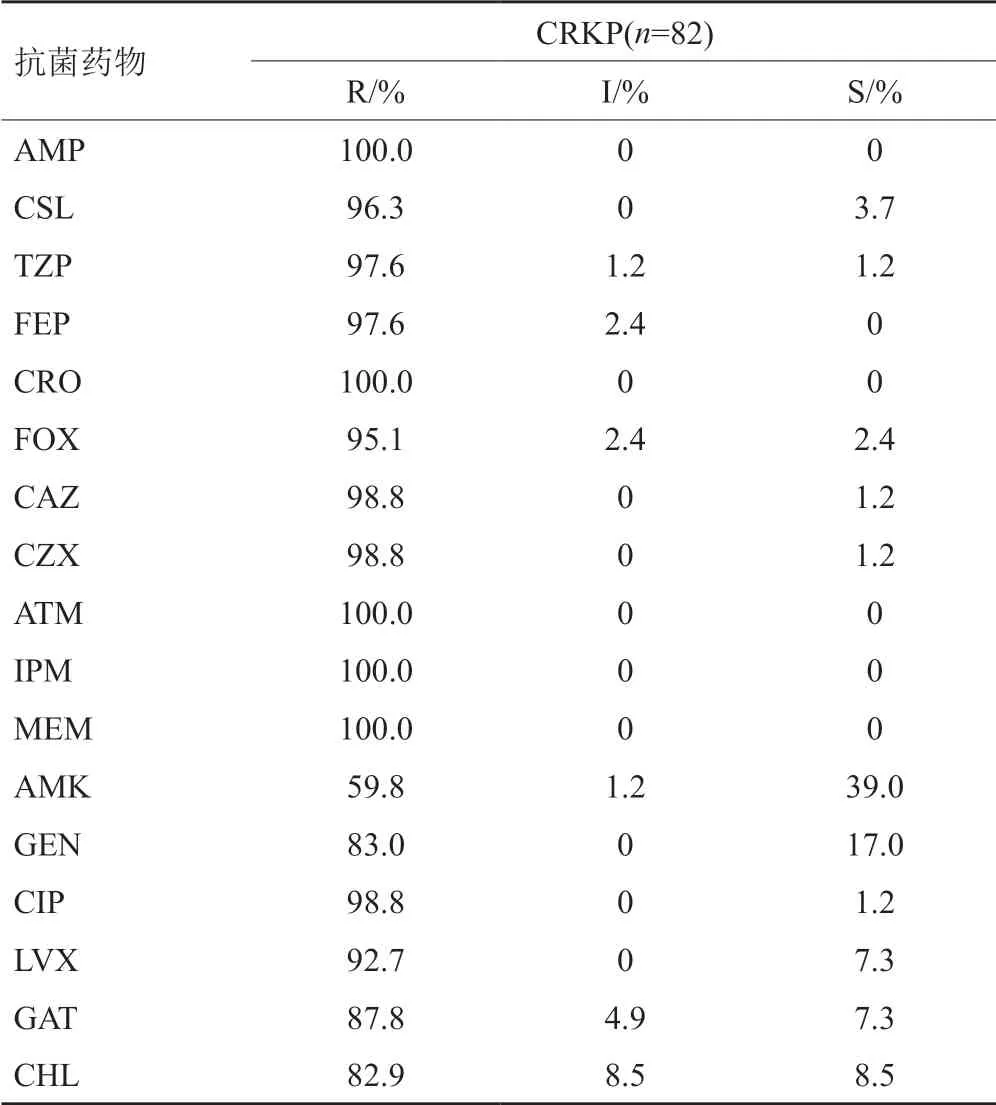
表1 17种抗菌药物对82株实验菌的药敏结果(%)Tab.1 Susceptibility results of 17 antibiotics for 82 isolates(%)
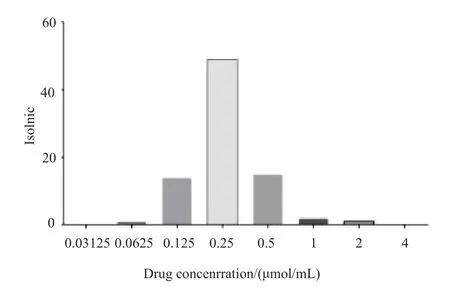
图1 GaN对82 株CRKP的MIC值的分布情况Fig.1 The distribution of MIC of Gallium nitrate for 82 isolates
2.3 BF强阳性筛选及CRKP生物膜抑制检测
通过检测,82株CRKP中BF强阳性(>4Ac)的CRKP为31株(37.8%)。1/2MICGaN对31株CRKP的BF作用结果,由SPSS 17.0软件统计分析,结果显示t=18.471,P<0.001,在α=0.05的检验水准下,差异有统计学意义,即1/2MIC GaN可以有效抑制CRKP的BF形成(表2)。
2.4 BF抑制率的检测
通过检测,GaN对实验菌株的BF抑制率为37.8%~60.9%,GEN对实验菌株的BF抑制率为32.4%~53.3%。两组BF抑制率结果,由SPSS 17.0软件统计分析,结果显示t=3.639,P<0.05,在α=0.05的检验水准下,差异有统计学意义,即GaN和GEN对实验菌株的BF形成的作用有差别,且GaN的作用优于GEN(图2)。
2.5 BF三维结构成像图分析
用CLSM观察CRKP在不同GaN浓度下BF三维结构并测量其厚度,由图显示:没有加入GaN的菌株形成了紧凑密集且厚的BF(A),平均厚度为(250±0.7)nm;加入1/2MIC的GaN形成的BF较为稀疏,为散在的小斑块(B),平均厚度为(90±0.4)nm;当GaN达到MIC值时,BF形成更为稀少,为散在的小点(C),平均厚度为(27±0.3)nm。由此显示,GaN对CRKP的BF形成有着良好的抑制作用(图3)。
3 讨论
细菌BF是多个细菌不可逆的黏附于机体或物体表面,被自身细胞分泌的基质包被而形成的细菌聚集膜样物[17]。肺炎克雷伯菌有着丰富的荚膜,并依靠荚膜多糖和细菌分泌的黏附分子及胞外DNA极易在物体表面形成BF结构[18]。BF一旦形成,膜内细菌可被不断释放,造成感染的迁延不愈与反复发作[7]。BF不仅可形成屏障来降低抗菌药物的渗透能力,而且膜内细菌由于营养物质的缺乏,代谢水平下降,对抗菌药物的敏感性也随之降低[19]。因此,积极寻找有效抑制BF形成的新型抗菌药显得尤为重要。
表2 GaN对31株菌的MIC值及BF的A570值(n=4,±s)Tab.2 MIC of gallium nitrate and biofilm A570 for 31 isolates (n=4,±s)
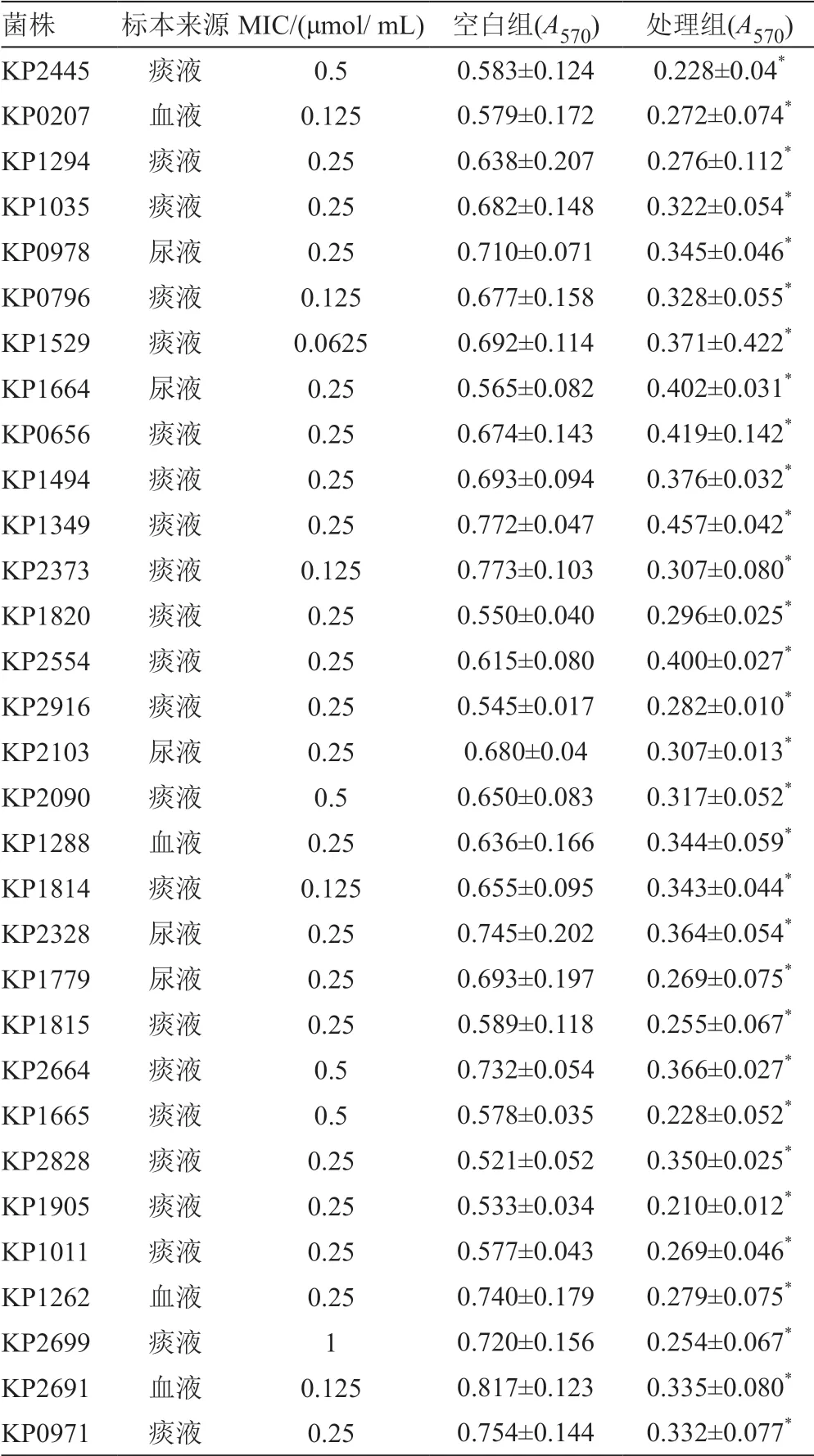
表2 GaN对31株菌的MIC值及BF的A570值(n=4,±s)Tab.2 MIC of gallium nitrate and biofilm A570 for 31 isolates (n=4,±s)
注:空白组代表用药前BF的A570值;处理组代表用药后BF的A570值;与空白组比较,“*”P<0.05
菌株 标本来源MIC/(μmol/ mL) 空白组(A570) 处理组(A570)KP2445 痰液 0.5 0.583±0.124 0.228±0.04*KP0207 血液 0.125 0.579±0.172 0.272±0.074*KP1294 痰液 0.25 0.638±0.207 0.276±0.112*KP1035 痰液 0.25 0.682±0.148 0.322±0.054*KP0978 尿液 0.25 0.710±0.071 0.345±0.046*KP0796 痰液 0.125 0.677±0.158 0.328±0.055*KP1529 痰液 0.0625 0.692±0.114 0.371±0.422*KP1664 尿液 0.25 0.565±0.082 0.402±0.031*KP0656 痰液 0.25 0.674±0.143 0.419±0.142*KP1494 痰液 0.25 0.693±0.094 0.376±0.032*KP1349 痰液 0.25 0.772±0.047 0.457±0.042*KP2373 痰液 0.125 0.773±0.103 0.307±0.080*KP1820 痰液 0.25 0.550±0.040 0.296±0.025*KP2554 痰液 0.25 0.615±0.080 0.400±0.027*KP2916 痰液 0.25 0.545±0.017 0.282±0.010*KP2103 尿液 0.25 0.680±0.04 0.307±0.013*KP2090 痰液 0.5 0.650±0.083 0.317±0.052*KP1288 血液 0.25 0.636±0.166 0.344±0.059*KP1814 痰液 0.125 0.655±0.095 0.343±0.044*KP2328 尿液 0.25 0.745±0.202 0.364±0.054*KP1779 尿液 0.25 0.693±0.197 0.269±0.075*KP1815 痰液 0.25 0.589±0.118 0.255±0.067*KP2664 痰液 0.5 0.732±0.054 0.366±0.027*KP1665 痰液 0.5 0.578±0.035 0.228±0.052*KP2828 痰液 0.25 0.521±0.052 0.350±0.025*KP1905 痰液 0.25 0.533±0.034 0.210±0.012*KP1011 痰液 0.25 0.577±0.043 0.269±0.046*KP1262 血液 0.25 0.740±0.179 0.279±0.075*KP2699 痰液 1 0.720±0.156 0.254±0.067*KP2691 血液 0.125 0.817±0.123 0.335±0.080*KP0971 痰液 0.25 0.754±0.144 0.332±0.077*
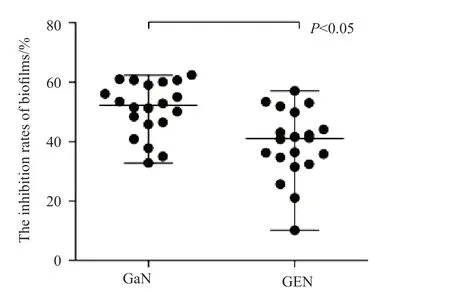
图2 GaN与GEN对CRKP生物膜的抑制率(%)Fig.2 The effect of GaN and GEN on the biofilm formation of CRKP (%)

图3 CLSM检测GaN作用下的BF形成Fig.3 Three-dimensional reconstructions of CLSM images of biofilm formation
本研究中收集了安徽省地区34家医院临床分离的82株CRKP,均为多重耐药菌,给临床治疗带来了困难,需加以足够的重视。本研究中,首先测定了GaN对实验菌株的MIC值。从分布来看,MIC值范围为0.0625~2μmol/mL,其中MIC50、MIC90分别为0.25和0.5μmol/mL。为进一步探索GaN对BF形成的抑制作用提供了前提条件。
细菌BF一旦形成就很难被清除,因此如何有效地抑制BF形成至关重要。本次研究中,临床分离菌株为82株,BF强阳性菌株所约占37.8%(31/82),由此可见CRKP的BF形成的严重性。Kaneko等[20]研究发现GaN对铜绿假单胞菌的生长及其BF形成有抑制作用,不仅可以杀灭浮游菌,而且对已经形成BF的细菌也同样有效。本研究中采用的培养基为低铁培养基,在这种铁营养缺乏的生存条件下,细菌自身会产生一种小分子量铁螯合物,进而从外界摄取必要的铁元素以供自身需要。实验采用1/2MIC GaN作用于31株BF强阳性的CRKP菌株,处理前后BF形成数据结果有统计学意义,表明GaN对BF形成有较好的抑制作用。本实验中,GaN对CRKP的BF抑制率与GEN对CRKP的BF抑制率有统计学意义,GaN对生物膜的抑制率优于GEN。CLSM三维结构成像图直观显示,BF在1/2MIC与1MIC GaN条件下形成的量有明显的降低,进一步证明了GaN对BF形成有着较好的抑制作用。
本研究中仅对GaN在体外的抗菌作用进行了试验,并没有检测其在体内的抗菌作用。但有文献表明,GaN可以通过扰乱结核分枝杆菌的铁代谢来发挥抑菌作用,并且在小鼠肺结核模型中抑制了细菌的生长[21]。同时也有文献报道,给志愿者口服500mg镓制剂,血清药物浓度为1μg/mL(约为0.0039μmol/mL),远达不到抑菌浓度[22]。这可能是由于胃肠道中不溶性镓氢氧化物的形成,降低了镓的生物利用度[23]。在进一步的研究中,我们将使镓与其有协同作用的抗菌药物联用,从而提高镓在体内的生物活性,提高其应用于治疗临床耐药菌感染的潜力。并且,GaN已通过美国食品和药物管理局(FDA)批准用于治疗肿瘤性高钙血症,具有很低的组织毒性[18]。
综上所述,本研究发现CRKP形成BF的能力具有普遍性及严重性,伴随着其BF的形成,其耐药性也普遍提高。新型抗菌剂GaN对CRKP的BF形成有着较好的抑制作用,此结果将为GaN用于临床抗感染治疗奠定基础。由于GaN具有抗菌活性,其与生物材料结合,从源头阻止或减少细菌在侵入性医疗器材上的黏附与定植,进而防止或降低细菌对宿主的感染,不失为目前抗感染的有效之举。
猜你喜欢
中国合理用药探索(2022年3期)2022-11-25
浙江农业学报(2022年7期)2022-07-28
现代农村科技(2022年1期)2022-01-21
浙江临床医学(2021年12期)2022-01-13
中国循证心血管医学杂志(2021年10期)2021-11-05
现代临床医学(2021年5期)2021-11-02
昆明医科大学学报(2021年4期)2021-07-23
纺织科技进展(2021年5期)2021-07-22
食品界(2021年7期)2021-07-19
商品与质量(2020年47期)2020-12-18