野野村放线菌ATCC 39727的双组份信号转导系统的生物信息学分析
2019-05-05 03:00岳雪王帅李大成王勇健董惠钧
中国抗生素杂志 2019年4期
岳雪 王帅 李大成 王勇健 董惠钧,*
(1 聊城大学药学院,聊城 252000;2 聊城大学生物制药研究院,聊城 252000)
野野村放线菌(Nonomuraeasp.ATCC39727)可以产生糖肽类抗生素达巴万星前体A40926,经酯化、酰胺化、水解和成盐等步骤合成达巴万星。达巴万星主要用于治疗革兰阳性菌引起的皮肤和皮肤组织感染,抗菌活性优于同类药物万古霉素和替考拉宁[1]。野野村放线菌基因组已测序完成,大小约11.85Mb,基因组上分布有33个次级代谢生物合成基因簇。糖肽抗生素达巴万星前体A40926的合成基因簇dbv包含37个开发阅读框(open read frame,ORF),大小为71kb,指导氨基酸前体物先合成七肽骨架,经环化和后修饰生成A40926[2]。
相对于高等动植物,微生物有着超强的环境适应能力。微生物这种超强能力的遗传基础是其拥有复杂多样的信号转导系统,其中最重要的信号转导系统是双组分信号转导系统(two-component signal transduction system,TCS)。TCS目前只在细菌、古生菌和植物中有发现,而在人类和其他哺乳动物体内尚未发现。通常TCS系统由组氨酸激酶(histidine kinase,HK)和应答调节蛋白(response regulator,RR)构成。HK中含有重复序列的N端胞外感受域和保守的C端胞内传导域(HisKA),由N端胞外感受域感应外界的特定环境刺激信号。RR通常包括一个保守的N端接受域和一个多变的C端输出域(REC)组成,通过天冬氨酸残基调控细胞内的一系列应答反应[3]。大多数细菌体内都存在数量不等的TCS,它们调控细菌的趋化性、感知渗透压、pH、金属离子、形态分化、营养物质代谢以及次级代谢产物的生物合成等诸多生理过程。
放线菌具有复杂的次级代谢产物和网络。与之相对应,放线菌中也具有丰富多样的TCS系统,两者之间息息相关。本文通过生物信息学方法对Nonomuraeasp.ATCC39727中TCS的分类、保守结构域、进化关系以及功能进行分析,旨在深入解析ATCC39727的生长和代谢相关基因的表达和调控,为进一步解析A40926合成代谢和调控,构建高产工程菌株提供理论支撑。
1 材料与方法
1.1 ATCC39727基因组中双组份信号系统预测和结构功能分析
根据双组份信号系统HK和RR蛋白保守结构域分析ATCC39727基因组中可能存在的双组份系统。本文中ATCC39727基因组序列信息来自NCBI数据库。ATCC39727双组份信号系统的鉴定方法:(1)利用 Pfam 数据库中保守的HK和RR结构域HATPase_c(Pfam00512)和Response_reg(Pfam00072)对ATCC39727基因组进行搜索比对,分别鉴定基因组中的HK和RR;(2)用NCBI的Blast P对得到的HK/RR序列进行在线比对。
双组份信号系统的确认标准:(1)HATPase结构域应该位于HK蛋白的C端;(2)HisKA位于HATPase 结构域的前面,HATPase_c的上游具有二聚化或磷酸化结构域DHp;(3)RR的N端含有磷酸受体,Response_reg结构域位于RRs的C-末端。最后,参照 Williams和Whitworth的方法[4]对成对的TCS(HK-RR对)、融合HK和孤儿HK/RR进行鉴定。如果HK和RR 位于同一操纵子上,则该HK和RR成功配对为成对 TCS;如果HisKA和REC结构域同时出现在一个蛋白上,那么该蛋白定义为融合HK;如果HK/RR在同一操纵子上找不到对应的RR/HK,则该HK/RR定义为孤儿HK/RR。
1.2 双组份信号进化树和调控网络构建
分别将ATCC39727基因组中所有HK和RR蛋白序列导入Mega 6.0软件中,首先进行Clustal W同源比对,然后选用NJ法构建进化树,并进行Boot-strap验证,重复次数设置为1000。
利用Pfam、SMART和BLASTp进行保守结构域分析,并用UniprotKB数据库进行辅助验证,结合文献报道,预测TCS各组分的功能,并在此基础上构建ATCC39727 TCS信号转导网络。
2 结果与分析
2.1 ATCC39727基因组中TCS分布
与其他微生物相比,放线菌具有复杂的形态分化和丰富的次级代谢网络。与之相对应,放线菌也含有丰富的TCS系统,与大肠埃希菌和枯草芽孢杆菌平均25个成对TCS相比,天蓝色链霉菌和野野村放线菌则分别含有67对和60对TCS(表1)。进一步对比发现,野野村放线菌TCS系统中的HK和RR蛋白更加保守,几乎没有特异性结构域。通过搜索比对和分析发现,ATCC39727基因组中含有丰富的TCS基因,共有213个相关编码基因。进一步分析其中112个基因编码HK蛋白,包含60个TCS HK-RR和52个孤儿HK,99个基因编码TCS RR蛋白,包含60个HK-RR和39个孤儿RR,另外还有2个融合HK(表2)。尽管在细菌基因组中TCS基因所占比例不大(通常低于5%),但在调控微生物生理生化代谢中发挥重要作用。
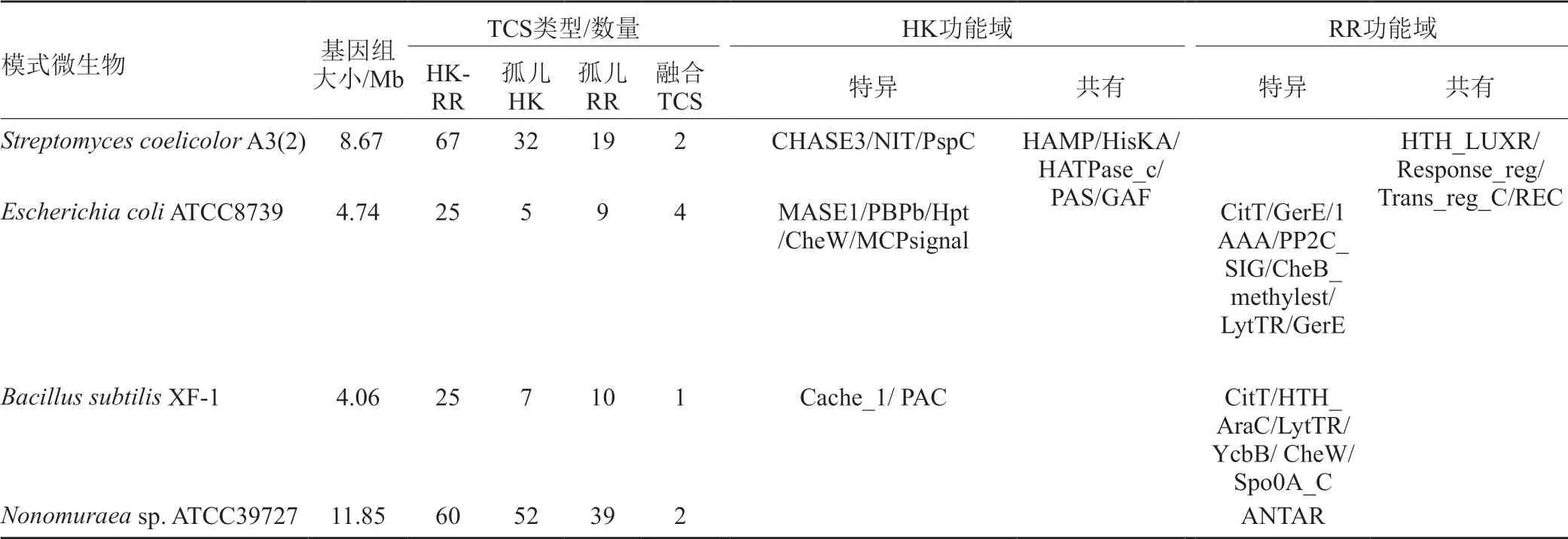
表1 几种模式微生物TCS系统比较Tab.1 Comparison of TCSs from different model microorganisms
2.2 ATCC39727基因组HKs蛋白分类
从ATCC39727基因组中搜索到所有可能的HK蛋白后,用Pfam、SMART和NCBI进行蛋白结构域分析。TCS的HK蛋白通常为跨膜蛋白,包含胞外信号感应域(sensing domain)、胞内信号传递域(transmitter domain)和ATP酶域(ATPase domain)[5]。根据结构域的组成情况,ATCC39727中112个HK蛋白分为14组,其中65个HK蛋白含有多个重复序列和HATPase_c结构域,35个HK蛋白含有HAMP、HisKA和HATPase_c3个结构域,其余12个HK蛋白含有各异的结构域组合,感应结构域主要有HAMP、GAF和PAS(图1)。HATPase_c结构域是胞内负责将ATP的磷酸基团转移到组氨酸上的催化结构域,HisKA则是接受磷酸的受体结构域,HAMP结构域是受体组氨酸激酶和甲基接受蛋白的细胞质螺旋连接结构域,PAS结构域与感应外界氧、氧化还原电位和光的变化有关,GAF结构域主要对环核苷酸结合发挥作用,例如cGMP和cAMP等。
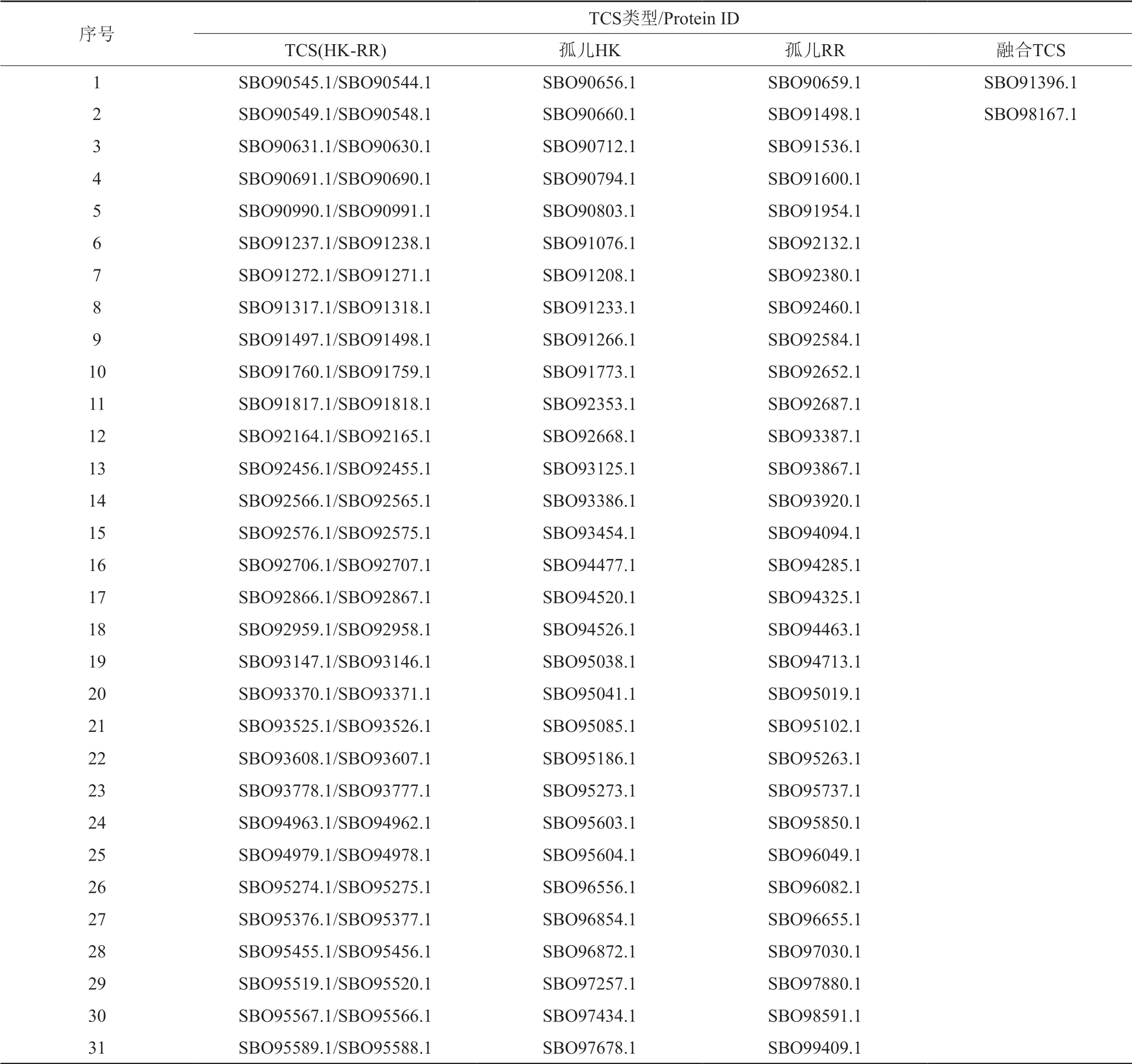
表2 Nonomuraea sp.ATCC39727基因组中双组份系统Tab.2 Two-component signal transduction systems in Nonomuraea sp.ATCC39727
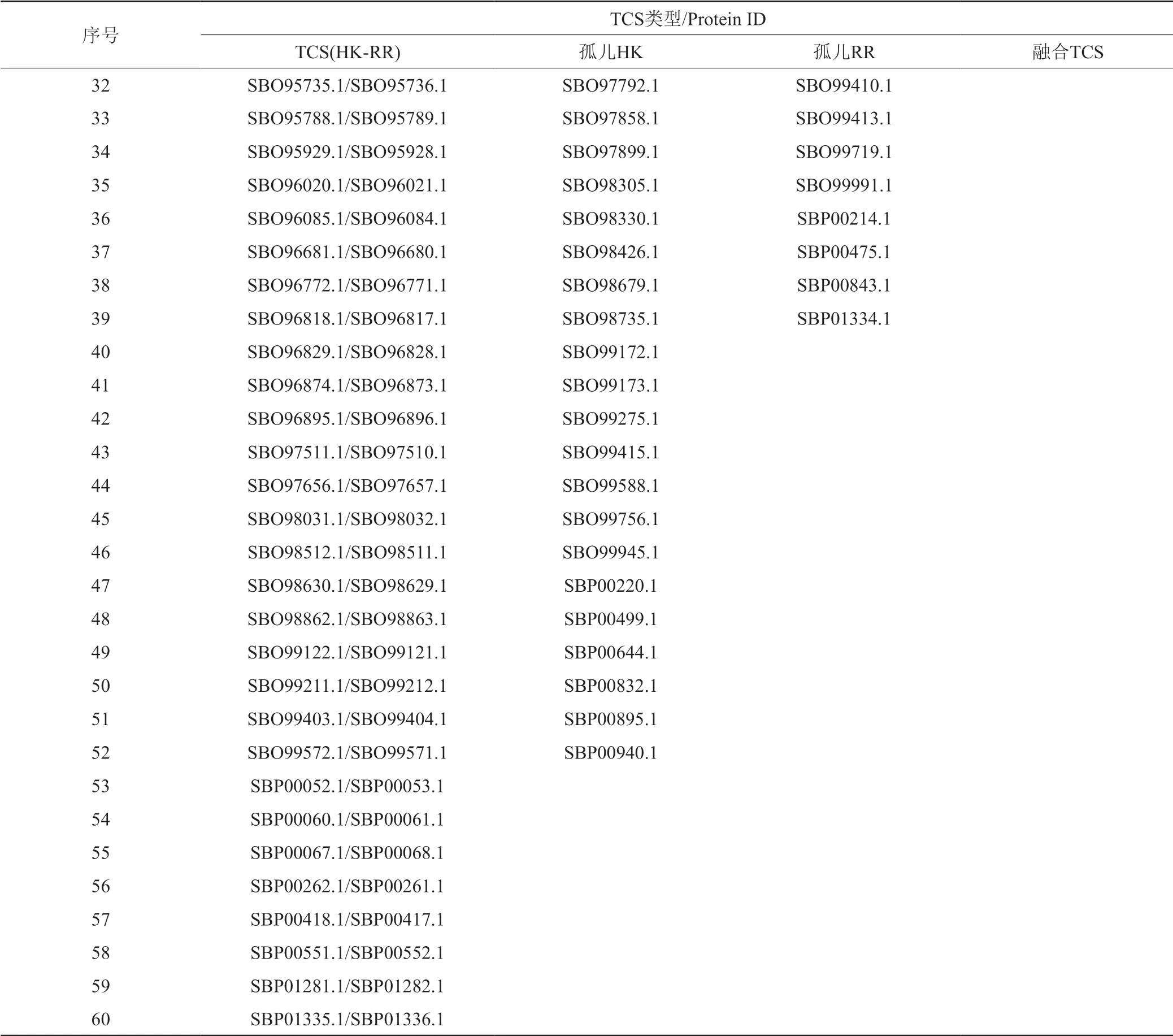
续表2
根据结构域功能可将HK蛋白分为胞质感应HK、胞外感应HK和膜感应HK。含有PAS和GAF结构域的HK为胞质感应HK,含有跨膜和HAMP结构域的HK为胞外感应HK,仅含有跨膜结构域和HATPase_c的为膜感应HK。从图1可以看出,ATCC39727中的HK蛋白绝大多数是胞外感应和膜感应类型。
为了解ATCC39727基因组中TCS基因的相互关系,分别对ATCC39727菌株中所有的HK和RR蛋白序列进行比对和进化分析,发现行使不同功能的HK蛋白之间的序列保守性弱,呈现多样性,而RR蛋白的保守性则要强一些。另外从HK-RR来比较,同源相近的HKs对应的RRs并不同源,反映了TCS进化的多样性(图2)。
2.3 ATCC39727基因组RRs蛋白分类
与HK相比,RR在长度和组成上都相对保守,它由N端含有Asp残基的高度保守的信号接受域(receiver domain)以及C端的效应结构域(effector domain)两部分组成。根据结构域的差异,ATCC39727中的RR蛋白可以分为6类(图3)。其中含有REC和HTH_LUXR结构域的RR蛋白最多,共有57个;其次有24个RR蛋白含有REC和Trans reg_c结构域;RR蛋白中最多的结构域是REC结构域(趋化蛋白CheY同源的受体结构域),ANTAR结构域是RNA结合输出域,HTH_LUXR 结构域起到结合DNA和转录激活的作用。

图1 TCS HKs蛋白结构域类型Fig.1 Conserved domain compositionof TCS HK proteins
2.4 ATCC39727 TCS功能预测
根据ATCC39727基因组中HKs和RRs蛋白的保守结构域和相对位置,发现了60对不同的TCS。根据Blast P结果注释和相关文献,对60对TCS的功能进行了预测和分析(表3)。结果表明,ATCC39727中的TCS同其他微生物中的功能类似,主要包括细胞壁和细胞膜的合成和调节,例如VraS/VraR(调节肽聚糖合成)、LiaS/LiaR(与细胞壁压力和抗菌肽调节相关)、MtrA/MtrB(调节渗透压)、DesR/DesK(调节细胞膜流动性);调节细胞形态分化,例如BaeS/BaeR(调节DNA起始复制)、ArlS/ArlR(调控接合和自溶)、DevR/DevS(启动细胞休眠)、DegS/DegU(调控生物膜形成)、PrrB/PrrA(调控细胞早期增殖)、YdfH/YdfI(参与细胞分化);参与营养元素代谢,例如NarS/NarL(参与硝酸盐代谢)、PhoP/PhoR(参与磷代谢)、NreB/NreC(参与硝酸盐异化和亚硝酸盐代谢)、NarX/NarP(调节氮代谢);调节重金属离子平衡和抗性,例如CzcS/CzcR(调节Zn代谢)、CopS/CopR(与铜抗性基因有关)、CutS/CutR(调节铜代谢)、IrlR/IrlS(与重金属抗性有关)、PcoS/PcoR(活化铜抗性基因)、TcrY/TcrX(与铁缺乏有关);与细胞抗性和毒力相关,例如YvfT/YvfU(与细菌毒力有关);调节细胞呼吸和途径代谢,例如ResD/ResE(调控有氧和无氧呼吸)、MprB/MprA(全局调控子)、UhpC/UhpB(调控己糖磷酸化)。
2.5 ATCC39727中TCS调控网络
通过对ATCC39727基因组中的TCS进行功能预测,并结合相关文献,构建了ATCC39727中部分TCS调控网络。TCS调控网络的显著特征是相互交叉,呈现“一对多或多对一”的级联反应(图4)。例如在缺氧情况下,ResD诱导YclJK双组份表达,进一步YclJK正调控YsfCD表达,后者又同时被DesSU负调控。又例如在磷酸盐缺乏时,PhoR/PhoP系统激活,诱导另一组磷代谢相关的双组份系统PhoA/PhoB,同时会抑制氮代谢相关蛋白GlnA的表达,而GlnA会通过TnRA蛋白与DegS/DegU相互作用。在ATCC39727中存在多个与金属离子代谢相关的TCS。CopS/CopR在环境中铜离子的压力下激活,进而诱导结构基因copABCD表达,copABCD的表达会进一步激活PcoS/PcoR,Pco相关蛋白的表达外排铜离子、抵抗铜离子的毒害。类似的,锌离子的存在会激活CzcS/CzcR,进而诱导czcC基因的表达。CopS/CopR和CzcS/CzcR两个双组份系统都会抑制oprD基因(细胞膜孔道蛋白)的表达。
3 讨论
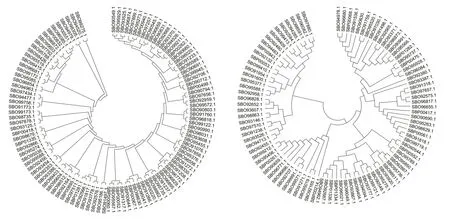
图2 TCS HKs和RRs蛋白进化分析Fig.2 Phylogenetic trees of HK and RR proteins
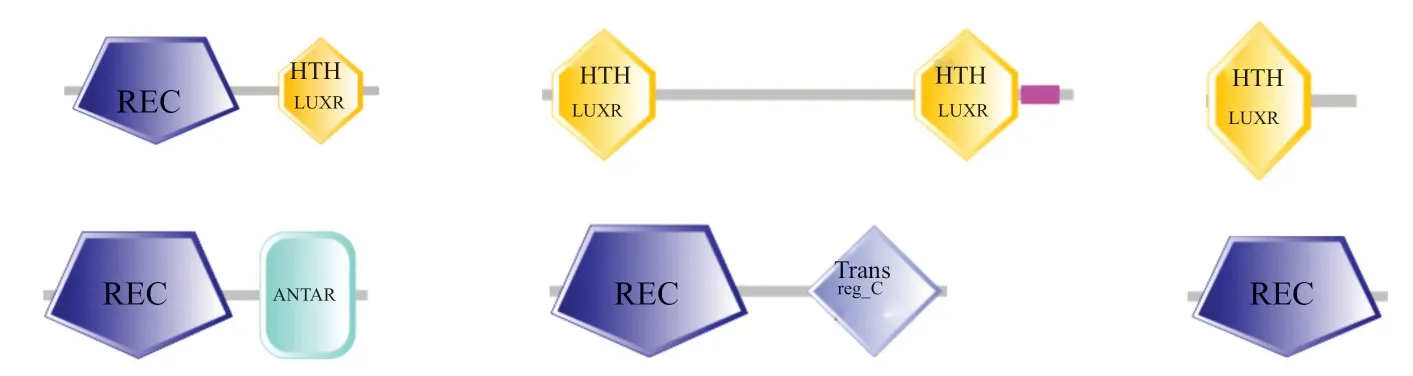
图3 TCS RRs蛋白结构域类型Fig.3 Conserved domain composition of TCSRR proteins
糖肽抗生素是一类重要的抗生素,是临床上治疗耐甲氧西林金黄色葡萄球菌等“顽劣菌株”的最后一道防线。目前糖肽抗生素已经发展到第二代,都是直接来源于微生物的天然产物或衍生物。以A40926为前体的达巴万星是第二代糖肽抗生素中活性最强的,其半衰期长达174h,临床上一周仅给药1次。野野村放线菌是A40926生产菌株,其全基因组已测序完成,基因组大小11.85Mb,基因组上分布有丰富多样的TCS双组份系统,共有213个相关HK和RR编码基因。ATCC39727中的只有极少数TCS系统得到实验验证,其中途径特异性调控基因dbv22(HK)和dbv6(RR)是一对TCS系统,但Grasso等[33]证实dbv6并不影响A40926的表达。尽管TCS系统中典型的HK蛋白含有4个保守结构域,但ATCC39727的HK蛋白的保守结构域有14种组合类型,其中最保守的结构域是HATPase,每一种组合类型都有该结构域,这种组合多样的特征可能是由于其为了更好的适应复杂的环境变化而发生的进化。本文通过生物信息学分析初步构建了部分TCS系统的调控网络。在此基础上,本文将在下一步工作中重点研究ATCC39727中双组份对A40926次级代谢途径的影响。
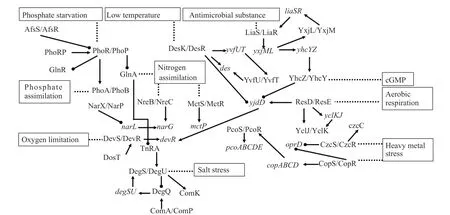
图4 ATCC39727中TCS调控网络Fig.4 TCS regulation network in ATCC39727
猜你喜欢
现代园艺(2022年19期)2022-09-21
军事文摘(2022年16期)2022-08-24
湖北农业科学(2022年11期)2022-07-18
河北农业大学学报(2022年2期)2022-04-26
小读者(2021年19期)2021-11-07
今日农业(2021年11期)2021-08-13
中国生殖健康(2020年4期)2020-12-09
实用肿瘤学杂志(2020年4期)2020-12-08
中西医结合肝病杂志(2020年2期)2020-10-27
作文评点报·中考版(2019年4期)2019-02-18