
中等强度带通噪声对C57小鼠内毛细胞形态与功能的影响
2019-04-28 09:08李根薛娜宋雷吴皓
中华耳科学杂志 2019年2期
李根 薛娜 宋雷 吴皓
1上海交通大学医学院附属第九人民医院耳鼻咽喉头颈外科(上海200092)
2上海交通大学医学院耳科学研究所(上海200092)
3上海市耳鼻疾病转化医学重点实验室(上海200092)
噪声引起的听力损失全球范围内高发,相关研究表明全球12%以上人口有听力损失的风险[1]。噪声性听力损伤是感音神经性听力损失的最常见形式之一,由于这种损伤在很大程度上是可预防甚至可提前干预的,所以关于噪声损伤的机制以及治疗的研究也一直是听觉研究领域的热点[2,3]。
在先前的研究中人们主要关注点在于噪声后阈值以及感觉毛细胞的变化。直到近年,一系列研究发现噪声导致的听力损伤最初病变主要在听觉毛细胞与传入神经的突触连接处[4,5],而不是毛细胞的丢失。这种病变多数情况下并不伴有阈值的改变,所以起病隐匿,称为噪声性隐性听力损失。当然,虽然早期的损伤并较少出现感觉毛细胞与传入神经的丢失,但是随着时间的推移,毛细胞与传入神经的丢失也会明显加速[5]。而无论是早期的突触损伤还是随后发生的毛细胞与传入神经的丢失,它们所涉及的病理机制都很广泛,且均具有很强的可干预性。这些特征也使得噪声暴露模型广泛应用于治疗药物筛选[6],基因治疗[7,8],乃至耳聋基因的研究中[9]。
目前关于噪声暴露机制以及模型的研究有很多[10,11]。针对小鼠的噪声研究多数是以CBA小鼠为动物模型,而对于基因建模的常用小鼠品系C57小鼠的带通噪声暴露却很少有较为系统的研究。所以本研究拟探讨经8-16kHz,98dB SPL带通噪声暴露2小时后,C57小鼠听力改变以及毛细胞形态与功能的变化,从而建立C57小鼠噪声暴露引起轻度永久阈移的模型。此模型可以为其他基因型噪声暴露的研究提供参考,同时也可以为噪声暴露机制以及各种耳聋机制的研究以及耳聋药物的筛选提供理论依据。
1 材料与方法
1.1 实验动物:取40只SPF级C57雄性小鼠,年龄为6-8周。排除中耳及内耳疾病后进行ABR听阈检测,筛选ABR听阈正常的小鼠作为实验对象,动物饲养以及动物实验方案均严格按照动物伦理委员会实验动物规范进行。
1.2 噪声暴露:将实验动物放在金属网笼中,使用扬声器给与小鼠自由声场噪声暴露。噪声频率为8-16kHz,强度为98dB SPL,暴露时间为2小时。
1.3 听性脑干反应测试(Auditory brainstem response,ABR):测听实验在隔声屏蔽室内进行。测听前使用水合氯醛(480mg/kg)腹腔注射麻醉动物,待小鼠麻醉状态稳定后对其进行听力测试。测听设备为TDT RZ6系统,测听软件为Biosig,实验采用纯音(Pure tone)作为刺激声,声音强度设定为为0-90 dB SPL。每个频率从90dB SPL开始测试,按5 dB SPL递减直至检测不出重复的ABR波形。
1.4 免疫荧光染色:将小鼠麻醉后进行全身PBS心脏灌注,灌注后断头并取出小鼠耳蜗。将耳蜗标本置于4%多聚甲醛溶液中固定过夜。固定结束后将耳蜗脱钙一周,一周后将耳蜗置于PBS中解剖。取出基底膜后用PBS清洗3次再加入封闭液(按照10%驴血清血清,1%Triton X-100,89%PBS进行配比)。室温下封闭1h后加入抗Myo7a与抗CtBP2(按1:200稀释),一抗4℃过夜。一抗孵育完成后用PBS清洗3次,每次10分钟,清洗结束后加入二抗(按1:500稀释),并在37℃温箱孵育1小时。二抗孵育结束后使用PBS清洗3次。清洗结束后进行基底膜铺片,加入封片剂封片。
1.5 激光共聚焦显微镜成像:将制备的样本置于激光共聚焦显微镜(ZEISS LSM 880)下,用40×与60×水镜拍摄,所选择的激发光波长分别为488 nm和594nm。在40X镜下拍摄基底膜毛细胞,60X水镜下层扫拍摄内毛细胞带状突触,扫描层距为0.1μm。拍摄后用分析软件(ZEISS ZEN)分析数据。
1.6 内毛细胞膜片钳记录:取待测小鼠,将小鼠过量水合氯醛麻醉后颈椎脱臼。快速取出小鼠耳蜗后将耳蜗置于培养皿中,并于细胞外液中解剖耳蜗。取出基底膜后将其固定在Dish中在60X水镜下对内毛细胞进行全细胞模式记录。放大器为axon 200B(Axon公司),记录软件为jclamp,电极电阻为4-6MΩ。我们以-80 mV到+70 mV的ramp电压对内毛细胞钙电流进行记录。得到数据后将不同电压下的钙电流减去其对应电压的漏电流即可算得I-V曲线,I-V曲线中电流峰值即为ICa。将I-V曲线以玻尔兹曼方程拟合可得到钙通道激活曲线,从而可计算出钙通道半激活电压Vh以及斜率因子K。同时,通过给予不同时间的电压刺激,记录不同刺激时间前后内毛细胞膜电容的改变(ΔCM)以及刺激过程中的电流变化。刺激时间包括10,50,100,200ms,去极化电压为0mV。计算刺激前后膜电容的数值并相减可得到ΔCM,对刺激过程中钙电流数值进行面积积分可得Qca。
1.7 统计学处理:数据记录与统计学分析采用Excel软件,作图采用Sigma Plot 12.5软件,各组数据以(x±SE)表示,噪声暴露前与第十五天ABR I波波幅与潜伏期的统计采用two-way ANOVA统计方法,其他数据统计均采用配对样本t检验。其中P<0.05为有统计学差异。
2 结果
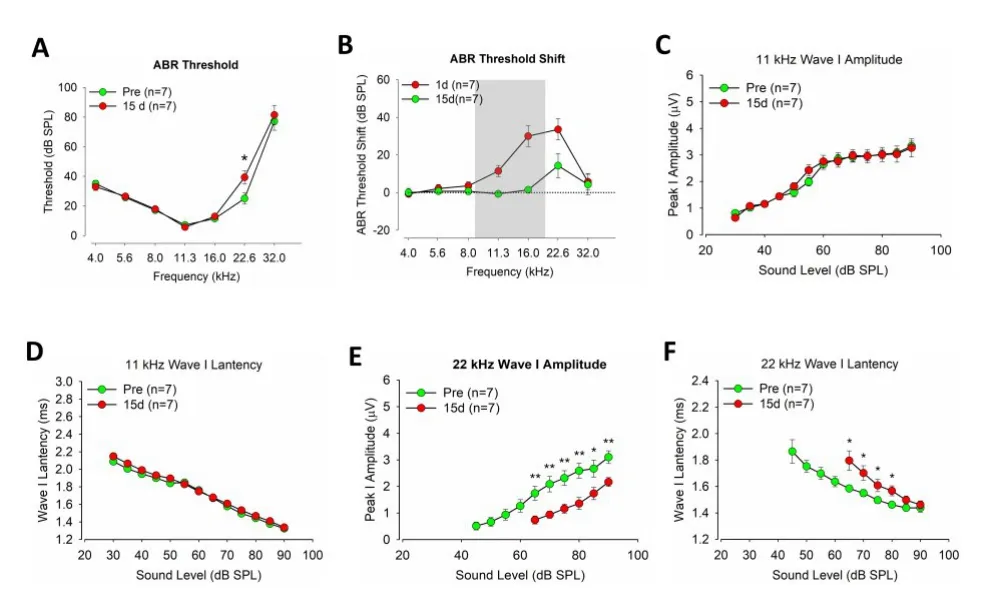
图1 噪声暴露前后C57小鼠听力的变化Fig.1 ABR test of C57 mice before and after noise exposure
2.1 噪声暴露后C57小鼠ABR的改变
噪声后第一天C57小鼠ABR阈移较明显,在第十五天时中低频阈值完全恢复(Fig.1,D),在22kHz区域,实验组ABR阈值显著高于对照组(图1,A),t=2.4136,P=0.0327。相比于噪声暴露前,噪声暴露后C57小鼠在11kHz的ABR I波波幅与潜伏期没有显著改变(图2,C,D),但是在22kHz,65-90dB声强下,实验组小鼠ABR I波波幅的显著低于对照组(图2,E),F=61.6741,P=0.0007。同时潜伏期相比于对照组显著延迟(图2,F),F=7.8658,P=0.0205。
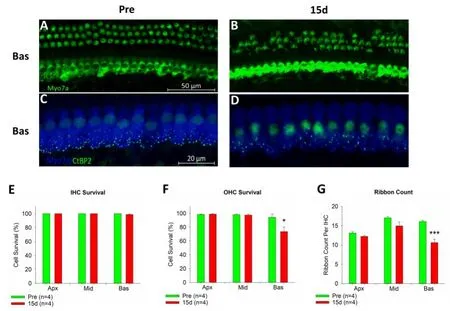
图2 内外毛细胞计数与内毛细胞带状突触计数Fig.2 Immunofluorescence staining of hair cells
2.2 噪声暴露后C57小鼠毛细胞存活率与内毛细胞带状突触数量的改变
毛细胞计数结果显示,实验组与对照组内毛细胞计数无统计学差异(图2,E)。在顶圈与中圈外毛细胞计数在两组中无统计学差异。但是在底圈,实验组外毛细胞存活率约为73%,显著低于对照组(图 2,A,B,F),t=2.3559,P=0.0402。与对照组相比,实验组小鼠内毛细胞带状突触数量在中圈部分下降,在底圈约有34%的丢失,显著低于对照组(Fig.2,C,D,G),t=5.9261,P=0.0004。
2.3 噪声暴露后C57小鼠内毛细胞钙电流与突触囊泡释放能力的改变
我们使用-80 mV到70 mV的RAMP电压刺激内毛细胞并记录钙电流(图3,A)。膜片钳记录结果显示实验组与对照组的钙电流幅值(ICa),钙通道半激活电压(Vh),以及钙通道半激活时对应的斜率因子(K)无显著性差异。(图3,B-D)。
内毛细胞去极化可触发突触通过胞吐作用释放神经递质。胞吐过程中囊泡与细胞膜的融合增加了细胞膜面积,从而使得细胞膜电容增加。而刺激前后膜电容的增加量(ΔCM)反映了递质的释放量[12]。我们给予内毛细胞0mv去极化刺激,改变去极化的时间(包括10mv,50mv,100mv,200mv),并记录去极化过程中流入细胞的钙电荷(QCa)以及ΔCM(图3,E)。
记录结果显示在10ms,50ms,100ms的去极化刺激下,两组小鼠内毛细胞的ΔCM与QCa无显著性差异,但是在200ms的去极化刺激下,实验组小鼠内毛细胞的ΔCM显著小于对照组(图3,F),t=2.1154,P=0.0428。而QCa无显著性差异(图3,G)。结果表明内毛细胞突触囊泡持续释放的能力下降。
3 讨论
噪声是导致听力损失常见的环境因素,同时噪声性聋也是感音神经性听力损失最常见的形式之一。噪声性聋发生的机制及其复杂,涉及纤毛损伤,谷氨酸兴奋性毒性,缺血再灌注损伤,细胞核肿胀,线粒体肿胀,胞质空泡化,钙超载,自由基的形成以及其下游炎症,免疫,凋亡通路的激活等等[13,14]。自由基衰老学说认为自由基的产生是促使机体衰老的重要因素[15]。结合噪声后加速并持续丢失的毛细胞,突触以及螺旋神经节[4,5],我们可以认为噪声暴露加速了耳蜗的衰老与退化。
因此噪声暴露的小鼠模型不仅可以用于噪声性听力损失,感音神经性听力损失的机制研究,还可以用于耳蜗氧化应激,凋亡自噬,炎症免疫乃至毛细胞再生等方面的研究。噪声性聋部分模拟了老年性聋,而且噪声在生活中无处不在,所以噪声暴露也常常用于药物筛选以及致聋基因的研究中[2,3,16]。
在这里我们描述了98dB SPL,8-16kHz带通噪声2小时暴露后C57小鼠听力以及耳蜗病理的改变,建立了噪声暴露引起C57小鼠轻度永久性阈移的模型,不仅为之后的研究提供了基础,也为其他噪声暴露模型的建立提供了参考。
有相关研究表明98dB SPL噪声暴露下CBA发生暂时性阈移,在本研究中我们发现98dB SPL噪声暴露后第十五天后C57小鼠发生了轻度永久阈移。产生阈移的原因可能是噪声对纤毛的损伤导致[17]。此时,在22kHz我们观察到波幅的下降以及潜伏期的延迟,表示内毛细胞突触结构以及其对应的传入神经受到了损伤[18]。一部分原因可能是谷氨酸的大量释放导致了螺旋神经节的兴奋性毒性损伤[4],如螺旋神经节的肿大,回缩,AMPA受体的减少等。另一部分原因可能是由于内毛细胞突触结构与功能的损伤,如Ribbon数量的下降及其位置的改变等[4,5]。相关研究证明噪声暴露后的毛细胞中的ROS,RNS以及钙离子水平显著升高[19,20],谷氨酸的兴奋性神经毒性同样会导致螺旋神经节细胞钙离子的超载[21]。而无论是钙离子还是ROS或RNS,当它们过度增加时都可以作为信号分子诱导下游炎症以及凋亡通路的激活[22,23]。这些机制部分解释了噪声后毛细胞,螺旋神经节的损伤与丢失以及ABR I波波幅降低,潜伏期延迟的现象。
耳蜗可以感受声音,编码声音信号,并将编码的声音信号传入听觉中枢。声音信号在内毛细胞与I型螺旋神经节突触连接处进行编码[24]。声音信号的编码需要神经递质的参与,而在内毛细胞中神经递质以突触小泡的形式锚定在带状突触Ribbon结构的周围[25]。当内毛细胞去极化时钙离子通道开放引发钙内流,流入的钙促发囊泡的释放[26]。突触囊泡不断进行快速释放,补充,回收是内毛细胞需要快速持续编码声音信号的基础[27]。
钙电流记录的数据表明在噪声后第十五天钙离子通道的基本特性并没发生显著性的改变(图3,A-D)。我们给予了不同时间的去极化刺激,通过记录刺激前后细胞膜电容的改变(ΔCM)去比较实验组与对照组内毛细胞突触的快速囊泡释放(<50ms去极化时间)以及持续囊泡释放(>50ms去极化时间)能力[12]。结果表明噪声暴露后在8-11khz区域无明显阈值上升,C57小鼠内毛细胞突触快速囊泡释放能力无明显下降,但是持续囊泡释放能力有所下降。出现这种结果的原因可能是囊泡的补充或者回收收到了一定的损伤[28],而具体的机制有待于更深的研究。
综上所述,98dB SPL的8-16kHz噪声暴露2小时对C57小鼠的听力造成了不可逆的损伤,损伤主要发生在中高频区域,包括内毛细胞突触囊泡释放功能的改变以及高频区域外毛细胞与带状突触的丢失。
猜你喜欢
中国听力语言康复科学杂志(2021年6期)2021-12-21
中华耳科学杂志(2021年1期)2021-02-01
听力学及言语疾病杂志(2020年4期)2020-12-20
复旦学报(医学版)(2020年2期)2020-04-17
发明与创新·中学生(2019年3期)2019-03-20
浙江大学学报(理学版)(2019年1期)2019-02-27
时代英语·初中(2018年2期)2018-05-14
时代英语·初中(2018年2期)2018-05-14
时代英语·初中(2018年2期)2018-05-14
时代英语·初中(2018年2期)2018-05-14
