
系列玉米雄穗突变系的创制
2019-04-24 08:55牛鹏飞田承华姜建芳申晓慧田怀东
山西农业科学 2019年4期
李 南 ,牛鹏飞 ,闫 枫 ,田承华 ,李 珍 ,姜建芳 ,申晓慧 ,栾 鑫 ,田怀东 ,3
(1.山西大学生命科学学院,植物种质与作物资源遗传实验室,山西太原030006;2.山西省农业科学院高粱研究所,山西晋中030600;3.山西景康农业技术推广有限公司,山西寿阳045400)
MNU(N-甲基-N-亚硝基脲)是一种水溶性的化学诱变剂,对鸟嘌呤具有烷化作用。SATOH等[1]使用MNU磷酸盐缓冲液对水稻的穗部进行诱变,成功获得了系列突变体。从水稻的诱变结果可以得知,MNU是一种高效的可以诱导农作物发生基因突变的化学诱变剂。目前,MNU在哺乳动物的肿瘤诱发以及微生物诱变中运用较为广泛,但在农作物的应用中,除水稻外,其他作物生殖细胞的MNU诱变未见报道。近年来,山西大学生命科学学院植物种质与作物资源遗传实验室对水稻、玉米及高粱的玉米花粉生殖细胞进行大规模MNU诱变[2-3]。其中,玉米优异自交系A378是具有Lancaster与Reid血缘属性互补的优异玉米自交系,该自交系穗位低,果穗长,穗部性状好,适合作母本。因此,选取该自交系作为MNU诱变处理的亲本用于繁育后代群体。
玉米雄穗器官的形成是一个动态的发育过程。这个过程主要包括生长锥未伸长期与生长锥伸长期、小穗分化期、小花分化期、性器官分化形成期。玉米雄性生殖器官发育成熟后,开花散粉与雌穗完成授粉过程[4]。玉米雄穗着生于茎秆顶端,由主轴、分枝、小穗和小花组成。主轴上着生有4~11行成对小穗;分枝的数目不定,可能随生理和环境条件的变化而改变,各分枝通常有2行成对小穗[5]。玉米雄穗研究过去主要聚焦于雄性生殖器官及相关组织的发育过程、组织构成、生理特性及其与栽培环境间的相关性[6-7]。近10多年来,国内外开始深入关注玉米雄穗器官的发育、构成和性状表达的遗传机制研究。
玉米雄穗性状突变体的开发和利用是开展玉米雄性生殖器官及相关组织遗传学研究的有效途径。截至目前,国内外主要鉴定出12个玉米花粉不育突变基因(ms系列)[8-13]。这些花粉不育突变影响花粉壁的发育、小孢子壁的发育、小孢子的形成或影响花粉细胞的有丝分裂和减数分裂,而关于玉米雄穗器官及相关组织的其他性状突变体未见报道。全面阐明玉米雄穗发育和性状表达的遗传机制,有必要创制并解析更加多样的雄穗突变材料。
本研究通过玉米成熟花粉生殖细胞MNU处理所得的后代群体雄穗性状的筛查分析,创制出3种类型的玉米雄穗突变系,旨在为进一步在玉米雄性生殖器官突变体的初步鉴定与遗传分析等方面提供理论依据,对阐明玉米雄性不育生殖器官的基因鉴定及其功能解析具有重要意义。
1 材料和方法
1.1 试验材料
将山西大学生命科学学院植物种质与作物资源遗传实验室前期研究中MNU处理优异玉米自交系(A378)成熟花粉生殖细胞所得的543粒M1种子种植于山西大学特色生物资源研究基地,进行M1~M2后代群体的繁育(图1)。
1.2 试验方法
2016年种植M1种子群体、培育M1成株群体,通过M1成株群体的自交获得M2籽粒群体,按每份播种30粒的标准于2017年种植M2籽粒群体,进一步获得M2成株群体(图1)。在M1~M2籽粒与植株群体的繁育过程中,籽粒群体中发生的无胚、畸形胚以及幼苗群体中发生的白化苗、黄化苗、畸形苗与弱化苗等现象,导致群体数目的逐代衰减。通过543粒M1种子群体的繁育,分别得到358株M1成株与341个M2成株群体。
1.3 测定项目及方法
1.3.1 M1与M2雄穗性状的筛查 通过与野生型自交系(A378)雄穗性状相比,主要调查了所得成株群体植株雄穗主轴与分枝的形态、分枝数目、小穗与小花形态以及花粉成熟期散粉状态等雄穗的外观性状特征。将显著变化的雄穗外观性状在M1与M2成株群体中的可遗传性以及在M2成株群体中的表型分离,作为突变性状的判断依据。
1.3.2 玉米花粉活力染色及显微观察 参考崔桂梅等[14]的方法,使用含0.1 mol/L磷酸盐、0.5%TTC的染色液(pH值7.2),对形态特征发生显著变化的M1成株与M2成株的离体花粉进行染色。使用CH20BIMF200显微镜对染色后的花粉进行观察,并分析其活力和育性。
2 结果与分析
2.1 玉米雄穗外观性状突变系的获得
在358株M1成株群体中,仅仅出现3株雌穗顶生的候补突变系,其他类型的雄穗性状变化并未表现。由于M1雌穗顶生候补突变系无散粉能力,所以,将野生型自交系的花粉授于该候补突变系的正常雌穗,获得杂交的M2种子;将其余M1成株自交,获得M2种子。在341份M2成株群体中,观察到1份雄穗雌化候补突变体性状的再现,并筛查到6份雄性不育的候补突变系和2份雄穗分枝畸形的候补突变系(表1)。在雄穗外观上,M2雄性不育候补突变系表现出雄穗主轴发育不良、无分枝、无花粉的特征,M2雄穗雌化候补突变系呈现雄穗主轴退化、雌穗顶生、分枝上部分小穗呈子房状雌化的特征,M2雄穗分枝畸形候补突变系具有正常发育的主轴,显示分枝短小、簇生的特征(图2)。

表1 M2群体中筛查到的雄穗突变系的外观特征

2.2 玉米雄穗候补突变系花粉的活力分析与显微观察
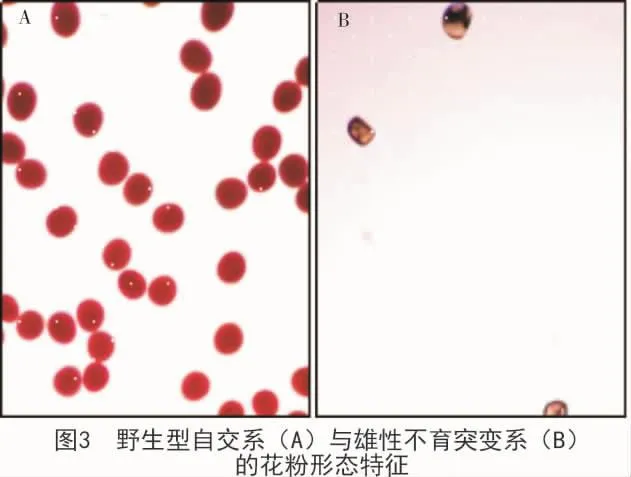
进一步对M2候补突变系的离体花粉进行TTC活力染色分析与显微观察,证实了所创制的雄性不育候补突变系丧失形成花粉的能力(图3)。雄穗雌化候补突变系的分枝小穗虽然包含少量的弱化花粉颗粒,但无散粉与受精能力;雄穗分枝畸形候补突变系的花粉量明显减少,有一定的授粉与受精能力。
2.3 玉米雄穗突变性状的遗传样式解析
对M2成株群体中各雄穗突变性状的表型分离进行了分析。再现的雄穗雌化突变体性状在其与野生型自交系杂交的M2群体中表现出显性遗传的表型分离规律,所有6份雄性不育突变体的性状在其自交的M2群体中均表现出隐性遗传的表型分离规律,2份雄穗分枝畸形突变体的性状在其自交的M2群体中也表现出隐性遗传的表型分离规律(表2)。
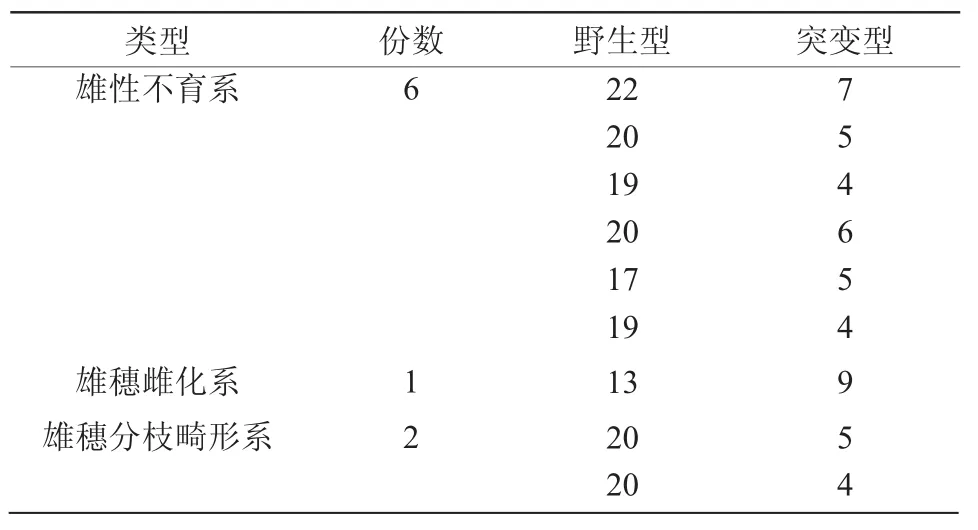
表2 M2群体中雄穗突变系的表型分离
3 结论与讨论
国内外已报道40多个玉米隐性核不育基因突变、部分雄性不育突变的初步定位与功能解析及12个雄性不育突变基因(ms系列)的克隆进展[8-13]。其中,ms1,ms2,ms7突变导致花粉壁发育异常,ms10和ms13突变引起小孢子壁的加厚与降解,ms6,ms8,ms22和 ms23突变影响小孢子的正常形成,ms5突变导致有丝分裂期的花粉细胞中单核或双核的缺失,ms14突变抑制了花粉细胞的减数分裂,ms17突变引起纺锤体形成的异常[15-21]。本研究所创制的6份雄性不育突变系具有的主轴发育不良、无分枝、无花粉性状变化起因于隐性突变,可能与花粉的形成与发育过程有关。期待这些突变体的利用可进一步促进关于花粉形成的发育生物学与遗传性研究。
关于其他类型的雄穗性状突变体,PHILLIPS等[22]与丁帅涛等[23]分别报道了小穗退化玉米突变体与光温敏无雄穗系玉米突变体,但关于其他类型雄穗性状的突变基因未见报道。本研究创制出的雄穗雌化突变系具有主轴退化、雌穗顶生、部分分枝小穗雌化的性状特征,而雄穗分枝畸形突变系具有分枝短小、簇生的性状特征。这些突变系的雄穗性状变化可能起因于和小穗、小花、性器官组织细胞分化有关的基因突变。这些突变体有望用于关于雄穗组织分化与形态构建的遗传学研究。
玉米雄穗器官组织的形成和发育与玉米产量有着密切的关系。本研究所创制的系列雄穗突变体除了可用于雄穗生殖器官与组织的发育生物学与遗传性研究之外,还有望适用于关于玉米产量的遗传育种学研究。
猜你喜欢
果树实用技术与信息(2022年6期)2022-11-21
作物学报(2022年3期)2022-01-22
语文周报·教研版(2021年28期)2021-08-19
作物研究(2020年5期)2020-12-08
农民致富之友(2019年31期)2019-11-23
陕西农业科学(2019年4期)2019-05-13
北京农学院学报(2019年1期)2019-02-22
河南农业科学(2017年4期)2017-04-12
植物保护(2017年3期)2017-01-21
中国蔬菜(2016年8期)2017-01-15
