
外源茉莉酸甲酯对盐胁迫下玉米根系吸水的影响
2019-04-13 01:32严加坤汪亚妮
广东农业科学 2019年1期
严加坤,严 荣,汪亚妮
(榆林学院生命科学学院陕西省陕北矿区生态修复重点实验室,陕西 榆林 719000)
【研究意义】植物生长过程当中或多或少都要受到来自环境胁迫的影响,例如干旱、热、冷及盐等[1-2]。据不完全统计,全球超过7% 的可利用的土地受到盐碱的影响而不能作为正常的耕地使用[3-4]。土壤高含盐量严重影响着植物的生长。因此,研究提高作物抗盐能力的方法具有重要意义。【前人研究进展】在盐胁迫下,植物根系首先感受到高盐带来的渗透胁迫,此时植物通过生理调节例如降低蒸腾作用以减少水分散失,从而维持一定的水分状态;在此过程中,植物根系的吸水作用、茎秆的输水作用起到重要作用[4-6]。一般情况下,植物根系吸水主要通过共质体途径、质外体途径及跨膜运输实现的,而一旦植物受到渗透胁迫将会降低质外体运输;因此,盐胁迫下,水分的跨膜运输对于植株整株的水分平衡起到重要作用[7],而这种扩膜运输有很大程度上受到水通道蛋白活性剂数量的影响[8]。报道显示,盐胁迫下,外源施加硅处理能有效的降低植物根系过氧化氢积累从而接触对水通道蛋白功能活性的抑制进而提高高粱根系的吸水能力[4],说明通过调控水通道活性来改善根系吸水状况从而提高植物对盐胁迫的抗性具有可行性。茉莉酸是一种被广泛作为应对生物胁迫的调节剂是使用的植物激素,并且科研工作者围绕植物生物胁迫做了大量工作[9-10]。最近研究发现,外源施用茉莉酸能有效地提高植物的非生物胁迫抗性[11-13],说明茉莉酸在植物应答非生物胁迫方面具有重要作用。盐胁迫下,外源施用茉莉酸类似物茉莉酸甲酯能够通过提高过氧化氢酶、超氧化物歧化酶及抗坏血酸过氧化物酶活性来减少丙二醛和过氧化氢的积累从而降低氧化损伤近而缓解盐胁迫给小麦幼苗带来的生长抑制[14]。此外,茉莉酸增加盐胁迫下植物抗氧化系统活性也被广泛报道[11-12,15];外源茉莉酸提高菜豆(Phaseolus vulgaris)、拟南芥(Arabidopsis thaliana)及番茄(Solanumlycopersicum)的根系吸水能力也见诸报道[16]。【本研究切入点】短期盐胁迫下,水通道蛋白调控的根系吸水能力的增强被认为是植物应答盐胁迫的重要手段[4,17]。那么短期盐胁迫下,水通道蛋白否参与里茉莉酸介导的植物盐抗性的提升?【拟解决的关键问题】本研究以广适玉米品种郑单958为材料,试图通过外源施加茉莉酸甲酯探索水通道蛋白在玉米适应盐胁迫初期的作用,以期理清水通道蛋白在玉米抗盐胁迫初期的作用,为后期盐碱地玉米种质资源创制提供理论依据。
1 材料与方法
1.1 试验材料及苗期管理
试验于2018年6—9月在陕西省榆林市榆林学院陕西省陕北矿区生态修复重点实验室气候室进行。供试材料为郑单958,种子经1%次氯酸钠溶液消毒15 min,用蒸馏水清洗5次,置于30℃培养箱中催芽3 d。将大小一致的幼苗转移至1/4 Hoaglands营养液中,水培营养液通过加氧机24 h通气供氧。设置光照/黑暗为14 h/10 h,温度为25℃,湿度为55%,光照强度为30 000 lx。待玉米第三片叶片完全展开,50%外源施加1 µmol/L茉莉酸甲酯处理3 d,之后给与100 mmol/L氯化钠处理,另外50%继续Hoaglands营养液培养。试验设Hoaglands营养液(CK)、Hoaglands营养液+茉莉酸甲酯(CK+MeJA)、氯化钠胁迫(S)和盐+茉莉酸甲酯(S+MeJA)4个处理。
1.2 测定项目及方法
盐胁迫5 d后测定玉米生物量积累[17]。玉米采样后按照地上部分和地下根系部分进行分样,装入信封经105℃高温杀青30 min后60℃烘干至恒重,采用分析天平称重。试验设置15个重复。盐胁迫2 h 后测定叶片光合参数、相对含水量[18]、叶片水势[19]、根系水力学导度[19-20]。为验证水通道蛋白作用,进行氯化汞抑制试验[21]。
叶片光合参数选用Li-6400测定。经过2 h胁迫,选用完全展开的第三片叶片测定净光合速率、气孔导度及蒸腾速率。光合参数设置为:光量子500 µmol/m2s,流速为500 µmol/s,待读数稳定后记录。
相对含水量:经过盐处理2 h后,取完全展开的第三片叶片,然后迅速用天平称其鲜重(FW)。称量完毕后,将叶片浸泡在蒸馏水中 6 h,然后将其取出并用吸水纸将表面擦干,测定其总重(TW)。然后将叶片置于 60℃烘箱中烘干并称取其干重(DW)。计算叶片相对含水量。
叶片水势采用3005型压力室进行测定(Soil Moisture Equipment Crop., Santa Barbara,USA),测定方法按照仪器说明进行。
根系水力学导度:经过盐处理2 h后,将玉米幼苗从根基部剪断,留中胚轴4 cm。然后将整株根系用硅胶塞密封后慢慢的对钢瓶加压,每次增加压力 0.1 MPa,直至压力增至 0.5 MPa 为止。每个压力梯度下等达到出流稳态后(大约1 min)用1.5 mL聚乙烯离心管放入脱脂棉吸取汁液,吸水时间统一为 60 s,然后在万分之一天平上准确迅速称量所吸水的重量。完成上述操作后,将根从压力室中取出,应用数字化扫描仪将根系图像存入计算机,然后采用CI-400型根系图像分析系统(Image Analysis Software, CID, Inc.Vancouver,WA)求出根系表面积。每个压力下根系吸水能力用单位时间内单位根表面积流入水量表示,可根据以下公式计算:
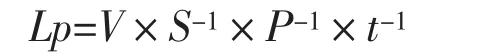
式中,Lp为根系水流导度(m/s·MPa),V为t(s)时间内通过测试根的水流总体积(m3),S为测试根段的表面积(m2),P为达到出流稳态时的平衡压(即外界所加压力,MPa)。最后用所加压力过程中单位面积下水流通量与压力差关系曲线的斜率表示。
氯化汞试验:采用称重法测定整株蒸腾速率。将玉米幼苗在50 µmol/L HgCl2中处理5 min,用蒸馏水清洗,迅速测定蒸腾速率。
试验结果选用最小显著性分析(LSD)用SPSS19.0 处理,采用Sigmaplot12.5 作图。
2 结果与分析
2.1 外源茉莉酸甲酯对盐胁迫下玉米生物量累积的影响
处理5 d后,盐胁迫显著降低了玉米生物量积累(表1);而茉莉酸甲酯处理有效缓解了盐胁迫所造成的玉米生长抑制,外源施加茉莉酸甲酯分别提高盐胁迫下根系生物量41.32%,地上部生物量40.69%。结果再次验证了外源施用茉莉酸能有效缓解盐胁迫对植物造成的生长抑制。

表1 外源茉莉酸对盐胁迫下玉米生物量积累的影响Table 1 Effect of exogenous JA on the biomass accumulation of maize under salt stress
2.2 外源茉莉酸甲酯对盐胁迫下玉米叶片生理参数的影响
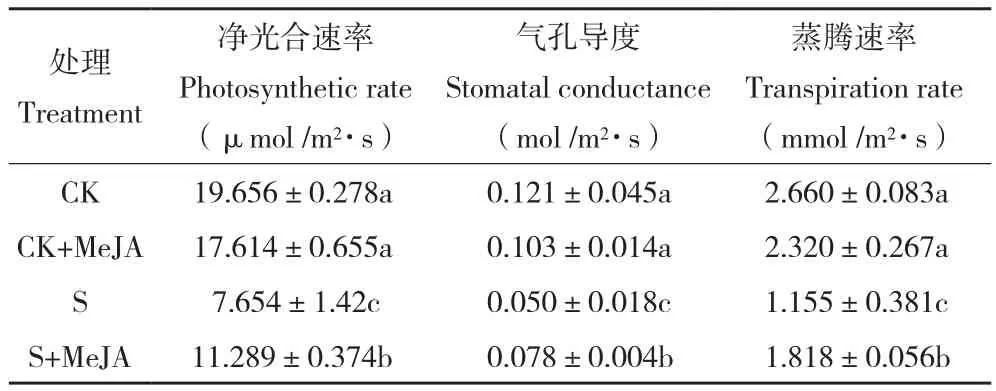
表2 外源茉莉酸对盐胁迫下玉米净光合速率、气孔导度及蒸腾速率的影响Table 2 Effect of exogenous JA on the net photosynthetic rate, stomatal conductance and transpiration rate of maize under salt stress
正常水分条件下,茉莉酸甲酯处理并没有显著影响玉米叶片净光合速率(表2),而盐胁迫条件下,与胁迫处理相比茉莉酸甲酯处理显著提高了玉米叶片净光合速率。说明茉莉酸处理有助于盐胁迫下植物维持相对较高的净光合速率(提高47.48%)。与光合结果类似,盐胁迫下茉莉酸甲酯处理显著提高了玉米叶片的气孔导度及蒸腾速率。通过对叶片相对含水量的统计发现,茉莉酸处理在提高玉米叶片蒸腾速率的同时并没有降低玉米叶片相对含水量,与其结果类似,茉莉酸处理显著提高了盐胁迫下玉米叶片相对含水量(表3)。叶水势结果与叶片相对含水量结果类似,说明盐胁迫下茉莉酸减轻玉米生长的抑制可能与良好的叶片水分状态有关,而高的蒸腾和叶片相对含水量可能与根系吸水能力的提升有关。
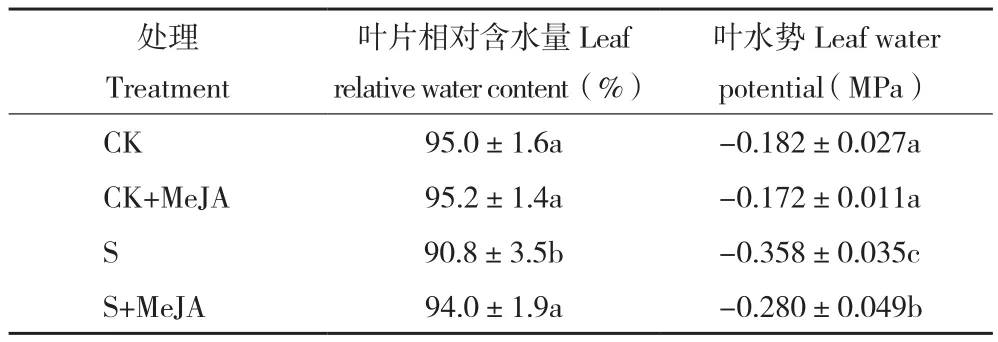
表3 外源茉莉酸对盐胁迫下玉米叶片相对含水量和叶水势的影响Table 3 Effect of exogenous JA on the leaf relative water content and water potential of maize under salt stress
2.3 外源茉莉酸甲酯对盐胁迫下玉米根系水力学导度的影响
为理清根系吸水在维持叶片高含水量中的作用,本试验测定了玉米根系水力学导度。结果(图1)显示,正常条件下,茉莉酸甲酯并没有对玉米根系水力学导度产生显著影响;而在胁迫条件下,茉莉酸甲酯处理显著提高了玉米根系水力学导度,提高41.60%。为验证水通道蛋白在这一过程中的作用,进行了氯化汞试验。结果(图2)显示,相同处理条件下(盐胁迫),氯化汞处理时其叶片蒸腾速率并没有显著差异,说明茉莉酸处理带来的高的玉米根系水力学导度的确与水通道蛋白活性有关。

图1 外源茉莉酸对盐胁迫下玉米根系水力学导度的影响Fig. 1 Effect of exogenous JA on the root hydraulic conductivity(Lp) of maize under salt stress

图2 氯化汞及茉莉酸对盐胁迫下玉米蒸腾速率的影响Fig. 2 Effect of mercuric chlorid and JA on the transpiration rate of maize under salt stress
3 讨论
土壤盐碱化严重限制的植物的生长发育,进而导致粮食减产[22-23];然而研究显示内陆地区正常的地下水灌溉农田正在加速土地盐碱化[24]。因此,植物抗盐碱研究迫在眉睫。本研究结果表明,外源施用茉莉酸甲酯能有效缓解盐胁迫造成的生长抑制,这与前人研究结果一致[25-27]。
3.1 茉莉酸缓解玉米盐胁迫
环境胁迫(盐、干旱及氧化胁迫等)严重抑制植物的叶片气体交换从而抑制植物的净光合速率。本试验中外源施加茉莉酸甲酯显著提高了盐胁迫下玉米的净光合速率,相似的研究在硅提高高粱[21]、茉莉酸提高番茄[11]的抗盐研究中也有发现,这也再次证明茉莉酸能有效的提高玉米的抗盐性。而经过5 d胁迫处理的生物量积累数据也再次证明这一点。
3.2 茉莉酸改善盐胁迫下玉米的水分运输能力
环境因素胁迫严重抑制植物根系水分吸收[17,19,29],而植物整株水分平衡对植物维持正常的生命活动具有重要意义[5,28]。本试验中外源施用茉莉酸甲酯能够有效的提高玉米叶片相对含水量及叶水势,说明茉莉酸能有效的缓解盐胁迫初期给植物带来的叶片水分匮缺。通常盐胁迫对植物的负面效应分为两阶段:首先,胁迫初期高的盐浓度产生渗透胁迫,促使植物根系吸水能力下降从而降低植物叶片含水量进而影响植物光合等生理过程;接下来过多的摄取阳离子,从而产生钠毒害。植物在经历盐胁迫初期主要以渗透胁迫为主,因此茉莉酸提高玉米叶片的相对含水量及叶水势对于植物应对短期的盐胁迫具有重要作用。
环境胁迫下,由于蒸腾拉力的下降,根系水分的跨膜运输对于植物根系吸水起到重要作用[8]。而水通道蛋白是水分跨膜运输的重要工具,在渗透胁迫下对于植物水分吸收及运输起到至关重要的作用[7]。我们发现盐胁迫显著降低了植物的根系水力学导度,这种降低主要是由蒸腾拉力的下降导致的。而外源施用茉莉酸能够缓解盐胁迫带来的玉米根系水力学导度的降低,在这种缓解作用又与水通道蛋白活性紧密相关(氯化汞抑制实验结果),说明水通道蛋白参与了外源茉莉酸介导的玉米根系吸水能力的提升。
4 结论
综上所述,我们认为同硅提高高粱抗盐能力机制类似,盐胁迫下茉莉酸能够通过调控玉米根系水通道蛋白活性来提高玉米根系水力学导度从而提高玉米根系吸水能力进而维持茉莉酸处理下玉米高的叶面蒸腾速率及高的叶片相对含水量。而这种良好的整株水分平衡状态又有利于叶片高的净光合速率的维持从而导致茉莉酸处理下高的玉米生物量积累。
猜你喜欢
农业工程学报(2022年3期)2022-04-16
中国油脂(2022年1期)2022-02-12
中国畜禽种业(2021年9期)2021-09-22
西南农业学报(2021年7期)2021-08-23
节水灌溉(2021年5期)2021-05-26
农业工程学报(2021年3期)2021-04-15
节水灌溉(2020年9期)2020-09-16
节水灌溉(2020年7期)2020-07-15
中国农业科技导报(2020年12期)2020-03-15
润滑与密封(2019年3期)2019-03-22
