
海水胁迫对冰菜细胞损伤和有机小分子渗透调节物质积累影响
2019-04-13 01:32段瑞军吴朝波黄圣卓梅文莉戴好富
广东农业科学 2019年1期
段瑞军,吴朝波 ,王 军,黄圣卓,梅文莉,戴好富
(1.中国热带农业科学院热带生物技术研究所,海南 海口 571101;2.海南好授粉农业科技服务有限公司,海南 乐东 570228;3.海南正和职业培训学校,海南 海口 571101)
【研究意义】植物对盐渍化生境的适应实现方式主要通过渗透调节、肉质化、离子区域化、盐离子选择性吸收及气孔泌盐等作用,其中渗透调节为主要方式,植物细胞中合成有机小分子溶质为渗透调节的方式之一。研究发现,盐胁迫下,植物积累脯氨酸、甜菜碱、可溶性糖和多胺等有机小分子渗透调节物质,行使其调节、保护、清除活性氧、促进生长等功能,在植物耐盐性上具有重要作用[1]。【前人研究进展】冰菜(Mesembryanthemum crystallinumL.)叶片肉质化、娇嫩、较脆,富含叶酸、β-胡罗素、蛋白质和多种微量元素,营养丰富[2],可在含盐度与海水等同(含盐度3.5%)的条件下正常生长,是理想的盐碱地推广蔬菜,为理想的岛礁蔬菜种植候选品种。目前关于冰菜的研究主要集中在栽培种植方面[3-4],其生理生化方面仅有一些零星报道[5-6]。【本研究切入点】笔者所在课题组自引种冰菜在海南文昌盐碱地试种以来,通过不同浓度海水处理所在课题表明,冰菜在0~100%浓度下均能完成生活史,但海水浓度高于60%冰菜生长受到抑制,并对其抗氧化酶系统进行了研究[7]。【拟解决的关键问题】为进一步摸清冰菜耐盐生理机理,优化了试验处理,测定不同浓度海水下膜质过氧化产物(MDA)和细胞膜透性,获取不同浓度海水下细胞膜的损伤情况,同时测定脯氨酸、可溶性糖、甜菜碱等有机小分子渗透调节物质的含量变化,以期揭示冰菜有机小分子渗透调节物质与海水胁迫的关系。
1 材料与方法
1.1 试验材料
冰菜种子收集于中国热带农业科学院热带生物技术研究所文昌试验基地,海水取自海口市白沙门。
1.2 试验方法
试验在中国热带农业科学院生物技术研究所进行,2018年3月播种育苗,冰菜苗长出4片真叶时进行移栽,在前期研究基础上设置5个浓度(20%、30%、40%、50%、60%)海水。移栽后一周进行海水处理,每周浇2次。分3个时期(即出现一级分枝、二级分枝、三级分枝)采集相同部位叶片进行MDA、细胞膜透性指标和脯氨酸、甜菜碱、可溶性糖等有机小分子渗透调节物质含量测定。
MDA含量测定采用硫代巴比妥酸显色法[8],细胞膜透性测定采用电导率法[9],脯氨酸含量测定采用茚三酮显色比色法[10];甜菜碱含量测定采用雷氏盐沉淀法[11],可溶性糖含量测定采用蒽酮比色法稍作改进[12]。
数据用Excel 2003整理,并绘制图表;用SPSS13.0进行方差分析和多重比较,进行统计显著性检验。
2 结果与分析
2.1 不同浓度海水对冰菜MDA含量和细胞膜透性的影响
逆胁迫下质膜是首先受到攻击的目标之一,膜质受到损伤,膜脂过氧化产物MDA含量增加、膜透性增强。根据不同浓度海水下、不同生长时期的冰菜MDA含量(图1)可知,冰菜在20%浓度海水处理下,MDA含量最低。一级分枝和二级分枝MDA含量增长规律一致,在20%~40%浓度海水处理下细胞中MDA含量积累无显著增加,当海水浓度≥40%时,细胞中MDA积累量显著增加。浓度海水达到60%时,一级分枝、二级分枝中MDA含量分别是20%浓度的1.56、1.70倍。三级分枝中MDA含量在海水浓度为50%时达到最大,是20%浓度的2.56倍,之后呈下降趋势。在相同海水浓度下,随着冰菜生长,MDA积累量不断增加,且浓度越高,积累量越多,20%、30%、40%、50%浓度海水胁迫下,三级分枝MDA含量分别比一级分枝增加0.57、0.79、0.88、0.91倍。

图1 海水胁迫对冰菜MDA含量的影响Fig. 1 Effects of seawater stress on MDA content of M. crystallinum L.
冰菜细胞膜透性结果(图2)表明,相同海水浓度下,冰菜细胞膜受损伤程度为三级分枝>二级分枝>一级分枝,在20%、30%、40%、50%、60%浓度海水胁迫下细胞膜透性三级分枝分别是一级分枝的2.09、1.72、1.33、1.65、1.42倍;同一分枝在不同海水浓度胁迫下,细胞膜透性随海水浓度增加而增强,冰菜细胞一级分枝、二级分枝、三级分枝膜透性在60%浓度海水胁迫下分别是20%浓度海水下的4.38、3.01、2.98倍。

图2 海水胁迫对冰菜细胞膜透性的影响Fig. 2 Effects of seawater stress on cell membrane permeability of M.crystallinum L.
2.2 不同浓度海水对冰菜脯氨酸积累量的影响
脯氨酸含量在一定程度上反映了植物的抗逆性,本试验将脯氨酸含量作为抗逆育种的重要生理指标。另外,由于脯氨酸亲水性极强,能稳定原生质胶体及组织内的代谢过程,能降低凝固点,可防止细胞脱水。由图3可知,不同浓度海水处理对各级分枝的影响总体趋势相似,脯氨酸含量在三级分枝中最高,其次为二级分枝,一级分枝中最少。当海水浓度≤30%时,随着海水浓度升高,脯氨酸含量增加,二级分枝较显著。当海水浓度≥30%时,随着浓度海水增加,脯氨酸含量呈现降低趋势,二级分枝和三级分枝下降显著。不同分枝中,30%浓度海水处理下脯氨酸含量升高显著,其中二级分枝和三级分枝脯氨酸含量已超过25 µg/g。一级分枝脯氨酸含量在各浓度海水处理下变化没有二级分枝和三级分枝明显。

图3 海水胁迫对冰菜脯氨酸含量的影响Fig. 3 Effects of seawater stress on proline content of M. crystallinum L.
2.3 不同浓度海水对冰菜可溶性糖积累量的影响
可溶性糖在冰菜生长发育中具有重要作用,不仅为各时期提供能量和代谢中间产物,且具有信号功能,也是植物生长发育和基因表达的重要调节因子。图4所示,不同浓度海水处理在各级分枝呈现相同的影响趋势,均随着浓度海水升高,可溶性糖含量增加,且含量呈现三级阶梯式显著升高;各级分枝高浓度海水胁迫下,可溶性糖含量显著高于低浓度处理,其中60%浓度海水处理下可溶性糖含量为20%处理的近2倍。不同浓度海水对一级分枝影响不如二级分支和三级分枝明显,因为一级分枝可溶性糖含量普遍较低,约为其他分枝的50%。
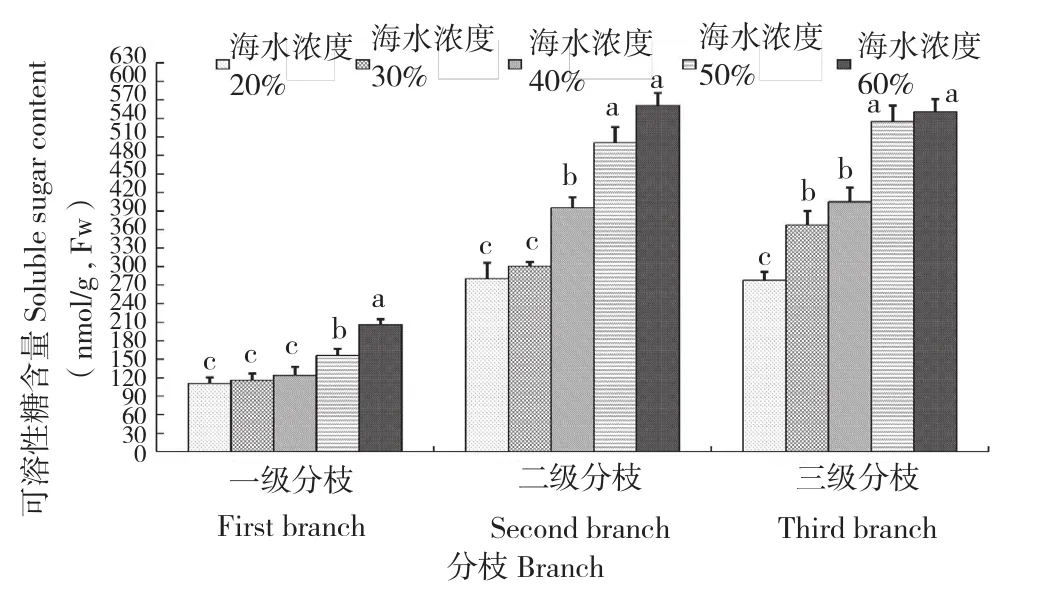
图4 海水胁迫对冰菜可溶性糖含量的影响Fig. 4 Effects of seawater stress on content of soluble sugar of M. crystallinum L.
2.4 不同浓度海水对冰菜甜菜碱积累量的影响

图5 海水胁迫对冰菜甜菜碱含量的影响Fig. 5 Effects of seawater stress on content of betaine of M. crystallinum L.
甜菜碱的积累对维持细胞内外的渗透平衡与细胞的正常生理活动有重要意义。本试验结果表明:不同海水浓度下,当海水浓度为20%~60%时,冰菜一级、二级分枝甜菜碱含量随海水浓度的增加而增加,当海水浓度为20%时,一级、二级分枝中甜菜碱含量分别为1 800、2 324 µg/g,当海水浓度达60%,甜菜碱含量明显增加,分别是20%海南浓度下的2.43、3.02倍。三级分枝中甜菜碱呈先增大后减小的趋势,在40%海水浓度下达到最大值(7 455 µg/g),为20%、60%下的1.82、2.32倍。相同海水浓度下,在20%~60%浓度海水胁迫下,一级、二级分枝在相同浓度下甜菜碱含量均呈增加趋势,60%浓度海水胁迫下冰菜一级、二级分枝甜菜碱积累量分别是20%浓度处理的2.27、2.29、2.05、1.53倍。60%浓度海水胁迫下,甜菜碱积累量二级分枝>一级分枝>三级分枝。
2.5 膜损伤指标、有机小分子、海水间的相关性分析
分析不同海水浓度下各指标相关性(表1)可知,海水浓度与膜损伤指标细胞膜透性极显著正相关,相关系数为0.972,与膜质过氧化产物MDA呈显著正相关,相关系数为0.957;海水浓度与甜菜碱含量呈正相关,其中与可溶性糖含量呈极显著正相关,与脯氨酸积累量呈负相关。电导率、MDA与可溶性糖含量呈极显著正相关,相关系数分别为0.966、0.963。可溶性糖积累与脯氨酸积累呈显著负相关,相关系数为0.907。综合分析可知,有机小分子渗透调节物质可溶性糖的积累能对膜损伤做出最快速的反应。

表1 冰菜在海水胁迫下各试验指标之间的相关性分析Table1 Correlation analysis of various test indexes of M. crystallinum L. under seawater stress
2.6 海水胁迫与各有机小分子渗透调节物质间在各生长时期的相关性分析
由图6A可知,冰菜一级分枝脯氨酸积累量与海水浓度呈低度负相关,相关系数为0.073;冰菜二级分枝脯氨酸积累量与海水浓度呈中度负相关,相关系数为0.651;冰菜三级分枝脯氨酸积累量与海水浓度呈高度负相关,相关系数为0.894。即冰菜生长期越长,脯氨酸与海水胁迫的相关性越高。由图6B可知,冰菜一级分枝可溶性糖积累量与海水浓度呈显著正相关,相关系数为0.850;冰菜二级、三级分枝可溶性糖积累量与海水浓度呈极显著正相关,相关系数分别为0.966、0.955。即冰菜各生长期时期、各海水浓度下,可溶性糖均能对海水胁迫做出响应。由图6C可知,冰菜一级分枝甜菜碱积累量与海水浓度呈极显著正相关,相关系数为0.950;二级分枝甜菜碱积累量与海水浓度呈显著正相关,相关系数分别为0.906;三级分枝甜菜碱积累量与海水浓度呈低度负相关,相关系数分别为0.015。即冰菜生长前期,甜菜碱能及时为对海水胁迫做出响应,随着冰菜生长,甜菜碱对海水胁迫响应逐渐减弱。

图6 海水浓度与脯氨酸、可溶性糖和甜菜碱的相关性分析Fig. 6 Correlation analysis of seawater concentration with proline, soluble sugar and betaine
3 讨论
3.1 海水胁迫对冰菜细胞膜的损伤
逆境胁迫下植物细胞内自由基代谢平衡遭到破坏,导致膜脂透性增大、膜脂过氧化物积累,使细胞膜系统严重受损。研究发现,盐浓度与细胞膜透性呈正比[13]。电解质外渗率即电导率可以反映细胞膜透性,代表细胞膜系统的完整性,细胞膜透性越大表明膜系统受损程度越大。对白刺的耐盐性研究发现,3种白刺的电导率值均随盐胁迫增加而增大[14];丙二醛是膜脂过氧化产物,能与细胞内各种成分发生反应,进而引起膜损伤[15],其含量高低可间接表现植物受逆境胁迫的强弱。孙海菁等[16]研究发现,在中低盐度胁迫下细胞膜透性增加不明显,而在高盐度胁迫下迅速增加,与本研究结果一致。本研究表明,细胞膜透性、MDA均随海水浓度增加而增加,在海水浓度为20%~60%范围内,一级、二级分枝均无明显增加,海水浓度为50%时,各级分枝MDA含量、细胞膜透性均较20%下显著增加。同一海水浓度下,生长期越长,细胞膜透性越大,膜质过氧化产物MDA积累量越多,均呈现三级分枝>二级分枝>一级分枝的基本变化规律,表明,冰菜在20%~60%海水浓度范围内,具有极强的抗盐性。
3.2 海水胁迫有机小分子渗透调节物质的响应
盐胁迫下植物细胞内大量失水,导致渗透胁迫,为维持植物正常的生理代谢,通过渗透调节降低细胞水势来吸收水分。脯氨酸、甜菜碱、可溶性糖等有机小分子渗透调节物质,有助于植物调节细胞渗透压,消除或减轻盐胁迫损伤,维持植物生长。银杏幼树在盐胁迫下,可溶性糖含量随盐浓度的升高和盐胁迫时间延长呈先升后降趋势,脯氨酸含量在盐胁迫后期显著增加[17],枸杞幼苗在盐胁迫下脯氨酸和可溶性糖含量在不同处理时期均呈现上升趋势[18],花椰菜苗期盐胁迫下脯氨酸含量与盐浓度极显著正相关[19],对蓟幼苗NaCl胁迫下游离脯氨酸含量先增后减小[20];不同牧草中可溶性糖含量变化不同,披碱草、草地早熟禾和星星草随着盐浓度的提高,可溶性糖含量呈现增高的趋势,而老芒麦呈现先升后降的趋势[21]。盐胁迫后胡杨根叶中甜菜碱浓度显著提高,在处理后15 d达到最高值,特别是叶片中甜菜碱的浓度提高243倍,达到1 899.8µmol/L,根中甜菜碱含量也增加了9倍[22]。可见,脯氨酸、甜菜碱、可溶性糖在盐胁迫下对植物抗盐性具有重要作用,且各种植物中变化不一,但总的说来,盐胁迫下,上述渗透调节物质均会不断积累。本研究中,冰菜可溶性糖含量随着海水浓度增加,积累量不断增加,相同生长阶段,不同分枝中在60%浓度海水处理下可溶性糖含量为20%处理下的近2倍,同一海水浓度处理,可溶性糖含量三级分枝>二级分枝>一级分枝。说明盐胁迫时间越长、胁迫浓度越大,可溶性糖含量越高。脯氨酸含量在各级分枝中均表现出先增大后减小的趋势,表明脯氨酸能对低浓度海水胁迫做出响应,随着海水浓度增加,响应程度降低。甜菜碱在冰菜生长前期,即一级分枝、二级分枝表现为随海水浓度增加,积累量增多,冰菜生长至三级分枝时,甜菜碱积累量呈现先增加后减小的趋势;在20%~50%浓度海水下,同一浓度海水处理冰菜随着生长期增长,其体内甜菜含量增加,表明甜菜碱在20%~50%海水处理下,能及时对盐胁迫做出响应。
3.3 海水胁迫与膜损伤指标和有机小分子渗透调节物质的相关性
通过对MDA积累量、细胞膜透性、脯氨酸、甜菜碱、可溶性糖与海水相关分析表明,膜损伤指标MDA含量、膜透性与海水胁迫具有较高的正相关性,表明海水浓度增加对冰菜损伤作用增强。海水浓度与甜菜碱含量呈正相关,其中与可溶性糖含量呈极显著正相关,与脯氨酸积累量呈负相关;可溶性糖含量与细胞膜透性、MDA呈极显著正相关。冰菜3个生长期(一级、二级、三级分枝)有机小分子渗透调节物质与海水胁迫的线性回归分析结果表明,海水胁迫与冰菜生长一级分枝呈显著正相关,与二级、三级分枝呈极显著正相关。海水胁迫与甜菜碱的积累量相关性在一级分枝为极显著正相关,二级分枝为显著正相关,三级分枝为低度负相关,表明随着冰菜的生长相关性逐渐降低,即冰菜生长前期,甜菜碱在渗透调剂中起着重要作用。海水胁迫与脯氨酸含量呈负相关,且处理时间越长负相关度越高,在三级分枝时呈现极显著负相关,表明脯氨酸在盐胁迫下随着冰菜生长其调节作用在减弱。
4 结论
随着海水浓度增强,处理时间延长,冰菜细胞膜内膜质过氧化产物MDA迅速积累,细胞膜透性增强,而冰菜在20%~60%海水处理下依然能正常生长,维持生物量,可溶性糖、脯氨酸、甜菜碱均能做出不同程度的响应,调节冰菜细胞渗透压,维持冰菜正常生理活动,促进冰菜生长。在参与渗透调节的有机小分子物质中,与海水胁迫的相关性表现为可溶性糖>甜菜碱>脯氨酸。
猜你喜欢
养猪(2022年1期)2022-11-21
中老年保健(2022年3期)2022-11-21
中国畜禽种业(2022年8期)2022-09-17
中国现代中药(2022年1期)2022-03-03
安徽化工(2021年3期)2021-05-29
心肺血管病杂志(2020年5期)2021-01-14
棉花学报(2020年3期)2020-08-08
科学与财富(2019年15期)2019-10-21
分析化学(2018年11期)2018-01-16
发明与创新·中学生(2016年5期)2016-05-14
