
金观音与其亲本差异基因表达的遗传分析
2019-04-09 02:23姚雪倩郑玉成王鹏杰郑知临陈桂信叶乃兴
福建农林大学学报(自然科学版) 2019年2期
姚雪倩, 郑玉成, 王鹏杰, 林 浥, 郑知临, 陈桂信, 叶乃兴
(1.福建农林大学园艺学院,福建 福州 350002;2.茶学福建省高校重点实验室,福建 福州 350002;3.闽侯县农业局,福建 闽侯 350100)
茶树是多年生异花授粉植物,遗传背景复杂,杂合程度很高,杂种优势的利用难度较大[1].铁观音和黄旦是乌龙茶的主栽品种,福建省农业科学院茶叶研究所以铁观音为母本,黄旦为父本,从其杂交后代中选育出金观音、黄观音、金玫瑰、金牡丹等早生优质新品种[2].金观音,又名茗科1号,是福建省推广面积最大的乌龙茶新品种[3].金观音的生育期、芽叶色泽、成茶品质等农艺性状与亲本存在差异,在产量、香气品质和抗逆性等方面均超过父母本,表现出明显的杂种优势[4].目前关于金观音与其亲本间的差异研究只停留在亲子代间的形态特征、生理生化特性、制茶品质等方面[4],用遗传学理论来阐释茶树亲子代间差异基因表达的机理还鲜见报道.
本试验从金观音与其亲本间的差减文库中筛选出与生长发育、逆境胁迫、次生代谢有关的差异基因,采用荧光定量PCR技术检测其表达量的变化,分析差异基因的表达模式;并检测具有超高亲表达和低于双亲表达模式的基因在其他两个F1黄观音和金玫瑰上表达量的差异,旨在为揭示差异基因在杂种和亲本上的遗传规律提供参考.
1 材料与方法
1.1 材料
2016年3月中旬至4月中旬于福建农林大学南区教学茶场,以中小开面叶为标准采摘铁观音(♀)、黄旦(♂)及3个F1品种金观音、黄观音和金玫瑰的一芽二叶.每个品种分别称取3份以上,每份0.2 g,用锡箔纸包裹,做好标记,放入液氮速冻后,转入冰箱(-80 ℃)保存备用.
1.2 方法
1.2.1 金观音与其亲本差异基因的筛选 建库和筛选参照前人[5-7]的方法进行.以金观音及其亲本的春茶嫩梢为供试材料,采用双链特异性核酸酶(DSN)介导的抑制差减杂交技术,构建金观音—铁观音及金观音—黄旦两个正向差减cDNA文库,采用反向印迹杂交技术与荧光定量PCR技术相结合的方法筛选这两个正向差减cDNA文库,获得差异基因的阳性克隆.将阳性克隆的测序结果进行生物信息学分析,预测其生物学功能,并将这些差异基因进行归类.
1.2.2 金观音与其亲本差异基因实时荧光定量PCR分析 以金观音、铁观音和黄旦的一芽二叶为材料,以天根RNAprep Pure Plant Kit(Polysaccharides & Polyphenolics-rich)植物多糖多酚试剂盒提取的总RNA为模板,按照全式金的EasyScript®One-Step gDNA Removal and cDNA Synthesis SuperMix试剂盒方法合成的cDNA作为荧光定量PCR的模板.根据两个差减文库中筛选得到的参与生长发育、逆境胁迫和次生代谢相关基因的cDNA序列,以茶树GAPDH(KC337052.1)为内参基因[8],利用Primer Premier 5.0软件设计荧光定量PCR引物(部分引物见表1).荧光定量PCR反应体系含5 μL 2×TransStart®Tip Green qPCR SuperMix、上下游引物各0.2 μL、1 μL Template DNA ,用ddH2O补至10 μL;程序为:95 ℃ 3 min;95 ℃ 15 s,60 ℃ 34 s,40个循环.每一个基因模板设置3个生物学重复,试验通过Bio-Rad CFX 3.1荧光定量PCR仪完成.数据结果采用Excel 2003软件和2-△△Ct法[9]进行定量分析.

表1 荧光定量PCR的引物序列Table 1 Primer sequences for quantitative real-time PCR
1.2.3 金观音与其亲本差异基因表达量的比较及筛选 以F1金观音的荧光定量表达量高于母本铁观音或父本黄旦之一较高的表达量称为超高亲表达量,具有杂种优势;以F1金观音的荧光定量表达量低于母本铁观音和父本黄旦的表达量称为低于双亲表达量;以F1金观音的荧光定量表达量介于母本铁观音和父本黄旦的表达量之间的称为中亲表达量;母本铁观音和父本黄旦的荧光定量表达量差别很大,以F1金观音的表达量偏向双亲中表达量较高的一方称为偏高亲表达量,以F1金观音的表达量偏向双亲中表达量较低的一方称为偏低亲表达量.比较参与生长发育、逆境胁迫和次生代谢相关基因荧光定量表达量的差异,分别将这三类差异基因以金观音的表达量为参照进行分组.
1.2.4 差异基因在黄观音和金玫瑰中的实时荧光定量PCR分析 以黄观音和金玫瑰的一芽二叶为材料,总RNA提取,荧光定量PCR模板、引物、反应体系、反应程序和数据结果定量分析与“1.2.2”的方法一致.
2 结果与分析
2.1 金观音与其亲本差异基因的分类
分别从金观音—铁观音和金观音—黄旦差减文库中筛选出90和113条有效的单基因序列,其中,共有基因14个,76个差异基因的性状遗传黄旦,99个差异基因的性状遗传铁观音.在这两个差减文库中均筛选出参与生长发育、逆境胁迫和次生代谢等9类基因(表2).
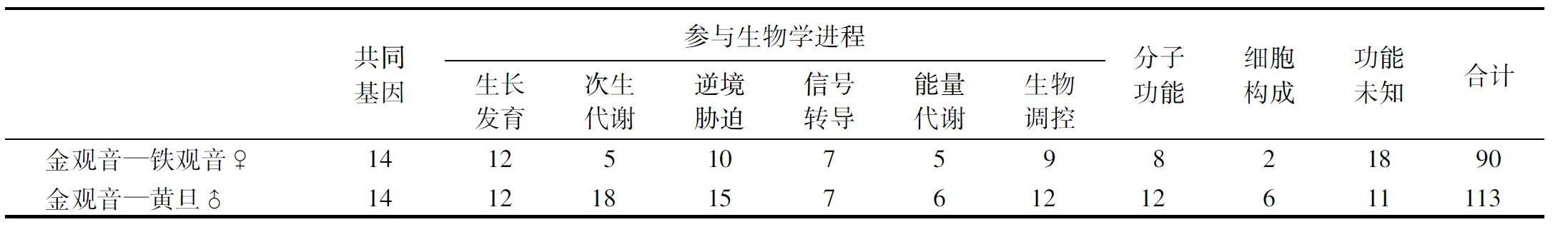
表2 两个差减文库差异基因的个数Table 2 The number of differential genes between 2 suppression subtractive libraries
2.2 金观音与其亲本差异基因的表达量
从金观音—铁观音和金观音—黄旦差减文库中筛选出的基因分别标记为“TGY+序列号”和“HD+序列号”
2.2.1 金观音与其亲本参与生长发育类基因的表达模式 F1金观音与其母本铁观音、父本黄旦参与生长发育类基因的表达模式分类如图1所示.由图1可知,此类基因有超高亲(杂种优势)、低于双亲、中亲、偏高亲和偏低亲5种表达模式.属于超高亲(杂种优势)表达量的基因有15个,分别为TGY5、TGY15、TGY189、TGY203、TGY206、TGY239、TGY253、TGY283、TGY454、TGY468、HD123、HD307、HD318、HD380、HD384;属于低于双亲表达量的基因有5个,分别为TGY145、TGY160、TGY497、HD80、HD292;属于中亲表达量的基因有2个,分别为TGY186、TGY327;属于偏高亲表达量的基因有4个,分别为TGY159、HD152、HD229、HD266;属于偏低亲表达量的基因有4个,分别为TGY93、TGY345、HD73、HD473.

图1 金观音与其亲本参与三类差异基因的表达模式Fig.1 Expression patterns for 3 kinds of differential genes between Jinguanyin and its parents
2.2.2 金观音与其亲本参与逆境胁迫类基因的表达模式 F1金观音与其母本铁观音、父本黄旦参与抗性类基因的表达模式分类见图1.由图1可知,此类基因有超高亲(杂种优势)、低于双亲和中亲3种表达模式.属于超高亲(杂种优势)表达量的基因有10个,分别为TGY23、TGY117、TGY297、TGY494、HD15、HD86、HD122、HD331、HD451、HD467;属于低于双亲表达量的基因有5个,分别为TGY145、TGY300、HD402、HD409、HD472;属于中亲表达量的基因有2个,分别为TGY304、TGY421.
2.2.3 金观音与其亲本参与次生代谢类基因的表达模式 F1金观音与其母本铁观音、父本黄旦参与品质类基因的表达模式分类见图1.由图1可知,此类基因有超高亲(杂种优势)、低于双亲、中亲和偏低亲4种表达模式.属于超高亲(杂种优势)表达量的基因有4个,分别为TGY23、HD277、TGY482、HD27(前两个基因与花青素苷合成相关,后两个基因与香气代谢相关);属于低于双亲表达量的基因有6个,分别为TGY139、TGY147、HD32、HD136、HD342、TGY149(前5个基因与花青素苷合成相关,最后一个基因与香气代谢相关);属于中亲表达量的基因有1个,为HD402(与花青素苷合成相关);属于偏低亲表达量的基因有1个,为HD512(与香气代谢相关).
2.3 差异基因在亲本与其3个F1品种的表达量
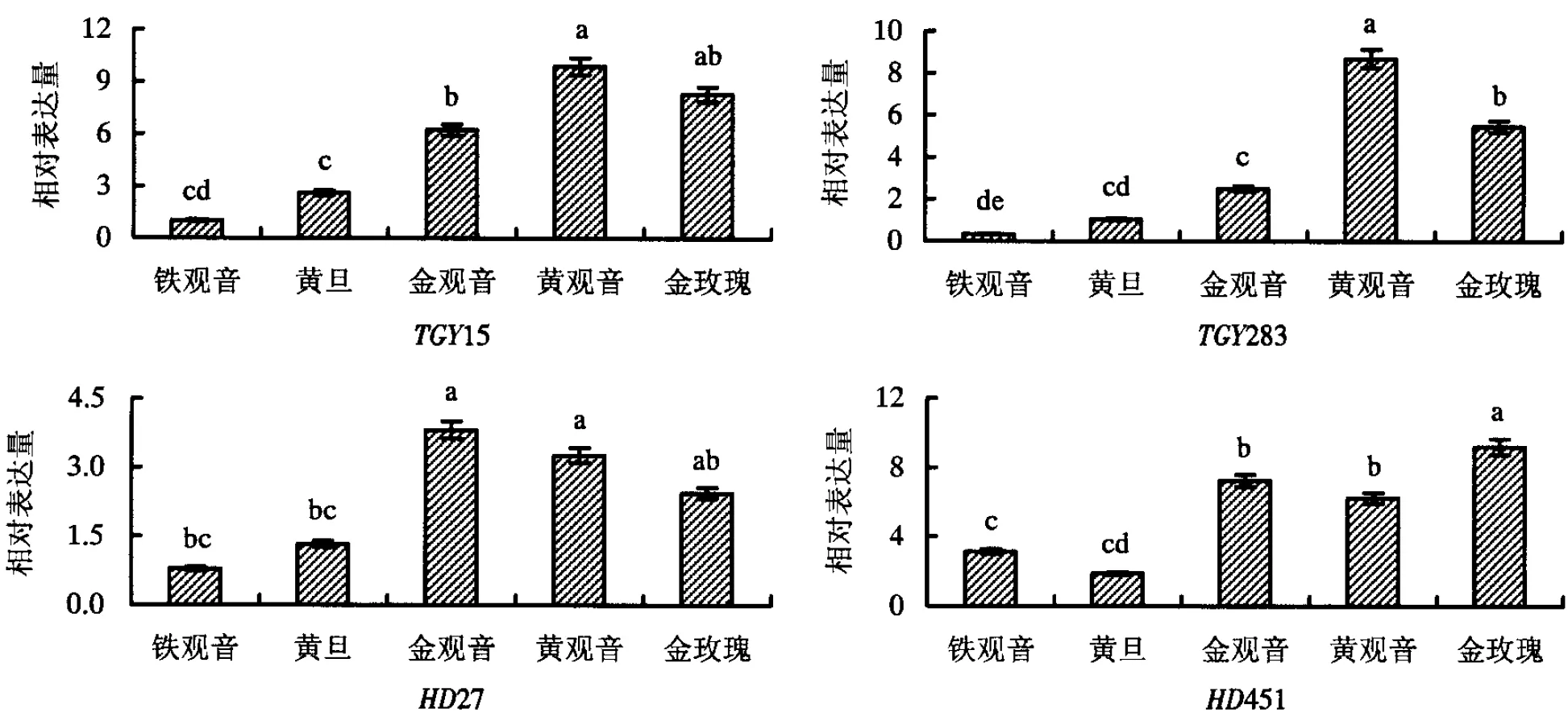
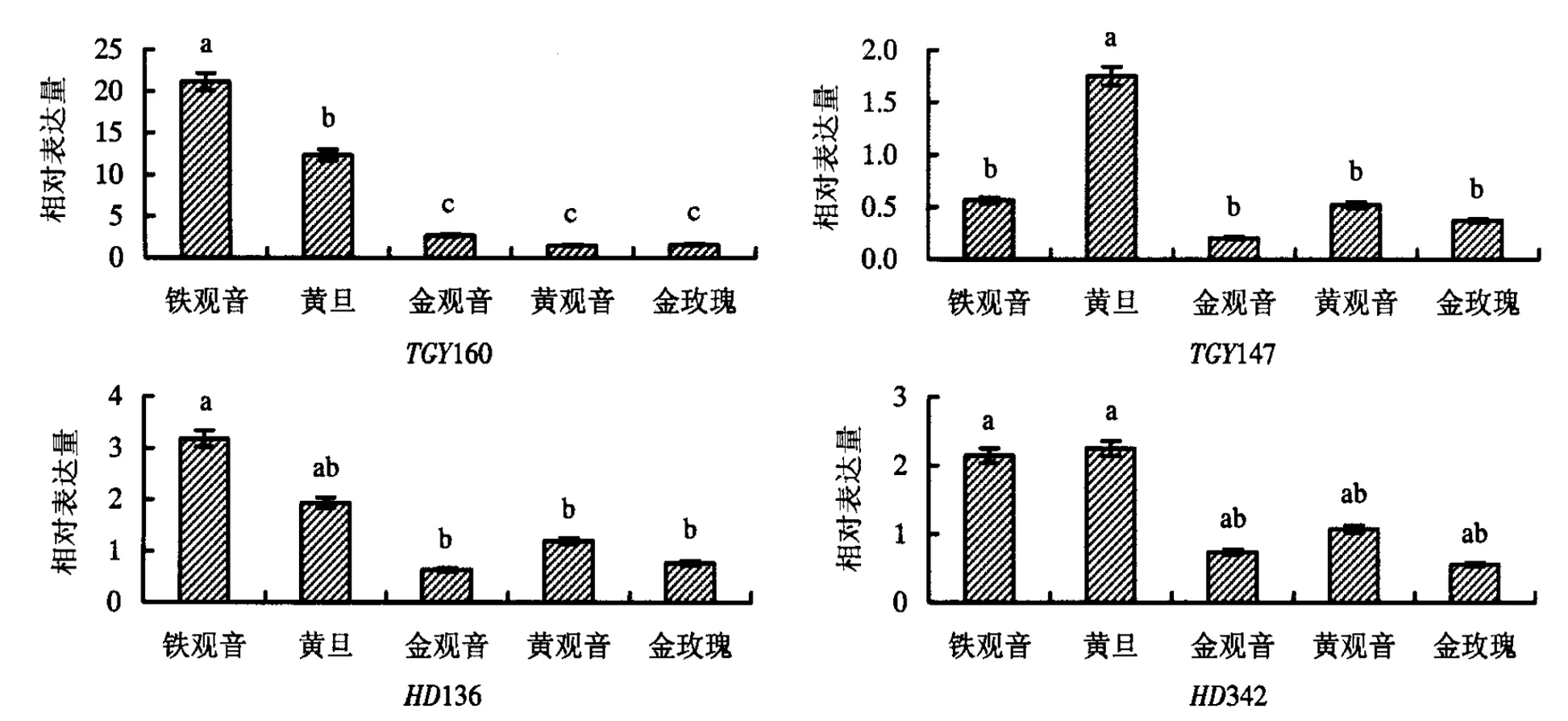
从超高亲和低于双亲两组表达模式中随机筛选12个基因(表1)进行荧光定量PCR分析,根据金观音、黄观音和金玫瑰3个F1品种与其父母本的表达量差异,又可分为3种类型,一是TGY15、TGY283(生长发育类基因)、HD451(逆境胁迫类基因)和HD27(香气代谢类基因),在3个F1品种上的表达量均超过双亲(图2),即存在杂种优势;二是TGY160(生长发育类基因)、TGY147、HD136和HD342(花青素苷合成类基因),在3个F1品种上的表达量均低于双亲(图3),即存在杂种劣势;三是TGY145、HD292(生长发育类基因)、TGY149(香气代谢类基因)和HD277(花青素苷合成类基因),在3个F1品种上的表达量部分超过双亲,部分低于双亲(图4),即杂种优势和杂种劣势共同存在.
图柱上附不同字母者表示差异显著(P<0.05),附相同字母者表示差异不显著(P>0.05).图2 杂种优势基因的表达量Fig.2 The expression of heterosis genes
图柱上附不同字母者表示差异显著(P<0.05),附相同字母者表示差异不显著(P>0.05).图3 杂种劣势基因的表达量Fig.3 The expression of hybrid weakness genes
3 讨论
从金观音—铁观音和金观音—黄旦两个差减文库中筛选出的遗传差异基因,其中一部分基因属于两个差减文库共有的,表明这部分基因由亲本共同遗传给后代的;另一部分基因则是遗传铁观音或黄旦,遗传铁观音的差异基因总数多于遗传黄旦的,表明金观音的遗传性状多数趋向母本,属于偏母本融合型杂种[10].
图柱上附不同字母者表示差异显著(P<0.05),附相同字母者表示差异不显著(P>0.05).图4 兼具杂种优、劣势基因的表达量Fig.4 The expression of genes with features of both heterosis and hybrid weakness
与生长发育、逆境胁迫和次生代谢相关的基因在F1金观音及其母本铁观音、父本黄旦中均有表达,但实生后代发生大幅度的变异,依据表达量上的差异可分为超高亲、低于双亲、中亲、偏高亲和偏低亲5种表达模式.其中,中亲表达所占比例很低,但这也反映出实生后代趋中变异的一个现象,即后代群体的平均水平常有返回某一中数的倾向[11].除中亲表达属于加性表达外,其余4种表达也称作非加性表达[12],这与Li et al[13]提出的关于双亲中不同表达量的基因比相同表达量的基因更易呈现非加性表达模式的观点相近.
茶树芽叶成熟期的早晚、芽叶色泽、抗逆性和香气等属于数量性状.与生长发育、逆境胁迫和次生代谢相关的基因在F1金观音中的表达量有的介于两亲本之间,有的超过双亲,有的则小于双亲的平均值,F1金观音表现出的变异现象符合数量性状的表型特点.郭吉春等[4]研究认为,金观音和黄旦属于早生种,铁观音属于晚生种,但本试验中参与生长发育基因(如TGY160、HD292等)的表达量并不都是金观音与黄旦相近且高于铁观音;铁观音幼嫩芽叶呈紫红色,黄旦和金观音幼嫩芽叶呈黄绿色[14],因此从两亲本与F1两个差减文库中筛选出的直接编码花青素苷合成途径的酶类(如TGY23、HD342等)可能来源于试验取样叶片紫红色与非紫红色的差异.研究表明,茶树幼嫩芽叶的紫化现象与叶片中花青素的积累密切相关[15].但本试验中参与花青素苷合成基因(如TGY147、HD277等)的表达量并不都是金观音与铁观音相近且高于黄旦;金观音的抗性和香气成分(如乙酸乙酯、α-萜品醇、水杨酸甲酯、香叶醇、茉莉内酯等)超过铁观音和黄旦[16],杂种优势强,但本试验中参与抗性和香气代谢基因(如TGY145、TGY149等)的表达量并不都是金观音高于双亲.上述前人的研究结果与本试验参与生长发育、逆境胁迫和次生代谢相关的基因在金观音及其亲本中表达趋势的结果不完全一致,表明F1和亲本中基因表达的差异既有可能受顺式作用元件、反式作用元件或二者共同作用的影响,也有可能与它们的基因表达调控、表观遗传机制、数量性状基因位点有关[17].
本试验筛选出的超高亲基因包括与生长发育、逆境胁迫和次生代谢相关的基因,表明F1金观音具有多方面的优势表现.但金观音杂种优势表现的程度不同,其优势有强有弱,这可能是环境因子与基因相互作用的结果[18].铁观音与黄旦杂交产生的其他F1黄观音和金玫瑰有的均存在杂种优势,可能来源于铁观音与黄旦大量优异等位基因的聚合,符合超显性假说的观点[19];有的并非都存在杂种优势,可能由于杂种一代与亲本之间的组织生长和发育过程呈动态变化,在供试样品取样时难以达到完全统一的生长势标准,因此出现基因表达差异.就生育期、抗逆性、芽叶颜色和香气等性状而言,每一性状受多少对基因的支配很难估计,这就涉及到数量性状与杂种优势的关系,推测生育期、抗性和品质等数量性状是从微效多基因控制过渡到主基因—多基因混合控制形成茶树杂种优势[20].铁观音与黄旦杂种一代表达水平上的多态性决定了研究茶树杂种优势机理的复杂性.3个F1品种基因表达之间的差异与它们不同性状的关系以及某个功能基因具体是哪种表达模式还需进一步探讨.
铁观音和黄旦作为重要的乌龙茶育种材料,通过杂交选育了一批优良的茶树品种,丰富了乌龙茶品种资源,对乌龙茶的发展产生了深远的影响.但铁观音与黄旦杂交后基因的遗传规律复杂,还有待深入研究.因此,对亲子代间的差异基因进行表达分析,了解杂种和亲本的遗传趋势,对杂种的早期选择、杂种优势形成理论的解析具有理论和实践价值.
猜你喜欢
蔬菜(2021年7期)2021-11-27
少年文艺·我爱写作文(2021年5期)2021-07-20
今日农业(2020年18期)2020-12-14
东坡赤壁诗词(2020年5期)2020-11-06
海峡姐妹(2019年11期)2019-12-23
小学生时代·综合版(2016年4期)2016-11-19
化工进展(2015年3期)2015-11-11
种子科技(2014年8期)2014-05-18
中国质量与标准导报(2014年10期)2014-02-28
茶叶通讯(2014年2期)2014-02-27
