
蚜蝇科昆虫比较线粒体基因组分析
2019-03-18 05:27霍科科
陕西理工大学学报(自然科学版) 2019年1期
李 娟, 李 虎,2*, 霍科科
(1.陕西理工大学 生物科学与工程学院, 陕西 汉中 723000;2.陕西省资源生物重点实验室, 陕西 汉中 723000)
蚜蝇,又称访花蚜蝇,是蚜蝇科(Syrphidae)昆虫的总称,全世界已知约有3亚科230属6000余种,广泛分布于世界各地[1]。成虫具有访花习性,幼虫的捕食范围较为广泛。通常,管蚜蝇亚科幼虫寄生在枯木中,食用土壤、池塘和溪流中腐烂的植物和动物残体[2-3];巢穴蚜蝇亚科的幼虫在蚂蚁巢穴里寄居[4];蚜蝇亚科幼虫具有捕食性,捕食蚜虫、蓟马和其他刺吸植物的害虫[5-7]。因此蚜蝇与人类生产关系密切。
蚜蝇科昆虫系统发育进化关系,多采用形态学形状和单个基因或少数几个基因相结合的方法进行研究。传统分类学鉴定需要相当高的技能和分类学专长,对于某些物种之间,尤其是形态相似性较高的物种,在形态鉴定过程中会出现很大的困难[8]。由于不同基因的进化速率和保守性的差异,在利用单个基因和少数几个基因分析不同阶元的系统发育时,结果往往是有差异的。近年来随着生物技术的快速发展,测序成本不断降低,使得线粒体基因组全序列被广泛地应用于种群结构和支序关系的研究。
线粒体基因组全序列被看做是一种很有前途的新工具,可用于深层理解昆虫的系统遗传学[9]。昆虫线粒体基因组全序列具有母系遗传、基因组成稳定、基因排列顺序相对保守以及极少发生重组等特点[10],因而被广泛应用于动物的起源、演化和分类,以及系统进化和亲缘关系的研究[11]。截止2018年9月,通过检索美国国立生物技术信息中心(National Center for Biotechnology Information,NCBI)的GenBank数据库和查阅文献资料,已完成线粒体基因组全序列测序的蚜蝇共有6种:Ocyptamussativus(登录号:KT272862)[12]、Syrphidae sp.(登录号:KM244713)[13]、黑带蚜蝇Episyrphusbalteatus(登录号:KU351241)[14]、大灰优蚜蝇Eupeodescorolla(登录号:KU379658)[14]、大角类蚜蝇Simosyrphusgrandicornis(登录号:DQ866050)[9]和长尾管蚜蝇Eristalistenax[15],其中Syrphidae sp.和大灰优蚜蝇Eupeodescorolla的线粒体基因组序列还不完整。
本文通过对现有的线粒体基因组全序列进行比较分析,总结蚜蝇科昆虫线粒体基因组全序列的结构特征,并构建基于线粒体基因组全序列的系统发育关系,验证并讨论当前的分类系统。
1 线粒体基因组全序列在系统发育中的优势
Tautz等[16]首次提出用DNA序列作为生物分类的主要依据,即DNA分类学。Hebert[17]等进一步提出DNA条形码计划,并指出线粒体基因细胞色素C氧化酶I(COI)可以作为全球生物识别系统的核心。COI基因可以很好地用于分析科与科之间的系统发育关系[18],但对于同种不同群体之间的系统发育关系的分析仍存在一定问题[19]。一个完整的线粒体基因组全序列可以提供更丰富的系统遗传信息,因此可能比单个基因或多个基因在解决系统发育和进化关系上具有更明显的优势[20]。随着GenBank数据库中线粒体基因组全序列的不断补充,大量的研究结果表明,线粒体基因组全序列可为深层次的分析生物系统发育和进化关系提供更充分的分子理论依据。
2 昆虫线粒体基因组全序列的应用概况
线粒体基因组全序列可用于昆虫的系统发育关系重建、物种和行为起源与进化、种群遗传变异分化、近缘种以及种下分类单元的鉴定等,其中应用于鳞翅目、鞘翅目等昆虫的系统进化关系和DNA条形码等研究领域已经取得了许多令人瞩目的成果。张敏等[21]通过对小红珠绢蝶线粒体基因组全序列分析蝶类高级阶元系统发育关系,其研究结果与传统的分类学结果基本一致,支持绢蝶科物种归为绢蝶亚科;绢蝶亚科、锯凤蝶亚科和凤蝶亚科属于凤蝶科,并且绢蝶亚科与锯凤蝶亚科为姊妹群。杨洪等[22]通过对烟蚜线粒体基因组全序列进行系统发育分析,结果证实了蚜亚科和长管蚜亚科的单系性。Timmermans等[23]基于53个线粒体基因组全序列的12个编码蛋白基因的联合数据对鞘翅目亚目间的系统发育关系进行了探讨,结果表明肉食亚目与藻食亚目互为姊妹群,多食亚目圆花蚤科(Scirtidae)在鞘翅目亚目的基部,叩甲系和扁甲系为单系,叩甲系与多食亚目的其他4个系所形成的进化枝形成姊妹群。杨秀帅[24]基于线粒体基因组全序列数据获得的夜蛾总科系统发育关系,支持最新的夜蛾总科六科系统和弄蝶科属于凤蝶总科。
3 蚜蝇科昆虫线粒体基因组全序列的结构与碱基组成
3.1 蚜蝇科线粒体基因组全序列的结构
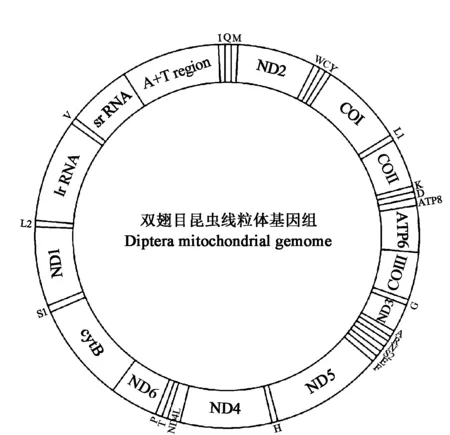
图1 双翅目昆虫线粒体基因组
蚜蝇科线粒体基因组包含37个编码基因,其中包括22个转移RNA基因,13个蛋白编码基因(PCGs)、2个核糖体RNA基因和非编码控制区,与双翅目昆虫的线粒体基因组特点相同(图1)[25];其中有23个基因在J链上编码,14个基因在N链上编码,符合昆虫线粒体基因组结构特点[26]。13个蛋白编码基因,其中9个基因位于J链,4个基因位于N链。由GenBank数据库已知的5种蚜蝇科昆虫线粒体基因组全序列长度以及长尾管蚜蝇线粒体基因组全序列的长度进行比较分析(表1),可得出蚜蝇科昆虫线粒体基因组全序列长度约在11 583~16 175 bp之间,平均长度为15 088 bp。其中未知种Syrphidae sp.的线粒体基因组全序列最短,为11 583 bp,黑带蚜蝇的线粒体基因组全序列最长,为16 175 bp[13-14]。
3.2 碱基组成
对线粒体基因组全序列中碱基A、T、G、C的组成进行比较分析(表1)发现,G的含量均低于C的含量(G%
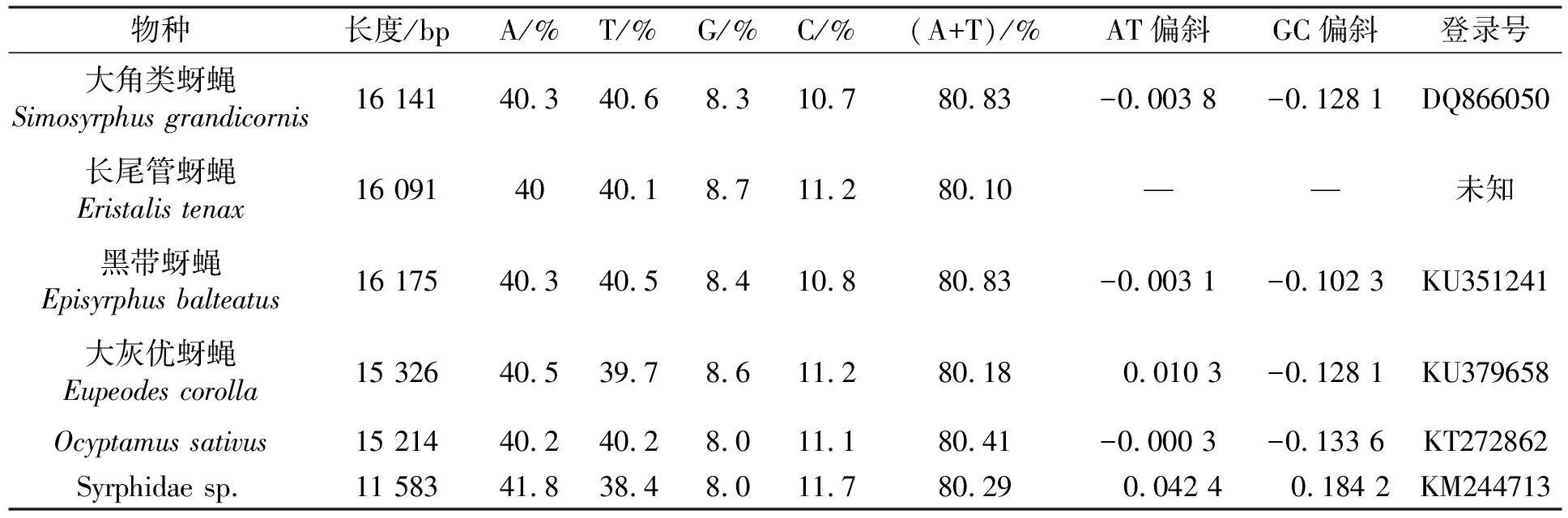
表1 6种蚜蝇线粒体基因组全序列比较
通常,线粒体基因组全序列碱基组成特征可用3个参数表示,即A+T含量、AT偏斜和GC偏斜,AT偏斜与GC偏斜分别使用公式AT-skew=(A-T)/(A+T)、GC-skew=(G-C)/(G+C)来计算[27]。蚜蝇科昆虫线粒体基因组全序列在碱基组成中表现出很强的AT偏向性((A+T)%>(G+C)%),对这6种蚜蝇科昆虫线粒体基因组全序列计算得出,大角类蚜蝇Simosyrphusgrandicornis和黑带蚜蝇Episyrphusbalteatus的线粒体基因组中A+T含量均为80.83%,Ocyptamussativus为80.41%,Syrphidae sp.为80.29%,大灰优蚜蝇Eupeodescorolla为80.18%,长尾管蚜蝇EristalistenaxA+T含量为80.1%[15]。
3.3 基因重排、基因间隔与基因重叠
通过比对发现,很多目昆虫的线粒体基因组基因重排现象非常普遍,例如鳞翅目、半翅目、缨翅目等[28-30]。不同物种基因重排排序方式可能不同,例如在鳞翅目中云南蝠蛾的线粒体基因组排序方式类似于果蝇,而不同于家蚕[31]。目前已测序的蚜蝇科昆虫线粒体基因组全序列中,并未发现基因重排现象。蚜蝇科昆虫线粒体基因组中出现了基因重叠和基因间隔,重叠范围大多在1~7 bp之间,最长的基因重叠区域位于atp8和atp6、nad4和nad4l之间。基因间隔的间隔距离范围较大且数量多,如黑带蚜蝇Episyrphusbalteatus在22个不同的位置上出现基因间隔,间隔范围为1~60 bp,长尾管蚜蝇Eristalistenax在20个不同的位置上出现了基因间隔,间隔范围为1~37 bp,大灰优蚜蝇Eupeodescorolla间隔基因出现在19个不同的位置,间隔范围为2~47 bp等。在大灰优蚜蝇Eupeodescorolla和黑带蚜蝇Episyrphusbalteatus中最长的间隔基因位于trnK和trnD之间,其次是位于trnE和trnF之间[14]。间隔基因的出现可能有利于储存较多的遗传信息,也可能会增加变异概率,有利于生物进化。
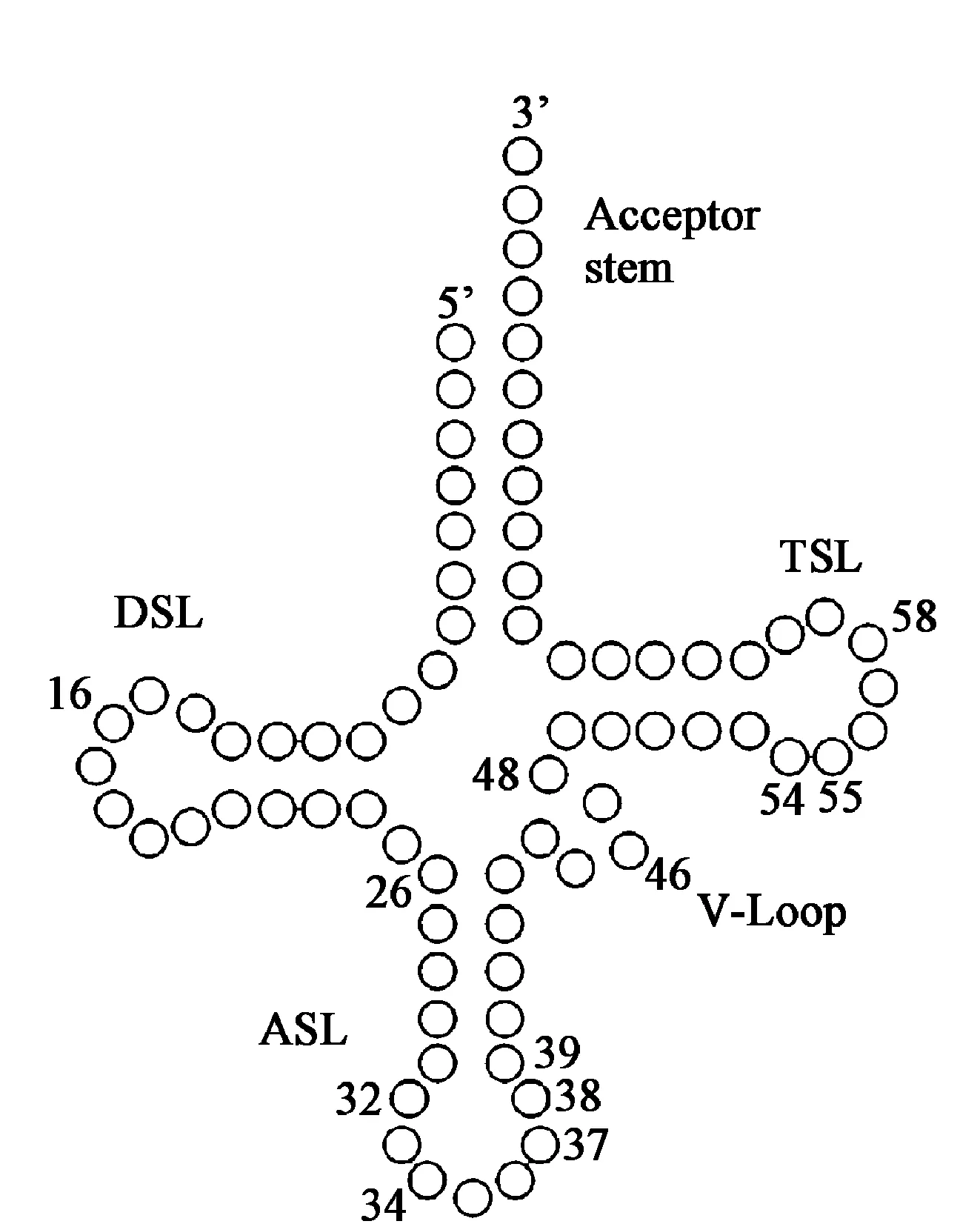
图2 tRNA典型的二级结构
3.4 转移RNA和核糖体RNA基因
黑带蚜蝇Episyrphusbalteatus和大灰优蚜蝇Eupeodescorolla线粒体基因组中,所有tRNA基因的二级结构都可折叠成典型的三叶草结构(图2)[32];所有的tRNA的二氢尿苷(DHU)臂是一个大环,而不是一个保守的茎环结构;trnM基因序列最保守,没有任何核苷酸变化;出现碱基错配现象[14]。rRNA包括rrnl和rrns,其结构特点未见报道。
3.5 密码子使用
大多数线粒体蛋白编码基因的起始密码子都是ATN,与之不同的是,长尾管蚜蝇Eristalistenax[15]的COI和ND1的起始密码子为TTG,COIII使用的起始密码子是TGG。除了大灰优蚜蝇中ND5的终止密码子是一个不完全的TA[14],其余所有的蛋白编码基因使用的终止密码子都为TAA。在黑带蚜蝇Episyrphusbalteatus和大灰优蚜蝇Eupeodescorolla线粒体基因组全序列中,由于Leu、Ile、Phe和Met是最常被编码的氨基酸,因此TTA(Leu)、ATT(Ile)、TTT(Phe)和ATA(Met)是最常见的密码子[14]。4个物种蛋白编码基因的AT偏斜和GC偏斜如表2所示[14],仅有大灰优蚜蝇Eupeodescorollae的AT偏斜为正数,其余均为负数,并且GC偏斜的绝对值都大于AT偏斜。

表2 PCGs核苷酸长度及偏斜值
4 基于线粒体基因组蚜蝇科系统发育关系研究
选取8个物种线粒体基因组全序列构建系统发育树,包括来自蚤蝇科Phoridae、鼓翅蝇科Sepsidae和佛蝇科Fergusoninidae的3个外群,这8个物种均属于双翅目,亲缘关系较近[14],其中蚤蝇科和蚜蝇科属于无缝组,鼓翅蝇科和佛蝇科属于有缝组。检索GenBank数据库已有的线粒体基因组全序列,使用最大似然法(ML)分析系统发育树(图3)。

图3 基于线粒体基因组全序列构建的最大似然法系统发育关系
3个外群中,Fergusoninataylori(佛蝇科Fergusoninidae)最先分出,位于树的根部,接着Megaseliascalaris(蚤蝇科Phoridae)和Nemopodamamaevi(鼓翅蝇科Sepsidae)分出并聚在一起构成姐妹群关系,蚜蝇科Syrphidae与(蚤蝇科Phoridae+鼓翅蝇科Sepsidae)构成姊妹群关系。5种蚜蝇聚类成一大支,构成单系,系统发育关系为(Syrphidae sp.+(Ocyptamussativus+(大灰优蚜蝇Eupeodescorollae+(大角类蚜蝇Simosyrphusgrandicornis+黑带蚜蝇Episyrphusbalteatus)))),其中大角类蚜蝇Simosyrphusgrandicornis和黑带蚜蝇Episyrphusbalteatus构成姊妹群关系。
5 讨 论
截至2018年9月,检索GenBank数据库和查阅相关文献发现,已测出线粒体基因组全序列的蚜蝇科昆虫共有6种,其中基因组完整的4种,不完整的2种。仅占蚜蝇科昆虫总数极小的比例,蚜蝇科昆虫线粒体基因组研究仍有很大的空间。
对这6种蚜蝇科昆虫线粒体基因组进行比较分析,发现蚜蝇科昆虫线粒体基因组包含37个编码基因(其中包括22个转移RNA基因,13个蛋白编码基因(PCGs)、2个核糖体RNA基因)和非编码区,在碱基组成上具有很强的AT偏向性,未出现基因重排现象以及tRNA基因的二级结构为典型的三叶草结构等,比较符合昆虫线粒体基因组的特点[26]。
基因重排现象在很多目的昆虫中都有发生,例如鳞翅目、半翅目、缨翅目等[28-30]。同样,在双翅目中也出现了基因重排,均出现在长角亚目中[26],例如在蚊科中伊蚊的trnA与trnR发生重排,形成trnR-trnA[33],但蚜蝇科目前仍未发现基因重排现象,这可能与双翅目昆虫线粒体基因的保守性有关[25],也可能是目前所测得的蚜蝇科昆虫线粒体基因组的数据过少,还不能充分反映该科物种线粒体基因的排列顺序。
在密码子使用方面,绝大多数线粒体蛋白编码基因的起始密码子都是标准的三联体ATN,个别蛋白编码基因使用特殊的起始密码子如TTG、TGG,研究表明,这种特殊的起始密码子可以减小基因间隔区,避免相邻基因发生重叠,可转录成为mRNA后经过RNA编辑转换成正常的起始密码子,然后翻译成特定的蛋白质[34]。终止密码子为TA和不完整的T,有研究者[35]推测不完整的T会在转录后通过向mRNA的3′端添加A进行多聚腺苷酸化,进而转化为完整的终止密码子而完成转录终止。
本研究结果支持蚜蝇科的单系性,这与Pu等[14]的研究结果相同。Li等[15]利用线粒体基因组全序列中所有的蛋白编码基因对8个物种进行系统发育分析,结果同样支持了蚜蝇科的单系性,并且蚜蝇科与头蝇科Pipunculidae是姊妹群关系。张卓等[36]利用COI~COII序列对4个不同地区的黑带蚜蝇种群进行分析,发现产生了遗传差异并且已经具有明显的遗传分化。张宏杰等[37]对管蚜蝇族Eristalini的7种蚜蝇Cty b基因序列进行分析,表明管蚜蝇族的4个属为单系且条胸蚜蝇属Helophilus和宽盾蚜蝇属Phytomia的亲缘关系相对较远。
蚜蝇科昆虫线粒体基因组全序列已测出的种数较少,在生物系统发育关系等方面的分析主要以单个基因或多个基因序列为主导。在系统发育分析中使用频率较高的线粒体基因有COI基因、COII基因、Cty b基因等。因此,应重视对蚜蝇科昆虫线粒体基因组的研究,以期获得更多的线粒体基因组数据来探讨蚜蝇科系统发育关系。
猜你喜欢
中华实用诊断与治疗杂志(2022年1期)2022-08-31
今日农业(2021年11期)2021-08-13
海洋通报(2021年1期)2021-07-23
生物学通报(2021年4期)2021-03-16
中国生殖健康(2020年4期)2020-12-09
中西医结合肝病杂志(2020年2期)2020-10-27
生物学通报(2020年11期)2020-10-22
发明与创新·中学生(2019年6期)2019-06-26
中成药(2018年7期)2018-08-04
中成药(2018年7期)2018-08-04
