日本血吸虫感染小鼠的髓源抑制细胞对T细胞的免疫抑制①
2019-03-13 05:06高勇强曹劲松肖建华
中国免疫学杂志 2019年3期
高勇强 黎 丽 胡 丽 梁 瑜 曹劲松 肖建华
(特殊病原体防控湖南省重点实验室,南华大学病原生物学研究所,衡阳421001)
血吸虫病(Schistosomiasis)是一种呈世界性分布、严重危害人类健康的人兽共患寄生虫病[1]。在血吸虫感染小鼠模型中,既有以Th1细胞为主的保护性免疫应答,又有以Th2细胞为主的病理性免疫应答,两者之间维持着动态的免疫平衡[2,3]。近年来研究发现,在荷瘤小鼠骨髓、脾脏、血液和肿瘤组织,以及肿瘤患者外周血和肿瘤组织中,广泛存在着一群髓系来源抑制细胞(Myeloid-derived suppressor cells,MDSCs),这些细胞来源于骨髓祖细胞和未成熟髓细胞,形态幼稚,以幼稚粒细胞(MDSC-G)和幼稚单核细胞(MDSC-M)为主,其共同的表面标志为Gr-1+CD11b+,具有强大的免疫抑制和广泛的免疫调节功能,如抑制CD4+T细胞增殖和抗原特异性CD8+T细胞分泌IFN-γ等,与肿瘤免疫逃逸和免疫耐受密切相关[4,5]。在寄生虫感染中,有关 MDSCs 的研究,近几年也有少量报道[6,7]。例如弓形虫、克氏锥虫、硕大利什曼原虫和牛带绦虫感染宿主后,宿主体内 MDSCs 大量增殖,肺部、外周血液和腹腔积液中 MDSCs 细胞数量也明显增多,且呈现抑制宿主的免疫应答的作用。为了研究在日本血吸虫感染小鼠体内诱导产生的 MDSCs 细胞是否具有免疫抑制功能及其作用机制,我们通过构建血吸虫感染小鼠模型来研究MDSCs对血吸虫感染的免疫抑制作用和探讨可能的作用机制,为血吸虫防治探索新的作用靶点。
1 材料与方法
1.1实验材料 60只6~8周,18~22 g,SPF级饲养的健康BALB/c小鼠,购自长沙史莱克景达实验动物有限公司。血吸虫尾蚴阳性钉螺购自岳阳市血吸虫病防治研究所。Percp-Cy 5.5 conjugated Rat Anti-Mouse CD11b antibody、APC conjugated Rat Anti-Mouse Gr1 antibody、PE conjugated Rat Anti-Mouse Gr1 antibody,以上抗体购自美国BD Bioscience公司。Anti-biotin-APC、CD4+T cell isolation kit(mouse)、CD11b+cell isolation kit(mouse)、CD8+T cell isolation kit(mouse),以上试剂盒购自德国Miltenyi Biotec公司。Dynabeads Mouse T-Activator CD3/28购自美国Invitrogen公司。BD FACSLysing Solution购自美国BD Biosciences公司。TaqMan®IL-10、IL-4、精氨酸酶Ⅰ(Arginase Ⅰ,AngⅠ)、NOS2、IL-13、IFN-γ、TNF-α molecule probe均购自美国Applied Biosystems公司。
1.2方法
1.2.1日本血吸虫感染小鼠模型的构建 60只BALB/c小鼠随机分为安慰剂组(无尾蚴的生理盐水,10只),正常对照组(10只小鼠)和感染组(尾蚴感染组40只);采用尾蚴腹部敷贴法构建日本血吸虫病小鼠模型。
1.2.2流式细胞术检测模型小鼠体内各个时间点MDSCs的动态比例变化 感染组小鼠分别在第2、4、8、12、16、20周等6个时间点,采用乙醚吸入麻醉,无菌操作摘取小鼠眼球采血,拉颈处死小鼠,分别取骨髓、肝脏和脾脏组织,研磨过滤制备单细胞悬液,流式细胞术检测小鼠体内MDSCs的比例变化。
1.2.3模型小鼠骨髓MDSCs粒系和单核系亚群的免疫磁珠分选及形态学分析 将模型小鼠处死后,无菌操作游离小鼠的股骨和胫骨,剪断两端骨骺,用staining buffer 冲洗出骨髓,制备单细胞悬液。取100 ml staining buffer 重悬106单细胞,加入CD11b抗体和Gr1抗体,然后用流式细胞仪分析和分选。将流式细胞仪(BD FACS Aira)分选出的骨髓初始MDSCs亚群分别取50 000个细胞进行离心涂片,用Wright-Giemsa染色进行形态学分析。
1.2.4模型小鼠骨髓MDSCs亚群对CD4+或CD8+T细胞增殖的影响 流式细胞仪分选出正常小鼠脾脏的CD4+T细胞、CD8+T细胞,CFSE法标记,将从模型小鼠骨髓MDSCs细胞中分选出的粒系和单核系亚群细胞分别与其共培养72 h,流式细胞仪检测CFSE染色的增殖峰,以评价粒系和单核系两个亚群分别抑制T淋巴细胞增殖的能力。
1.2.5Real-time PCR检测模型小鼠骨髓MDSCs亚群相关细胞因子及ArgⅠ、NOS2 mRNA的表达 用实时定量PCR法(Real-time PCR),检测IL-4、IL-10、IL-13、IFN-γ、TNF-α、TGF-β1等细胞因子以及ArgⅠ、NOS2两种酶在骨髓MDSCs亚群细胞中的mRNA表达水平。以GAPDH mRNA的表达为内参照。Real-time PCR引物由上海生工合成,Mouse IL-4:sense 5′-CATGGAGCTGCAGAGACTCTTT-3′,antis-ense 5′-TGCATGATGCTCTTTAGGCTTTC-3′;Mouse IL-10:sense 5′-ATTTGAATTCCCTGGGTGA-GAAG-3′,antisense 5′-CACAGGGGAGAAATCGATGACA-3′; Mouse IL-13:sense 5′-GTCCCTGGTGTTCTTCCT-GATAC-3′,antisense 5′-CAGCACTACAGAGTCGG-TTTCC-3′;Mouse TGF-β:sense 5′-TGATACGCCTGAGTGGCTGTCT-3′,antisense 5′-TTTGCTGTCACA-AGAGCAGTGA-3′;Mouse IFN-γ:sense 5′-GGCACAGTCATTGAAAGCCTAGA-3′,antis-ense 5′-GTCACCATCCTTTTGCCAGTTC-3′;Mouse ArgⅠ:sense 5′-TGTGGTGGCAGAGGT-3′,antisense 5′-GAGGGACGTATAGACG-3′;Mouse NOS2:sense 5′-TCCATGC-TAATGCGAAAG-3′,antisense 5′-ATGCGGAAGTTGTGG-3′;Mouse GAPDH:sense 5′-TGGAGAAACCTGCCAA-GTATGA-3′,antisense 5′-CTGTTGAAGTCGCAGGAG-ACAA-3′。
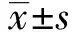
2 结果
2.1流式细胞术检测小鼠体内MDSCs 正常对照组小鼠与安慰剂组相比,MDSCs累积无显著差异,表明外界饲养环境及安慰剂处理不会导致小鼠的MDSCs累积(图1)。而连续监测日本血吸虫感染小鼠在第2、4、8、12和16周骨髓、脾脏、肝脏和外周血的MDSCs,可见比例不断增加,在第8周达到高峰,至第20周时仍维持在较高水平(图2)。

图1 正常组和安慰剂组小鼠不同组织器官MDSCs的比例
2.2日本血吸虫感染模型小鼠骨髓MDSCs粒系(G)和单核系(M)亚群的免疫磁珠分选 通过磁珠分选法从小鼠骨髓MDSCs中分离纯化CD11b细胞后,并运用流式细胞技术分析纯化细胞中CD11b和Gr1细胞含量。结果发现所分离的细胞中MDSCs细胞含量约为99%,其中CD11b+Gr1+双阳性细胞占比94.5%。进一步通过ly6C抗体分选出CD11b+Gr1+亚群中单核系MDSCs(CD11b+Gr1+Ly6C+)亚群(M亚群)和粒系MDSCs(CD11b+Gr1+Ly6C-)亚群(G亚群)(图3)。
图3 小鼠骨髓MDSCs粒系(G)和单核系(M)亚群的磁珠分选

图2 日本血吸虫感染模型小鼠体内MDSCs的动态比例变化

图4 模型小鼠骨髓MDSCs亚群细胞的形态学分析(Bar=50 μm)
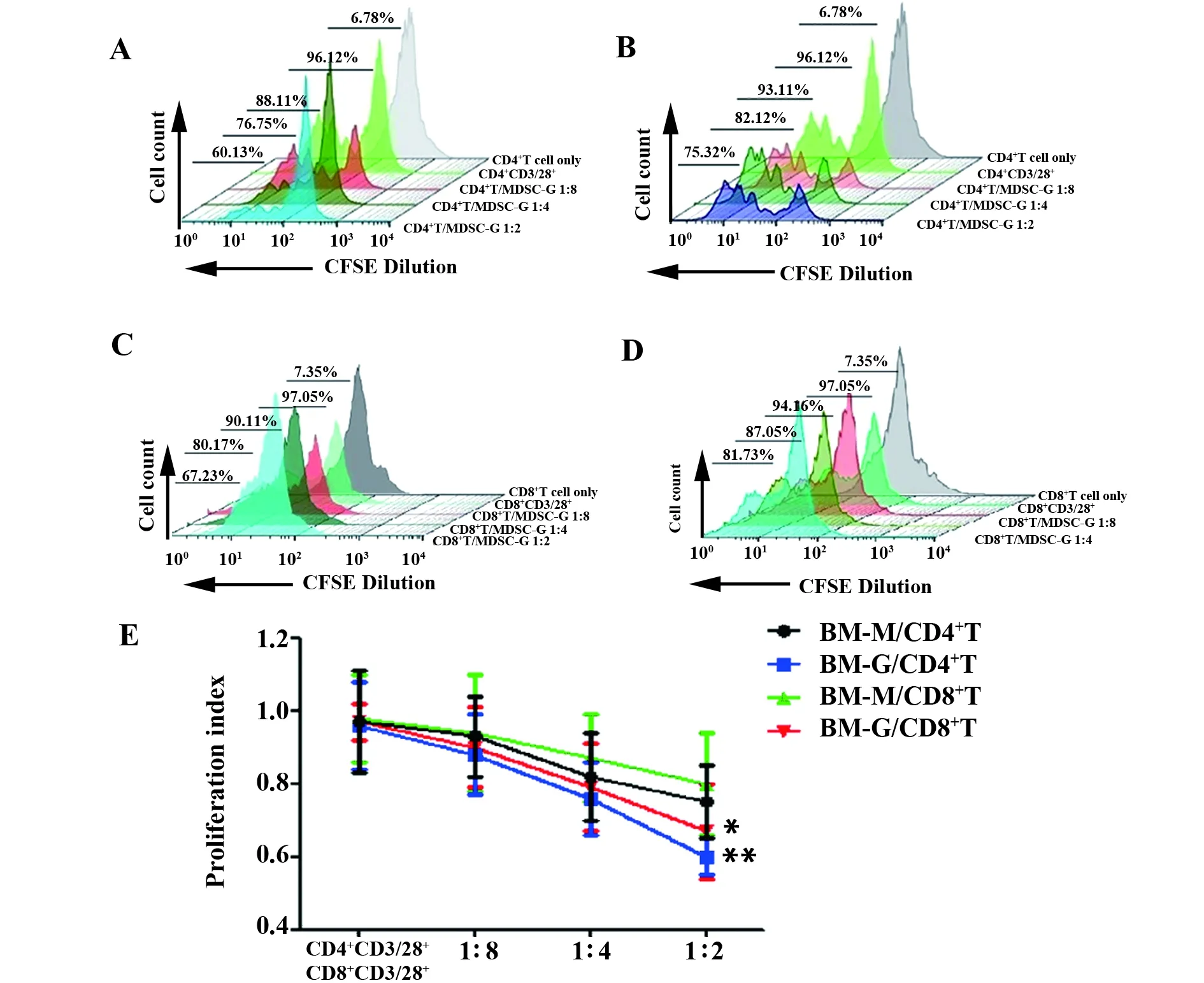
图5 不同浓度MDSCs亚群对CD4+T和CD8+T细胞的增殖抑制作用
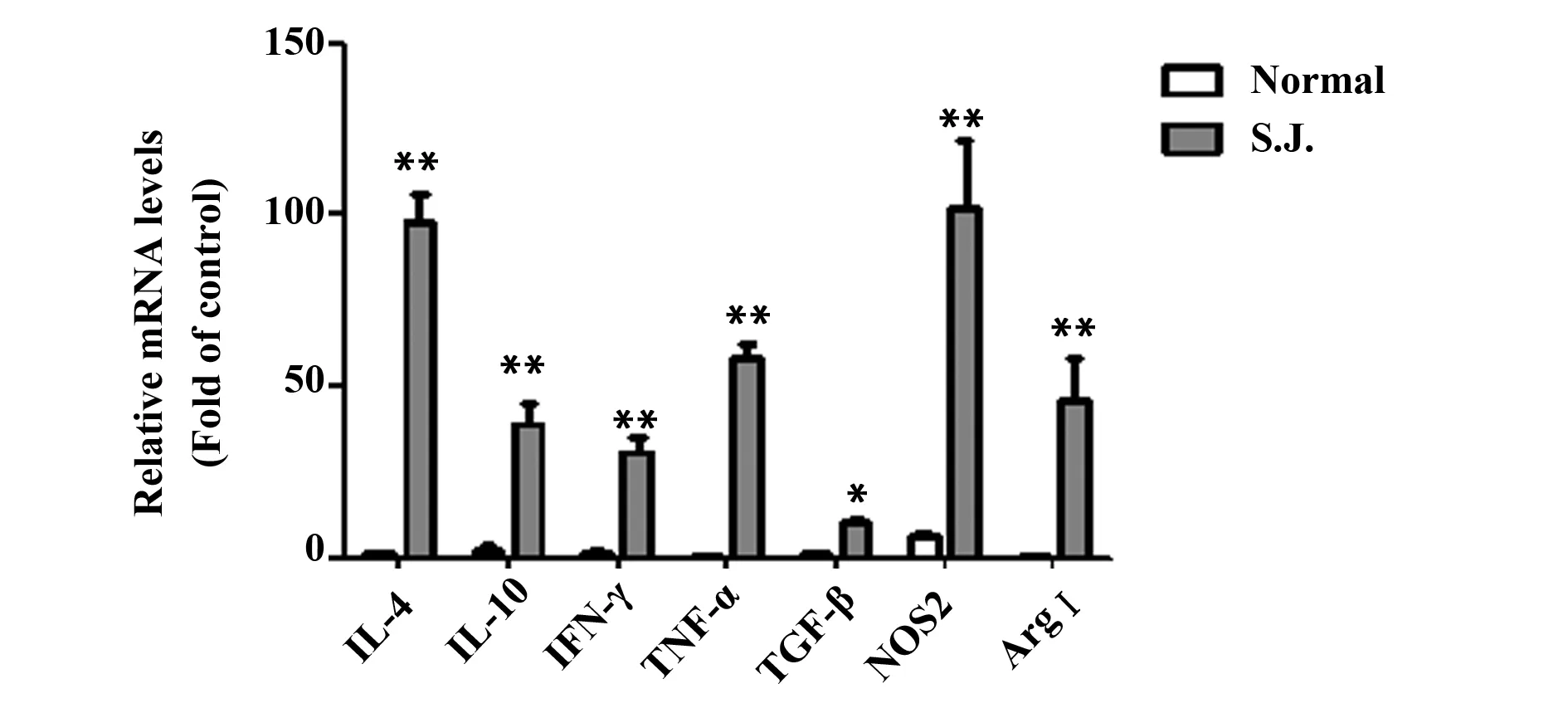
图6 血吸虫感染模型小鼠中相关细胞因子和酶基因的mRNA表达
2.3日本血吸虫感染模型小鼠骨髓MDSCs粒系和单核系亚群细胞的形态学鉴定 将分离纯化的模型小鼠骨髓MDSCs亚群用Wright-Giemsa染色,镜下显示G亚群细胞体积较大,表现为环形核和分叶核的幼稚粒细胞,核质较大;M亚群细胞体积较小,表现为圆形核和椭圆形核的幼稚单核细胞,核质较大。见图4。
2.4血吸虫感染模型小鼠在第8周时骨髓MDSCs粒系和单核系亚群对CD4+T和CD8+T细胞的增殖抑制 采用细胞内荧光染料CFSE标记正常小鼠脾CD4+T细胞和CD8+T细胞,预先用Dynabeads CD3/28-T-Activator 活化CD4+T细胞、CD8+T细胞,取血吸虫感染模型小鼠体内MDSCs在第8周时的累积高峰,与骨髓MDSCs粒系(BM-G)和单核系(BM-M)亚群共培养,72 h后,流式检测CFSE染色的增殖峰,发现骨髓MDSCs细胞中G亚群和M亚群均能不同程度地抑制CD4+T细胞、CD8+T细胞的增殖;并且随着共培养体系中G细胞和M细胞数量比例增加而抑制作用逐渐增强(图5)。
2.5血吸虫感染模型小鼠在第8周时骨髓MDSCs的相关细胞因子及ArgⅠ、NOS2两种酶的实时定量PCR检测 Real-time PCR方法检测模型小鼠在第8周时骨髓MDSCs 细胞的抑制性细胞因子及NOS2、ArgⅠ的基因表达水平。与正常组对照,其差异有统计学意义(图6)。
3 讨论
在各种不同的肿瘤或感染动物模型中,骨髓、血液、脾脏及肿瘤部位均会有MDSCs的增加[8,9],在寄生虫感染中,有关 MDSCs 的研究,近几年也有少量报道[6,7]。为此本实验构建了日本血吸虫感染BALB/c 小鼠模型,来研究MDSCs在寄生虫感染中的免疫抑制作用。本研究分离制备出模型小鼠骨髓、脾、肝、血4种组织的单细胞悬液,流式细胞术检测MDSCs在第2、4、6、8、12、16、20周的动态比例变化,与对照组正常小鼠骨髓、脾、肝、血液等组织中的CD11b+Gr-1+表型的MDSCs细胞相比较;发现在4种组织中MDSCs细胞均有不同比例的升高,尤其在血吸虫病的慢性期(肉芽肿病变期)升高显著。表明在血吸虫感染病程中,骨髓MDSCs大量积聚到外周器官参与抑制机体免疫反应,避免过度免疫反应对重要脏器的损害,以实现对机体的保护。
为了进一步认识MDSCs细胞,本研究对分选出的MDSCs细胞进行了形态学分析,通过免疫磁珠分离方法分离出的CD11b+细胞中约有97%共表达CD11b和Gr-1表型(图1)。形态学上可看到这些细胞形态大小不一,胞质呈淡蓝色,细胞核呈分叶形、环形、椭圆形和多形核(图2)。再根据Ly6G的表达不同分选出单核系MDSCs(CD11b+Gr1+Ly6G-)亚群(M亚群)和粒系MDSCs(CD11b+Gr1+Ly6G+)亚群(G亚群)。采用流式细胞仪分选分离出模型小鼠骨髓MDSCs的两个亚群,分别将其与CD4+T或CD8+T细胞共培养,发现G细胞亚群与M细胞亚群均可以抑制CD4+T或CD8+T淋巴细胞增殖,且抑制作用呈剂量依赖性,以G细胞亚群的抑制作用更强大,表明因感染因素刺激导致体内增加的骨髓MDSCs在体外也具有免疫抑制作用。
既往研究显示,病理条件下MDSCs发挥免疫抑制作用的机制是其能够分泌和产生大量TGF-β、诱导型一氧化氮合酶(Inducible nitric oxide synthase,iNOS)和ArgⅠ,以及通过细胞间相互作用促进T细胞凋亡和CD3ζ的表达,妨碍T细胞活化增殖等功能[10-12]。此外,MDSCs还可以分泌IFN-γ、IL-4、IL-13和IL-10等细胞因子,这些细胞因子可以调节机体Th1细胞向Th2分化从而辅助MDSCs营造抑制性环境,另外还可增加NOS2、ArgⅠ和血红素氧化酶1(HO-1)的表达,从而消耗机体内环境的精氨酸并产生NO及其他代谢产物,最终抑制T细胞的功能[13-15]。为此本研究也检测了血吸虫感染模型小鼠体内的骨髓MDSCs的相关细胞因子IL-4、IL-10、IL-13、IFN-γ及TGF-β1的mRNA表达水平,同时检测了NOS2和ArgⅠ的mRNA表达水平。Real-time PCR检测结果显示各个细胞因子及两种酶均较对照组的正常小鼠有不同程度的升高,其差异有显著性,表明上述因素参与了模型小鼠骨髓MDSCs的免疫抑制作用。
本研究通过构建日本血吸虫感染小鼠模型,研究了MDSCs在寄生虫感染领域中的作用。MDSCs是维持机体免疫平衡的因素之一,了解其亚群的分化和作用机制将有助于制定寄生虫感染的防治策略。
猜你喜欢
中国循证儿科杂志(2022年3期)2022-12-15
今日健康(2022年3期)2022-11-21
广东海洋大学学报(2022年2期)2022-03-31
井冈山大学学报(自然科学版)(2022年2期)2022-03-31
中国人兽共患病学报(2022年2期)2022-03-18
南京医科大学学报(自然科学版)(2020年2期)2020-03-27
医学信息(2019年19期)2019-11-15
中国食用菌(2019年7期)2019-08-13
新疆农业科学(2019年4期)2019-07-18
家庭用药(2018年3期)2018-03-23