
柴胡皂苷d对离体蟾蜍心脏功能的影响
2019-02-15 09:05陈静孙浪罗林邓欢廖富源李春臣罗倩钰陈卫杨春燕张建武
中国老年学杂志 2019年3期
陈静 孙浪 罗林 邓欢 廖富源 李春臣 罗倩钰 陈卫 杨春燕 张建武
(川北医学院 1药学院药物研究所,四川 南充 637007;2麻醉系;3影像学院;4基础医学院机能实验中心)
柴胡始载于《神农本草经》,为伞形科柴胡属植物,按性状不同分别习称北柴胡和南柴胡,其味苦、辛,性微寒,归肝胆经,具有解表退热,疏肝解郁,升举清气之功效〔1〕。柴胡的化学成分较复杂,迄今为止,已报道其含有皂苷、黄酮、挥发油、甾醇、香豆素、糖类、木脂素、有机酸、多炔类及生物碱等成分,柴胡皂苷是柴胡的主要生物活性成分〔2〕。目前对于柴胡皂苷的化学成分研究较为深入,已从柴胡属植物中分离出90多种柴胡皂苷,其中柴胡皂苷a、柴胡皂苷c、柴胡皂苷d 含量较高,药理实验证明柴胡皂苷d的活性最为明显〔3〕。近年来的研究表明,柴胡皂苷d的药理作用较为广泛,包括抗过氧化损伤、抗纤维化、抗炎、抗肿瘤、保肝、镇静及抗癫痫等作用〔4〕。但目前尚未有柴胡皂苷d对蟾蜍心功能影响的报道,本文观察柴胡皂苷d对蟾蜍心功能的影响,并探讨其影响机制。
1 材料与方法
1.1实验动物 健康蟾蜍100只,雌、雄不限,体重110~130 g,由川北医学院实验动物中心提供,实验动物许可证编号SYXK(川)2013-076。
1.2药品、药材 盐酸维拉帕米购自上海禾丰制药有限公司,规格:2.5 mg/ml,批号:43161001;盐酸肾上腺素购自远大医药(中国)有限公司,规格:1 mg/支,批号:161408;艾司洛尔购自齐鲁制药有限公司,规格:0.2 g/ml,批号:161204。柴胡皂苷d由中国科学院成都生物研究所李甫博士提供;任氏液由川北医学院机能实验中心配制。
1.3主要试剂与仪器 肌张力换能器为四川成都泰盟公司生产,电子天平为上海医用激光仪器厂生产,BL-420生物机能实验系统为四川成都泰盟公司生产,加样枪购自芬兰biohit。
1.4离体蛙心标本的制备 参考文献〔5〕制备离体蛙心标本。用斯氏法蛙心插管取1只蟾蜍,破坏脑和脊髓,将其背位固定于蛙板上暴露心脏后在左右主动脉下穿一线,将心脏翻至头端,在背面结扎前后腔静脉;在左主动脉下穿一线,距动脉圆锥2~3 mm处结扎;再从左右主动脉下方穿一线,打一个活结留作固定插管用。用眼科剪在动脉圆锥前端处向心脏剪一斜口,将盛有林格液的蛙心插管由此口插人心室腔内,连同近旁血管一起结扎固定,将心脏离体;用林格液反复冲洗心室内的余血,至插管内灌流液完全澄清无色后,保持插管内的灌流液1 ml,划线作标记。
1.5离体蟾蜍心活动记录 将蟾蜍心经蛙心夹和丝线与肌张力换能器相连,通过BL-420 生物机能实验系统记录离体蟾蜍心心搏曲线。
1.6动物分组及给药 取60只蟾蜍,分为①空白对照组:依次在蛙心套管中加入0.9、0.1 ml林格液;②柴胡皂苷d不同浓度组(实验组):在蛙心套管中分别加入0.9 ml林格液和0.1 ml终浓度分别为0.004 5、0.009 0、0.018 0 mmol/L的柴胡皂苷d;③维拉帕米组:在蛙心套管中同时加入0.9 ml林格液和0.1 ml的0.011 0 mmol/L维拉帕米,使维拉帕米终浓度为0.001 1 mmol/L;④维拉帕米+柴胡皂苷d组:先在蛙心套管中加入0.9 ml林格液和0.1 ml的0.011 0 mmol/L维拉帕米,5 min后彻底灌洗,再在蛙心套管中加入0.8 ml林格液、0.1 ml的0.011 0 mmol/L维拉帕米和0.1 ml的0.070 0 mmol/L柴胡皂苷d; ⑤艾司洛尔组:依次在蛙心套管中加入0.9 ml林格液和0.1 ml的0.12 mmol/L艾司洛尔,使艾司洛尔终浓度为0.040 6 mmol/L;⑥艾司洛尔+柴胡皂苷d组:先在蛙心套管中同时加入0.9 ml林格液和0.1 ml的0.406 2 mmol/L艾司洛尔,5 min后彻底灌洗,再在蛙心套管中同时加入0.8 ml林格液、0.1 ml的0.406 2 mmol/L艾司洛尔和0.1 ml 0.070 0 mmol/L柴胡皂苷d,每组10只。用BL-420生物机能实验系统测定每次给药后5 min内离体蛙心心肌的收缩力及收缩频率的变化情况,记录蛙心心率和心力峰峰值(表示心肌收缩力的大小)。
1.7统计学方法 采用SPSS19.0软件行t检验。
2 结 果
2.1不同浓度的柴胡皂苷d对离体蛙心心率、心肌平均收缩力和心肌张力峰峰值的影响 终浓度为0.004 5、0.009 0、0.018 0 mmol/L的柴胡皂苷d对离体蛙心心率影响差异无统计学意义(P>0.05),见表1。在蛙心套管分别加入终浓度为0.004 5、0.009 0、0.018 0 mmol/L的柴胡皂苷d后,离体蛙心心肌平均收缩力逐渐上升,约在给药后35 s时达到峰值,与给药前比较,差异有统计学意义(P<0.05,P<0.01),且随柴胡皂苷d浓度的增加,柴胡皂苷d对蛙心心肌平均收缩力提高率〔(柴胡皂苷d灌流后35 s平均收缩力-给药前平均收缩力)/给药前平均收缩力×100%〕逐渐升高,具有明显的药物浓度依赖性;给药35 s后离体蛙心心肌平均收缩力逐渐下降,约在5 min时低于其给药前的心肌收缩力,但差异无统计学意义(P>0.05),显示柴胡皂苷d对离体蛙心心肌收缩力呈现出一个典型的先增强后减弱的作用。见表2。在蛙心套管中分别加入终浓度为0.004 5、0.009 0、0.018 0 mmol/L的柴胡皂苷d后,离体蛙心心力峰峰值逐渐上升,约在给药后35 s时达到峰值,与给药前比较,差异有统计学意义(P<0.05,P<0.01);给药35 s后,离体蛙心心力峰峰值逐渐下降,约在5 min时蛙心心力峰峰值显著低于给药前(P<0.05,P<0.01),显示柴胡皂苷d对离体对蛙心收缩力呈现出一个典型的先增强后减弱的作用;但随着柴胡皂苷d浓度的增加,柴胡皂苷d对蛙心心力峰峰值提高率〔(柴胡皂苷d灌流后35 s时心力峰峰值-给药前心力峰峰值)/给药前心力峰峰值×100%〕逐渐升高,具有明显的药物浓度依赖性。见表3。
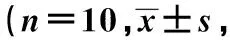
表1 不同浓度的柴胡皂苷d对离体蛙心心率的影响次/min)
1)给药后35 s与给药前比较,2)给药后5 min与给药前比较,表2、3同
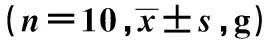
表2 不同浓度的柴胡皂苷d对离体蛙心心肌平均收缩力的影响
表3 不同浓度的柴胡皂苷d对离体蛙心心力峰峰值的影响
2.2维拉帕米单用及其与柴胡皂苷d合用对离体蛙心心力峰峰值的影响 在蛙心套管中单独加入终浓度为0.001 1 mmol/L维拉帕米后35 s和5 min时,离体蛙心心力峰峰值均下降,与给药前差异有统计学意义(P<0.05)。在蛙心套管中同时加入浓度为0.001 1 mmol/L维拉帕米和终浓度为0.009 0 mmol/L的柴胡皂苷d后,离体蛙心心力峰峰值逐渐升高,约在给药后35 s时达到峰值,与给药前比较,差异有统计学意义(P<0.05);给药35 s后,离体蛙心心力峰峰值逐渐下降,约在5 min时,其心力峰峰值低于给药前,差异有统计学意义(P<0.05),离体蛙心收缩力呈现出一个典型的先增强后减弱作用。在给药约35 s时,柴胡皂苷d和维拉帕米合用与单独用维拉帕米,离体蛙心心力峰峰值的两者比较差异有统计学意义(t=-5.794,P=0.001)。见表4。
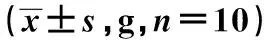
表4 维拉帕米单用及其与柴胡皂苷合用对离体蛙心心力峰峰值的影响
1)给药后35 s与给药前比较,2)给药后5 min与给药前比较;与维拉帕米0.001 1 mmol/L比较:3)P<0.01
2.3艾司洛尔单用及其与柴胡皂苷d合用对离体蛙心心力峰峰值的影响 在蛙心套管中单独加终浓度为0.040 6 mmol/L艾司洛尔后35 s和5 min时,离体蛙心心力峰峰值均下降,与给药前差异有统计学意义(P<0.05)。在蛙心套管中同时加入浓度0.040 6 mmol/L艾司洛尔和终浓度0.009 0 mmol/L的柴胡皂苷d后,离体蛙心心力峰峰值逐渐升高,约在给药后35 s达到峰值,与给药前比较,差异有统计学意义(P<0.05);给药35 s后,离体蛙心心力峰峰值逐渐下降,约在5 min时,其心力峰峰值低于给药前,但差异无统计学意义(P>0.05),离体蛙心蛙心收缩力呈现出一个典型的先增强后减弱的作用。在给药约35 s时,柴胡皂苷d和艾司洛尔合用与艾司洛尔单用比较,离体蛙心心力峰峰值比较差异有统计学意义(t=-4.234,P=0.005)。见表5。
表5 艾司洛尔单用及其与柴胡皂苷d合用对离体蛙心心力峰峰值影响
1)给药后35 s与给药前比较,2)给药后5 min与给药前比较;与艾司洛尔0.040 6 mmol/L比较:3)P<0.01
3 讨 论
本实验表明柴胡皂苷d对离体蛙心心肌收缩力具有增强的作用。维拉帕米和艾司洛尔分别为钙通道阻滞剂和速效选择性β1受体阻断剂〔6,7〕,主要用于心绞痛、心律失常、高血压等心血管疾病的治疗〔8〕,艾司洛尔还可改善心肌损伤、抑制炎症反应提高预后〔9〕。本研究中在蛙心套管中单独加入维拉帕米和艾司洛尔时,两者均使离体蛙心心力峰峰值下降;但当与柴胡皂苷d合用时,均使离体蛙心心肌收缩力增强。其原因是:单独用时,维拉帕米是通过抑制心肌细胞外钙离子的内流,降低胞内钙离子的浓度,使心肌收缩力降低;艾司洛尔是通过减弱或取消儿茶酚胺对心脏的兴奋作用,使心肌收缩力减弱〔6,7,10〕。当合用时,柴胡皂苷d通过激动离体蟾蜍心心肌的L型钙通道和心肌细胞上的β1受体,并通过鸟苷酸结合蛋白-腺苷酸环化酶-环磷酸腺苷-蛋白激酶A通路,使心肌细胞内的cAMP水平升高、PKA活性增强,使心肌细胞膜中L型钙通道磷酸化被激活,开放概率增加,进而使平台期Ca2+内流增加,内流的Ca2+又激活连接肌质网膜中的雷诺丁受体,通过钙触发钙释放机制使胞质内钙离子浓度进一步升高,引起正性变力作用〔11,12〕,从而使离体蛙心心肌收缩力增强。这说明对于加柴胡皂苷d 35 s后心肌收缩力下降的原因与相关报道一致〔13,14〕,可能与柴胡皂苷d体外诱导细胞膜通透性改变,从而导致细胞损伤或坏死、能量供应不足有关。细胞膜损伤、膜功能障碍时,Ca2+内流增多,大量钙盐沉积于线粒体,可造成呼吸链中断,氧化磷酸化障碍〔15〕,最终导致ATP产生不足,使心肌收缩力降低。
综上所述,柴胡皂苷d使离体蛙心收缩力的增强可能与柴胡皂苷d激动心肌细胞上的β受体进而激活L型钙通道,促进钙离子内流使胞质内Ca2+增加有关;使蛙心收缩力的减弱可能与细胞膜损伤、引起心肌ATP供能不足有关。
猜你喜欢
少年文艺·开心阅读作文(2019年5期)2019-05-28
读书文摘(下半月)(2018年11期)2018-10-21
少年文艺·开心阅读作文(2017年10期)2017-10-26
中国医学创新(2017年5期)2017-03-16
奥秘(2016年10期)2016-12-17
少年文艺·开心阅读作文(2016年10期)2016-09-10
少年文艺·开心阅读作文(2016年10期)2016-09-10
人力资源管理(2016年6期)2016-08-11
求是学刊(2015年4期)2015-07-30
园艺与种苗(2015年10期)2015-02-27
