
五指山地区海南苏铁种群结构特征与动态
2019-01-04 08:00:42谢春平吴昌魁赖水发王华晨
中南林业科技大学学报 2019年1期
谢春平,吴昌魁 ,付 桂 ,赖水发 ,方 彦,王华晨
(1.南京森林警察学院 刑事科学技术学院,江苏 南京 210023;2.海南省森林资源司法鉴定中心,海南 海口 570203;3.南京林业大学 生物与环境学院,江苏 南京 210037)
种群(Population)是同一物种在一定地质时期所占据地理空间的所有个体的集合[1]。它是群落结构和功能的最基本单位和物种适应的单位,具有数量特征、空间分布特征、遗传特征和系统特征4大特征[2]。这些特征决定了种群结构和种群动态是种群生态学的核心问题[3]。种群结构对了解种群现状及预测种群动态具有重要意义[4-5]。对于木本植物而言,以植株的径级或高度级表示的种群年龄结构特征是一种被广泛采用的方法,它不仅简化了种群生态学的研究过程,而且提供了有价值的种群动态信息[6];它直观反映了种群内不同大小个体数量的分布和配置状况,从而体现种群动态及其发展与演变趋势[7]。生命表是研究木本植物种群结构与动态的另一重要方法[8-9],其所体现出来的死亡率、出生率及存活曲线等,为种群年龄结构、数量动态统计和发展提供了重要信息[10],它也是种群统计的主要内容之一[11]。因此,通过种群年龄结构、生命表等方法揭示特定种群的结构及数量特征,对濒危物种的保护和管理具有重要的理论价值与现实意义。
苏铁类植物被誉为“活化石”,是现存种子植物中最原始的一个类群,具有重要的经济、文化、生态和科研价值[12]。它对研究植物的起源、演化、种子植物区系、古气候变迁、植物遗传多样性和亲缘关系等都具有重要意义[13]。因此,苏铁类植物不仅是《世界野生植物濒危物种红皮书》中所列的重点保护物种,同时苏铁属所有种在中国均被列为I级重点保护植物,已引起了国内外学者的广泛关注[12,14-17]。Borsboom et al.探讨了澳洲苏铁Macrozamia parcifolia种群的结构特征,并对种群的维护管理提出了相应的对策[18];Octavio-Aguilar et al.研究了墨西哥韦拉克鲁斯的泽米苏铁Zamia inermis,指出该地区的苏铁种群属于衰退状态,如不迁地扩繁保护,有灭绝的风险[16];Rao et al.报道了印度安得拉邦Pinjarikonda森林保护区发现的一个新的苏铁Cycas sphaeric种群,并对种群的现状、规模、分布等进行了相应的分析后,指出当前森林环境是适宜苏铁生存的,但同时也说明了该物种迁地保护的必要性[15]。我国学者对攀枝花苏铁C.panzhihuaensis[19]、潭清苏铁C.tanqingii[20]、仙湖苏铁C.fairylakea[17,21]、篦齿苏铁C.pectinata[22]、叉叶苏铁C.micholitzii[23]等均做了相应的研究,并取得了丰硕的成果。这些研究均指出了当前苏铁类植物受到栖息地破坏、病虫害、自然灾害、人为干扰、外来物种竞争等诸多因素的影响[14,24]。因此,对苏铁类植物的保护已刻不容缓。
虽然国内外学者已认识到苏铁类植物保护的重要意义,但对海南苏铁C.hainanensis的研究仍较为鲜见[25-26],而关于种群生态学方面的研究至今未见报道。海南苏铁主要分布在海拔100~1 000 m的海南岛低山地雨林或稀树灌丛中。近年来,其自然生境受到来自农业种植、热带果园、人工林、旅游、森林采伐、盗采挖、放牧等人类活动的严重干扰。海南苏铁的天然分布生境被蚕食,且严重破碎化,但处于这一特殊处境下仍未引起相关部门及学者们的关注。种群结构特征和数量动态研究是保护和恢复海南苏铁的基础工作之一,也是相关部门保护和管理该物种的参考依据。基于此,本研究以海南琼中黎族苗族自治县长征镇分布的海南苏铁为研究对象,通过对其种群年龄结构特征、静态生命表、存活曲线、种群动态数量等的分析,拟解决2个科学问题:1)研究区内的海南苏铁的种群现状如何?2)该种群的发展动态及趋势。通过研究,旨在为该物种的种群恢复策略、种群更新、资源保护等方面提供科学依据。
1 材料与方法
1.1 研究地概况
研究样地设置在海南省琼中黎族苗族自治县境内,该区域位于海南省中部,为生态核心区。该县境内分布有五指山、黎母山、吊罗山、鹦歌岭等自然保护区,生态环境良好。该区域位于热带海洋季风区北缘,雨水充沛,气候温和,四周群山环抱,有独特的山区气候特点。琼中县年均气温22 ℃,1月份均温16 ℃,绝对最低气温为-6 ℃,7月份均温26 ℃,绝对最高温度38 ℃。年均日照时长为1 600~2 000 h,太阳总辐射约为4 579 MJ/m2;年均相对湿度为80%~85%;年均降水量为2 200~2 444 mm,最大年降水量可达5 525 mm,最小年降水量为1 018 mm;年均蒸发量为1 824.1 mm。研究样地内土壤以黄壤土为主。
经2017年踩踏调查后,在海南省琼中县长征镇设置3块30 mh30 m的样地(样地编号依次为A、B、C),各样地具体情况如表1所示。其中,样地A和样地B设置在五指山自然保护区天然林内,其群落乔木层主要有水东哥Saurauia tristyla、小花五桠果Dillenia pentagyna、黄牛木Cratoxylum cochinchinense、枫香树Liquidambar formosana等;灌木层主要有中华水锦树Wendlandia uvariifolia subsp.chinensis、箬竹 Indocalamus tessellatus、粗叶榕Ficus hirta、大沙叶Pavetta arenosa、猪肚木Canthium horridum等;草本层主要有草豆蔻Alpinia katsumadai、卷柏 Selaginella tamariscina、草珊瑚Sarcandra glabra、尖苞柊叶Phrynium placentarium以及其它蕨类植物等。样地C设置在人工桉树林内,其乔木层基本为桉树Eucalyptus spp.,灌木层和草本层与上述2个样地类似。
1.2 样地设置与数据收集
2018年2月,在琼中黎族苗族自治县长征镇干田村附近(五指山自然保护区内及其周边)设置面积为900 m2的3个样地进行调查(见表1)。用手持GPS对各样地进行定位,记录海拔、坡度、坡向,重点对样地内出现的海南苏铁进行每木测量,包括高度、冠幅、长势、茎粗、茎长等,同时对群落内的伴生种及群落情况进行记录。

表1 海南苏铁调查样地的基本情况Table 1 Physical characteristics of the three plots of C.hainanensis in the study area
1.3 分析方法
1.3.1 种群年龄结构的划分
对于多数乔木树种而言,形成层有规律的分生使其胸径与树龄存在一定的线性关系[27]。因此,多数学者在研究极小种群或濒危物种的种群年龄结构时,可采用径级代替龄级的方法[11,28-30]。但是诸如木本蕨[31]、苏铁类[32-33]等植物由于其茎干结构的特殊性是无法用胸径来代表其年龄结构特征的,而是采用高度级或叶片长度等结构特征替代[23]。结合本研究的实际情况,参考汪殿蓓等[17]和田波等[20]的方法,利用高度级结构(Height)替代年龄级结构来分析种群的年龄结构特征。将海南苏铁植株的高度划分为6个龄级,即:I级,0~10 cm;Ⅱ级,10.1~30 cm;Ⅲ级,30.1~60 cm;Ⅳ级,60.1~90 cm;Ⅴ级,90.1~120 cm;Ⅵ级,>120 cm。归纳统计每个龄级的个体数,绘制种群结构分布图,对种群年龄结构进行分析。
1.3.2 静态生命表的编制
静态生命表中所包含的现存个体数a0、匀滑后的 x 龄级的存活数 ax、x龄级开始时标准化存活个体数lx、x到x+1龄级间隔期间内标准化死亡数d、x到x+1龄级间隔期间死亡率qx、x到x+1龄级间隔期间还存活的个体数Lx、x龄级到超过x龄级的个体总数Tx、x龄级个体的生命期望或平均期望寿命ex以及各龄级组致死力Kx等各指标的计算参考金慧等人的方法[34]。由于海南苏铁静态生命表无法满足编制过程中所需的3个理想化假设[35],因此参考解婷婷等人[36]的方法对数据进行匀滑处理。
1.3.3 种群动态的量化分析
以种群结构为基础参数,通过数学方法定量地推导出种群结构动态指数,可有效解决群落结构动态定量比较评价问题,从而克服植物种群结构动态比较评价中粗放的等级归类划分的缺陷[37]。该方法包括了种群整体的年龄结构动态变化指数(Vpi)和未来外部干扰情况下种群的年龄结构变化指数(V′pi),这2个指数在种群动态研究中被广泛采用[2,11,34-35]。具体计算方法见参考文献[37]。
2 结果与分析
2.1 种群年龄结构特征
在所调查样地中,共计有135株海南苏铁出现;其中样地A、B和C的种群密度分别为289、633和 589株·hm-2,整体种群密度为 500 株·hm-2。从图1种群结构分布来看,3个样地所表现的种群结构有所不同,主要表现为金字塔型和纺锤型。样地A种群结构为金字塔型,其中I龄级占样地内所有个体数的50%,个体数随着龄级的增大逐渐减少;II至VI龄级所占比例分别为23.1%、11.5%、7.7%、3.8%和3.8%。样地B和样地C种群结构接近于纺锤形或称之为非典型金字塔型[35],即整体上中低龄级种群数量较多;其中,样地B和样地C的低龄级(I级和II级)和中龄级(III级和IV级)分别占种群数量的54.1%、35.8%和40.4%、41.5%。同时,研究区内海南苏铁的整体种群结构也表现为非典型金字塔型,I~VI级个体数所占比例分别为25.2%、25.9%、22.2%、14.8%、5.9%和5.9%,整体上仍是低龄级和中龄级个体占有优势。
2.2 生命表分析
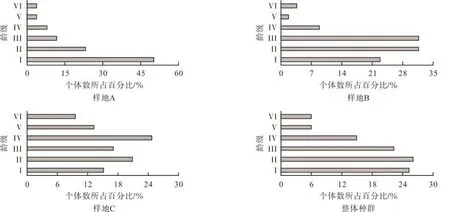
图1 不同样地海南苏铁种群的龄级结构Fig.1 The age structure of C.hainanensis population in different plots
静态生命表无法体现出某一种群的全部生活史,它所体现的是特定时间内多个世代重叠、年龄动态的过程,因此系统误差、死亡率为负的情况在静态生命表中是不可避免的[38]。但负值仍能够说明种群并非处于静态,而是发展或衰退中,仍具有生物学意义[2]。经匀滑修正后得出的海南苏铁静态生命表如表2所示。3个样地整体上表现出随着龄级的增加高龄级个体数量减少的趋势,其中以样地A和样地B的低龄级个体较为丰富。从期望寿命(ex) 值来看,样地A和样地C的植株个体平均生存能力最大值均出现在I龄级,分别为1.72和3.36;而样地B与前两者不同,出现在高龄级V。不同阶段的期望寿命值说明了种群对环境的适应性和生理活动的旺盛情况不同,这与群落环境资源、种内与种间竞争等有直接的关系。从致死力(Kx)和死亡率(qx)值来看,样地C种群各阶段的致死力和死亡率均小于0.2,这说明种群现状较为稳定,有80%以上的个体能够过渡到下一龄级阶段;而样地A和样地B仅有50%左右的个体能够渡过下一龄级,其各阶段的致死力和死亡率均在0.4以上。这一现象可能是样地C属于人工林,其郁闭度低的原因所致。由此不难看出样地环境差别对种群结构差异的影响。
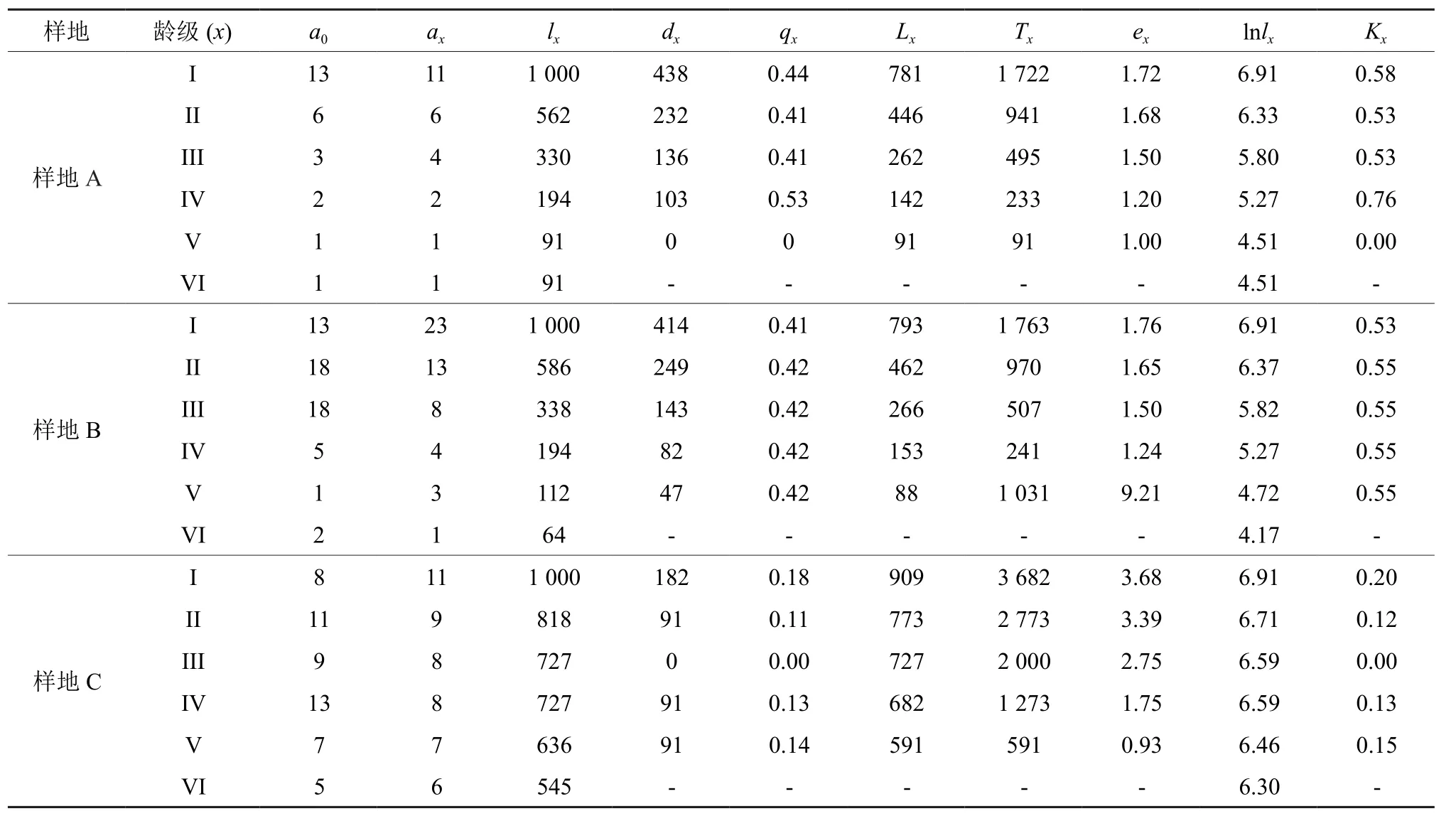
表2 不同样地海南苏铁种群静态生命表Table 2 Stationary life table of C.hainanensis population in different plots
2.3 存活曲线
存活曲线能够直观地反映种群个体在各龄级的存活状况,它是借助于特定年龄组的存活个体数量相对时间作图描述种群动态的常用分析方法[38]。以各龄级为横坐标、存活量为纵坐标绘制3个样地的存活曲线(见图2),同时利用幂函数和指数模型对存活曲线进行拟合(见表3)。按Deevey的划分,一般有3种基本类型:Ⅰ型是凸曲线,属于该型的种群绝大多数都是能活到该物种年龄,早期死亡率较低,但当活到一定生理年龄时,短期内几乎全部死亡;Ⅱ型是直线,也称对角线型,属于该型的种群各年龄的死亡率基本相同;Ⅲ型是凹曲线,早期死亡率高,一旦活到某一年龄,死亡率就较低[36,39]。结合表2及图2可知,样地A从I龄级至III龄级的死亡率趋于相同,说明各阶段死亡率较为平缓,而最高死亡率出现在IV龄级;样地B的存活曲线与样地A较为相似,故这2个样地的存活曲线类型应为II型向III型过渡的中间类型;从曲线结构的整体走向看,样地C的存活曲线亦偏向于II型。
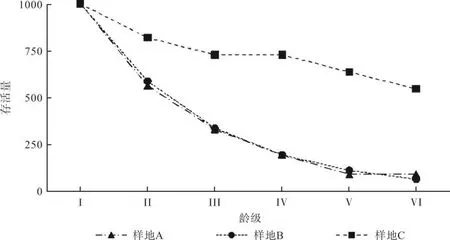
图2 不同样地海南苏铁种群存活曲线Fig.2 Survival curves of C.hainanensis population in different plots
利用Hett和Loucks对Deevey-II型和Deevey-III型曲线符合程度的方法检验[40],即符合Deevey-II型的指数方程Nx=N0e-bx和符合Deevey-III型的幂函数方程Nx=N0x-b,结果如表3所示。指数方程和幂函数方程拟合的结果均达到显著水平(P<0.01);结合R2值发现,3个样地的指数方程均要优于幂函数方程(见表3)。因此,可以认为海南苏铁的存活曲线更趋近于Deevey-II型。

表3 不同样地海南苏铁种群存活曲线检验模型Table 3 Testing models of survival curves of C.hainanensis population in different plots
2.4 种群动态量化分析
利用种群动态量化方法分析种群结构的变化,对种群动态的评价更为客观精确[36]。如表4所示,样地A的种群动态指数V值均≥0,即表明各龄级之间无衰退的迹象;样地B的V1、V5和样地C的V1、V3均出现负值的情况,这表明种群相应龄级个体数量有衰退现象。从整体情况来看,在不考虑外部干扰时,各样地种群均具有较大的Vpi值,分别为48.00%、23.43%和10.52%。在考虑随机干扰时,种群年龄结构动态指数V′pi值具有较明显变小的趋势,尤其以样地C较明显(0.32%)。虽然3个样地的V′pi值均大于0,但样地C偏离0的程度较其他2个样地小,其种群结构的稳定性要差于前两者。随机干扰的敏感性指数 P(即1/[K·min(Sn)])为衡量种群结构对于干扰的敏感性,各样地值依次为0.17、0.17和0.03。结合V′pi值综合分析不难看出,3个样地的海南苏铁均为增长且稳定的种群,但种群的增长性低;同时,样地A和样地B对外界干扰的敏感性要明显强于样地C。
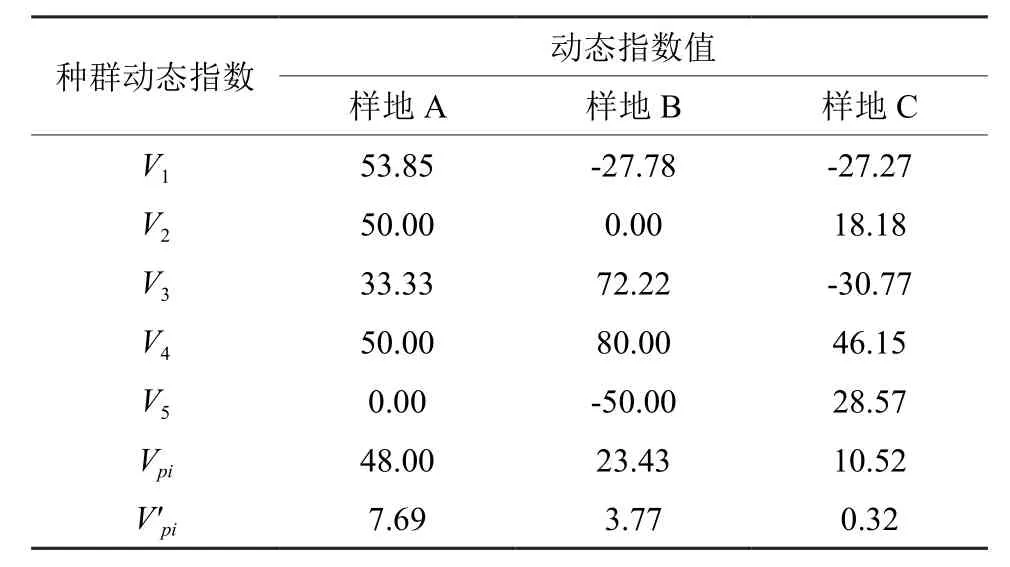
表4 不同样地海南苏铁种群龄级结构动态变化指数Table 4 Dynamic indices of C.hainanensis population and size structure in different plots
3 讨论与结论
3.1 讨 论
虽然长期观测的数据对种群动态的研究较为理想,但短期调查所获得的静态数据也一样可以了解种群的发展趋势[41]。静态数据所展示的是某一特定时期种群的结构特征,可对其近几年或数十年后种群的更新水平、增长率、死亡率等做出有效判断[42]。Wiegand et al.[43]在对沙漠中分布的合欢Acacia raddiana种群的短期研究中指出,更新苗稀缺是影响大树种群结构分布的主要因子,而这种更新苗稀缺的情况也成了该种群的常态特征;Venter和Witkowski[44]对南非分布的猴面包树Adansonia digitata种群结构研究指出,当前的幼龄级树足以在一段时期内维持该种群的现状。因此,利用种群结构特征来探究种群的发展趋势和稳定性已被多数学者广泛采纳[45-47]。
以茎干高度为分析指标,利用“空间序列代替时间变化”的方法对海南苏铁种群结构特征的分析表明,研究地分布的海南苏铁仍处于相对稳定的状态,即在现有条件下可维持种群的持续发展。但从不同样地的种群年龄结构特征发现,环境对海南苏铁的影响较大。样地A和样地B位于保护区天然林内,其受人为干扰因素较低,整体上其低龄级幼树幼苗要优于样地C。样地C在保护区外,其群落上层为桉树,林下受周边村民放牧、薪炭柴采集等活动的影响,故低龄级苗木的比例较前两个样地少。但是,其高龄级数量却远比其他两个样地高,出现这一情况与苏铁属植物自身的生物特性有一定的关系。黄应锋等[21]对深圳梅林仙湖苏铁的种群进行研究时指出,过高的群落郁闭度对仙湖苏铁的衰退有较大的影响。因此,排除人为干扰的影响,适当降低群落郁闭度对海南苏铁种群的发展是有利的。此外,静态生命表也反映出郁闭度对海南苏铁种群不同龄级数量的影响,样地A和样地B过半个体数在I龄级死亡,致死率高,这显示出海南苏铁幼树幼苗对光照需求强烈的特点。但是,在该区域的其他地方,笔者发现有数米高的海南苏铁个体在郁闭度95%以上的群落内生长良好,这从另一方面说明了成年海南苏铁对光的需求要远低于幼年个体。因此,保证有足够的幼龄级个体向成年级过渡是种群持续发展的一个关键环节。
汪蓓殿等人[48]对塘朗山仙湖苏铁群落主要种群的空间分布格局进行了研究,指出该群落内的优势种群混杂有中性树种和阳性树种,人为活动的影响对群落演替发展起着决定性的作用;而减少人为干扰促使群落向地带性顶级群落发展,对仙湖苏铁的保护是有利的。从3个样地种群结构特征来看,处于较高海拔的样地A种群结构为典型的金字塔型,而样地B和样地C为非典型增长种群。形成这一局面可能与人为干扰程度的强弱有一定的关系,海拔越低受人为干扰的程度越大。结合种群龄级结构动态变化指数发现,各龄级的动态变化指数在样地A均未出现负数的情况;样地B和样地C出现负数情况,说明种群数量在龄级之间转换出现了个体数减少的情况。综合分析表明,海南苏铁在现有条件下仍处于一种动态的稳定发展中,样地A的情况要优于其他2个样地。
海南苏铁是国家Ⅰ级重点保护植物,属于濒危物种和极小种群。但根据野外调查结果显示,其当前的野外种群生存面临着几个较为严重的问题:1)外界环境干扰严重。非法贸易一直是苏铁类植物存在的严重问题,在广西百色地区大量的苏铁作为园林观赏植物被盗挖,不法商贩对苏铁类植物进行非法收购贩卖[49],这一情况亦存在云南[22]、四川[24]、贵州[50]等苏铁类植物产区。笔者对当地村民进行走访调查获知,早年有不少园林苗木商贩以不足百元的价格对海南苏铁进行收购,从而使保护区外的苏铁种群大量减少。另外,村民介绍在当地村落附近曾分布有大面积数米高的海南苏铁种群,但通过此次调查发现其所描述的景象已不复存在。由于苏铁类植物具有食用和药用价值,因此被作为食物和药材亦被采集,从而对其种群发展有较大的影响。2)管护缺失。首先,非保护区内的海南苏铁管护不到位,尤其是被不法分子以“蚂蚁搬家”的方式不断盗挖现有的资源,而相关管理部门因人力物力不足,无法做到实时监控管理。其次,森林采伐设计不完善。在森林采伐作业设计时,主管部门仅考虑了采伐面积、出材量等,而林内是否具有保护植物及其他需要关注的问题均未进行考量,由此导致诸如海南苏铁这一类非用材森林产品(non-timber forest product)并未获得应有的关注与保护。笔者在研究地附近的人工林采伐迹地内发现有大量的苏铁出现;而由于采伐作业的需要,大量的海南苏铁因道路、倒木、工棚等被清理或掩埋。此外,周边村民因开荒种地、种植人工经济林等农业生产活动也严重影响了海南苏铁生境的完整与稳定,生境缩减及破碎化已是当前海南苏铁不得不面对的一个严重问题。3)司法实践的窘境。《刑法》规定的“毁坏国家重点保护植物罪”中有一限制前提条件,即“是否主观故意,是否明知”。由于宣传工作缺失及受教育程度限制,多数村民并不知晓海南苏铁为国家I级重点保护植物,而是将其作为普通杂树处理。因此,在该类植物被破坏时,很难对犯罪嫌疑人的行为进行取证,这给打击此类犯罪行为增加了难度。
因此,加大海南苏铁野外种群的保护工作已刻不容缓。结合海南苏铁的生存现状及野外调查结果,笔者建议对海南苏铁采取以下3点保护措施:1)就地保护与迁地保护并举。一方面,对海南苏铁集中分布区域建立保护小区,与当地村民签订相应的管理与保护协议,同时建立合适的生态补偿机制,让村民自动退出与保护物种对山地资源的竞争。这不仅能够较好地保护现有的海南苏铁资源,而且实现了人与自然的和谐共处。另一方面,如确实因开发建设、森林采伐等外界因素对海南苏铁造成影响的,应及时进行野外救护,进行迁地保护。特别是与科研院所、自然保护组织等进行合作,将无法在野外续存的海南苏铁种群进行移栽。这不仅使得海南苏铁的种质资源得以续存,而且亦可获得更大的保护关注度。2)加大繁育与引种研究。目前,苏铁属推广应用最成功的物种当属苏铁C.revoluta,它已广泛在城市园林中栽植。海南苏铁在形态特征与苏铁近似,其树形高大,枝叶开展,观赏性甚至要优于苏铁。因此,借鉴苏铁的成功经验,首先应对海南苏铁的生物学及生态学特性进行细致的调查研究,了解其对环境的适应性。其次,建立优质的种质资源库,优选母株,并通过人工繁育手段,不断扩大种苗资源,以满足园林市场的需求。3)加强宣传教育力度。宣传教育工作的缺失使得多数村民并未意识到保护海南苏铁的意义及重要性,因此加强宣传教育工作力度,层层抓起是今后保护工作的一个重点。笔者认为可采取以村民小组、行政村、乡镇为单位,层层推进,结合网络、报刊、宣传册、标语等,向当地村民宣传保护国家重点保护植物的必要性。此外,结合具体刑事案例告知村民破坏国家重点保护植物的严重性,从而让村民树立保护意识与法制观,最终实现保护海南苏铁的目的。
3.2 结 论
综合研究地海南苏铁的野外种群动态和生存特征,显示该区域海南苏铁种群的低中龄级植株数量占较大的比例。虽然当前海南苏铁为增长的稳定种群,但种群的增长性低,环境对种群年龄结构格局有一定的影响。生境破坏、人为干扰、非法贸易等是影响海南苏铁种群发展的主要因素。
由于受实验条件、调查时间等客观因素的制约,本研究仅对海南苏铁的种群年龄结构进行了初步的研究,而关于其种群分布格局、性比、生殖生物学等方面均未涉及,这也是该物种将来需要深入研究的主要内容。
致谢:对海南省五指山自然保护区护林员王家贤在野外调查工作中给予的帮助表示诚挚谢意。
猜你喜欢
湖南生态科学学报(2022年4期)2022-11-11 03:44:34
数学物理学报(2021年6期)2021-12-21 06:24:52
河南农业大学学报(2020年2期)2020-05-22 02:56:30
数学物理学报(2019年5期)2019-11-29 07:47:04
山西林业科技(2019年1期)2019-05-27 12:24:52
天然产物研究与开发(2019年1期)2019-03-01 05:41:14
红蜻蜓·低年级(2017年2期)2017-03-29 17:21:58
数学年刊A辑(中文版)(2016年1期)2016-10-30 01:46:20
中央民族大学学报(自然科学版)(2016年1期)2016-06-27 07:54:26
中央民族大学学报(自然科学版)(2016年2期)2016-06-27 01:29:00
