
作物水分利用效率研究方法及尺度传递研究进展*
2019-01-03 03:13姜寒冰张玉翠任晓东要家威沈彦俊
中国生态农业学报(中英文) 2019年1期
姜寒冰, 张玉翠, 任晓东,3, 要家威, 沈彦俊**
作物水分利用效率研究方法及尺度传递研究进展*
姜寒冰1,2, 张玉翠1, 任晓东1,3, 要家威1,2, 沈彦俊1**
(1. 中国科学院农业水资源重点实验室/河北省节水农业重点实验室/中国科学院遗传与发育生物学研究所农业资源研究中心 石家庄 050022; 2. 中国科学院大学 北京 100049; 3. 青海师范大学生命与地理科学学院 西宁 810000)
提高作物水分利用效率(WUE)是缓解农业生产水资源匮乏压力的有效途径, 而水分利用效率尺度传递是各尺度WUE相互表征、验证并应用于实际生产的基础。本文概述了作物叶片、植株、群体尺度WUE的主要观测技术, 包括叶片气体交换测定、碳同位素判别、桶栽称重、涡度相关观测等, 其中碳同位素判别法为研究作物水分利用的长期累积效应提供了新的思路, 且适用于多个尺度; 总结了各尺度WUE的影响因子及作物耗水的生理机制, 阐明各尺度WUE均受气孔导度调控。讨论了叶片到植株、叶片/植株到群体的尺度传递的可行性, 集中分析了尺度传递的主要限制因素, 指出叶片到植株的传递研究难点集中于叶片分布和光分布的不确定性、植物夜间呼吸和蒸腾以及植物适应环境的生理调节机制等过程; 而叶片/植株到群体的传递研究主要受冠层形态学差异、冠层阻力、土壤蒸发及植物同化物分配机制等限制。最后总结了尺度传递方法的现有研究成果。目前作物WUE尺度传递主要依靠模型的完善和观测手段的提高, 叶片到单株的尺度传递需关注日间与夜间耗水的分离及作物各部分的光合特性; 叶片/单株到群体的传递可先明确蒸散结构, 了解耗水特征, 再以气孔导度和冠层导度的关系为切入点, 利用模型探究传递机制。
水分利用效率; 尺度传递; 碳同位素; 气孔导度
当今社会不断增长的人口和气候变化给农业生产带来了空前挑战。而农业生产消耗大量的水资源, 据相关统计农业用水占到人类用水总量的2/3以上[1]。为满足世界未来的粮食安全和可持续性发展需求, 保持粮食产量稳定增长的同时尽量缩减农业生产用水, 作物水分利用效率(water use efficiency, WUE)成为农业研究的重要课题。
尺度传递是指不同尺度的水分利用效率相互表征的方法。大多是小尺度向大尺度的拓展, 例如利用叶片水平的瞬时WUE评估群体或区域水平的水分利用状况。就目前的观测手段而言, 群体尺度和植株尺度WUE及其相关参数直接测定难度较大, 尺度传递是叶片水平研究成果应用于实际生产的基础。同时尺度传递也可用于研究利用群体的WUE评估单株水平, 涡度相关、遥感监测等手段观测到的群体WUE表现并不能直接代表单株水平, 由群体到单株的尺度传递在优质种质材料筛选和作物品种评价等方面意义重大。为拓展作物水分利用效率相关理论研究, 并应用于实际生产, WUE的尺度传递与拓展成为相关领域未来研究的热点。
1 不同尺度水分利用效率的定义与观测手段
作物水分利用效率(WUE)是表征作物生产力与耗水量之间定量关系的指标, 可在叶片、植株、群体等多个尺度被观测和研究(表1)。针对不同尺度, 作物WUE具有不同的含义和对应观测手段。叶片水平WUE现存在两种定义, 即瞬时水分利用效率(WUET)、内部水分利用效率(WUEi)[2-4]。瞬时水分利用效率可由净CO2同化速率()和蒸腾速率()表征, WUET=/; 内部水分利用效率由净CO2同化速率()与适应当下水汽压差的气孔导度(s)之比表征, WUEi=s。WUEi受环境因子干扰较少, 可更严谨地量化叶片气孔对碳水交换速率的控制作用。植株尺度水分利用效率(WUEP)为作物全株干物质总量与耗水总量之比, 其中耗水量可用作物总蒸腾量表示[5]。WUEP表征一段时间内作物整株的生物量积累和水分消耗情况, 是研究植株固定CO2的生理生态机制的重要参数。群体尺度WUE可分为农田和冠层两个水平。农田水平的WUE(WUEf)是指整个农田生态系统损耗单位水量所生产的干物质总量(total biomass, TB)或经济产量(yield), WUEf=TB/ET或WUEf=Yield/ET; 冠层水平WUE(WUEc)由生态系统的初级生产力(gross primary productivity, GPP)与蒸散耗水总量(ET)表征。不同表达形式的冠层尺度WUE(WUEc)的变异特征也不尽相同。
作物WUE常用的观测手段包括气体交换法、涡度相关技术、稳定碳同位素技术、遥感监测以及农田水平衡等。叶片尺度WUE常用气体交换法测定, 观测结果反映作物叶片的瞬时生理状况, 多用于研究作物的短期水分利用状况或作物对环境因子改变的生理响应。农田尺度WUE能客观反映大田的水分利用状况, 但直接观测难度较大。近年来, 涡度相关技术迅猛发展, 逐渐成为生态系统潜热通量、感热通量最权威的观测方法。热平衡式茎流计和微型蒸渗仪等的应用也给田间蒸散的观测提供了技术支持。此外, 稳定同位素技术研究的深入为测定作物水分利用效率提供了新思路。植物在光合作用过程中, 因同位素扩散效应和光合酶系统对同位素的分馏作用, 导致植物体内的13C明显低于空气CO2的13C。质谱仪以此为原理测定碳同位素判别值(carbon isotope discrimination,13C)。13C是指植物干物质中稳定性碳同位素比率(13C/12C)相对于大气中用于植物光合作用的13C/12C的度量:
13C=(a-p)/(1+p) (1)
式中:p、a分别为植物干物质和空气中的稳定性碳同位素比率。叶片和植株尺度的WUE均可用13C表征。但碳同位素组成测定技术要求较高, 成本昂贵, 严重限制了碳同位素判别方法在WUE研究中的应用。目前, 相关学者正在尝试寻找13C的替代指标, 如植物体灰分含量、比叶面积、氮含量等, 均与13C和WUE存在一定的相关关系, 均有可能替代13C指示作物的水分利用状况。
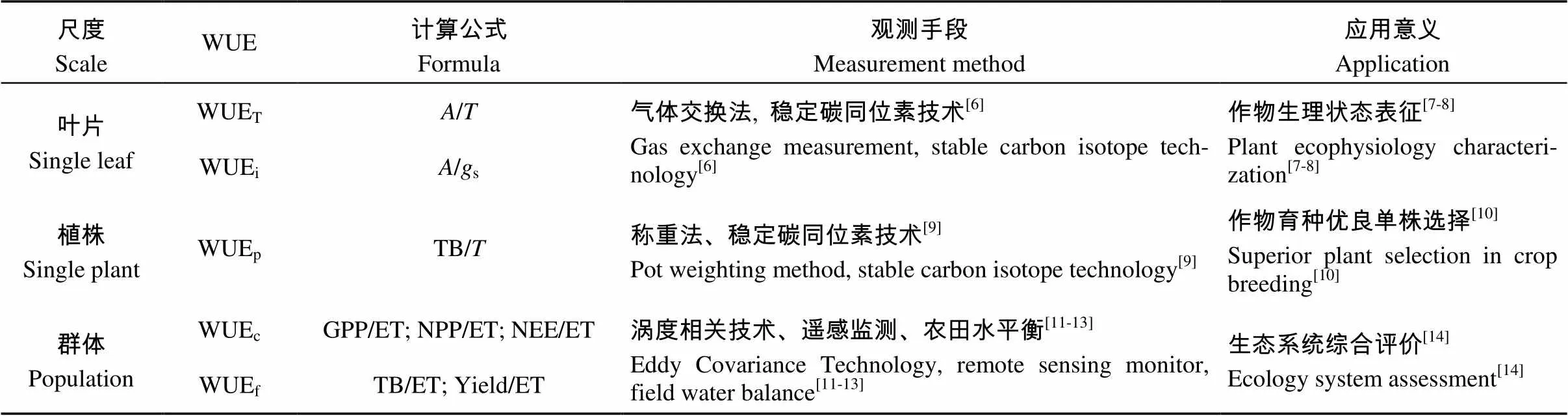
表1 不同尺度水分利用效率(WUE)的定义与观测手段
WUET: 瞬时水分利用效率; WUEi: 内部水分利用效率; WUEp: 植株尺度水分利用效率; WUEc: 冠层水平水分利用效率; WUEf: 农田尺度水分利用效率;: 净CO2吸收速率;: 蒸腾速率;s: 气孔导度; TB: 干物质量; GPP: 初级生产力; NPP: 净初级生产力; NEE: 净生态系统碳交换量; ET: 蒸散耗水量。Yield: 产量。WUET: instantaneous WUE; WUEi: internal WUE; WUEp: WUE at plant scale; WUEc: WUE at canopy level; WUEf: WUE at field scale.: net CO2absorption rate;: transpiration rate;s: stomatal conductance; TB: dry biomass weight; GPP: gross primary productivity; NPP: net primary productivity; NEE: net ecosystem carbon exchange; ET: evapotranspiration.
2 作物多尺度WUE研究现状
2.1 WUE的影响因子
作物WUE由光合作用与蒸腾作用决定, 凡影响作物这两个生理过程的因素都会影响作物WUE[15]。作物的比叶面积、叶片气孔导度、叶片含氮量、根系分布等生理特征会直接影响其光合与蒸腾, 而大气CO2浓度、温度、降水、光照等气象因子会间接调控作物的光合与蒸腾, 进而引起WUE变化。从单株水平来讲, 凡能影响作物生物量和蒸腾量的因素都是植株尺度WUE的影响因子。群体尺度WUE除受以上作物本身生理特性和气候条件的影响外, 还受光合产物分配机制的调控。由于作物水分利用效率尺度拓展对于调控作物生理特性方面关注较少, 故下文主要讨论环境因素。
CO2浓度升高会使各尺度的WUE均随之提高, 其中以叶片水平提高最为明显, 这一结论已被大量研究证实[16-18]。叶片水平上, CO2浓度升高会促使作物气孔开度减小, 同时光合作用增强, 蒸腾速率减弱, 故WUE显著升高, 但光合与蒸腾两个过程哪一个响应更为显著仍存在争议。Rogers等[19]对温室中的作物注入CO2, 结果显示作物光合作用没有明显升高, 认为WUE提高主要是蒸腾速率下降所致; 而王建林等[16]认为光合作用的提高贡献更大。对此目前仍无明确结论。群体尺度生物量随CO2浓度升高会明显增加, 蒸散量有降低趋势, 但变化不明显, 所以群体尺度WUE对CO2的响应幅度远小于叶片尺度[19-20]。实际生产中, 大气CO2浓度升高多与其他环境因素改变相耦合, 如温度、降水、土壤养分等。作物WUE对这些环境因子的复杂组合如何响应仍需进一步探讨。尤其群体尺度WUE对CO2浓度升高的响应过程和变化阈值及产量构成要素变化规律与内在机理等均对WUE的尺度传递研究具有重要意义。
温度变化对作物WUE的影响较为复杂, 不同作物的光合与蒸腾过程对温度的响应趋势不同。针对叶片尺度, Loader等[21]认为温度升高可提高植物的光合能力, 从而提高WUE; 但Nicotra等[22]发现温度更大程度上影响植物的蒸腾作用, 一定程度上可使WUE降低。这表明叶片WUE与温度之间存在显著的负二次相关关系。叶片WUE对温度变化响应的复杂性可能与叶片气孔导度、边界层阻力以及作物光合特性等有关, 这些均是WUE尺度拓展需要重点考虑的因素。
降水状况通过影响作物生育期内的相对湿度和饱和水汽压差(VPD)影响作物WUE。整体来讲, 干旱和半干旱区的作物WUE相对较高[23-24]。有研究[25]表明, VPD是影响作物各尺度WUE的最主要环境因子。Ponton等[26]比较分析了3个生态系统的群体WUE, 发现WUE与VPD具有显著的线性负相关关系, 而且VPD差异是3个生态系统WUE不同的主要原因。另外, 降水情况在一定程度上影响土壤湿度, 土壤水分适度亏缺会促使作物根系产生胁迫信号, 诱导气孔开度降低, 由于光合对气孔开度的依赖性相对于蒸腾较小, 叶片水平的WUE会有所提高。与此同时, 适度的水分亏缺也会增加群体产量, 提高农田尺度WUE[27]。但若水分输入过少, 导致土壤含水量过低时, 作物群体WUE便会降低[28-29], 这说明作物WUE除受气孔调控外, 也受非气孔因素调控, 且WUE在对干旱胁迫的响应上存在阈值。所以, 作物WUE尺度传递模型需考虑饱和水汽压差和土壤含水量的调控作用。
2.2 作物耗水生理机制研究
作物耗水的生理机制主要是在作物蒸腾和光合两个过程的研究基础上展开。叶片尺度和植株尺度的WUE及其对环境因子的响应主要由气孔开度调控[11], 此外也受作物本身光合性能的影响, 如羧化酶活性、腺苷三磷酸(adenosine triphosphate, ATP)含量等。13C能对作物耗水调控的气孔因素和非气孔因素做出综合反映, 是研究作物耗水生理机制的有效途径[30]。
作物WUE由光合作用与蒸腾作用两个生理过程控制, 而植物的光合与蒸腾受到大气CO2浓度、温度、湿度(饱和水汽压差)、光照强度等影响, 且响应机制复杂。大量研究表明作物叶片的光合与蒸腾对环境因子改变的响应具有趋同性, 很难确定两个过程处于怎样的状态会使作物水分利用效果最优。气孔最优化理论实质上就是寻求气孔在复杂环境条件下的最优状态以协调作物的光合与蒸腾过程, 实现高效用水[7]。Farquhar等[31]认为, 当蒸腾速率的偏导数与光合速率的偏导数之比为常量时, 气孔开度在一定时间内最优, 即/=, 其中保持恒定。然而多年研究表明实际生产中很难寻求恒定的值根据气孔最优化理论, Hetherington等[32]研究发现蒸腾速率大于0.4 mol×m-2×s-1的叶片WUE都比较低, 认为蒸腾速率是叶片尺度水分利用效率的关键控制因素[33]。蒸腾速率受气孔开度直接调控, 当气孔开度达到一定程度, 蒸腾速率过高, 作物会出现奢侈蒸腾, 造成水分的无效消耗[34]。奢侈蒸腾的出现主要是由于叶片蒸腾作用与光合作用对气孔开度的敏感性不同, 表现为作物单株尺度耗水量增大。在植株尺度上, 气孔导度是迄今为止发现的影响WUE的最为重要的因素。植株尺度WUE在不同水分条件下的变异性主要来源于不同作物气孔导度对水分条件改变的响应[35]。后有相关生物学家发现诱导植物体基因过表达可促使植株WUE提高, 并将此归因于气孔导度降低[36-37]。植株尺度的WUE虽然受气孔调控, 但与气孔密度和开度相关性不强[38-39]。所以目前利用Li-6400光合仪、气孔计等测定叶片尺度的光合、蒸腾速率, 并以此评估作物全株甚至群落水分利用状况的研究方法理论上是存在缺陷的。
3 作物水分利用效率尺度传递的可行性
就目前的作物水分利用效率研究手段和观测技术发展程度而言, 叶片尺度WUE的相关参数便于测定, 但其仅能反映作物瞬时的水分利用状况; 植株尺度WUE参数可通过盆栽称重或碳同位素判别值进行表征, 但其仅能代表单株作物的水分利用特性, 无法直接应用于农田节水研究; 群体尺度WUE最能客观体现实际生产中的水分利用状况, 但其难以实现准确观测。研究作物WUE的尺度拓展与传递问题成为相关领域未来研究工作的重点和难点。与此同时, 观测手段和相关模型的完善也给尺度传递研究提供了新的途径和数据支撑, 相关研究见表2。
3.1 叶片到植株尺度的水分利用效率拓展与传递
目前针对作物叶片向单株的WUE尺度传递方法研究相对较少, 多是基于叶片尺度的实测数据, 借助统计分析的方法实现。如利用单叶气孔导度获取全株冠层导度, 多是采用光合仪、气孔计等直接测定不同位置的叶片气孔导度, 再以整体平均法、权重法、顶层阳叶分层采样法、有效叶面积指数法等统计得出[43]。光合、蒸腾速率的传递同样如此。这种统计上的尺度转换并没有实现真正意义上的尺度传递或拓展。

表2 不同尺度作物水分利用效率及尺度传递研究成果
作物单株水平WUE的研究, 除盆栽称重或碳同位素方法外, 多在叶片尺度光合、蒸腾速率测定的基础上开展, 但诸多学者发现叶片尺度WUEi和WUET评估植株尺度WUEp的可靠性不确定, 叶片向植株的尺度拓展需要了解作物整体生理生态过程, 但叶片分布的不确定性、个体冠层内部光分布的复杂性以及作物适应环境的自我调节机制等因素给尺度拓展研究带来阻碍。Gulías等[25]以两种多年生草本为试验对象测定在不同水分条件下的叶片气体交换参数、生物量、耗水量以评估其叶片和单株尺度的WUE, 结果表明两尺度WUE相关性并不显著; 另外也有诸多研究得出类似结论[48-49]。阐明叶片与单株尺度WUE相关性差的原因对于实现尺度拓展意义重大。Medrano等[4]通过研究葡萄()叶片WUEi、日尺度累积WUE及植株尺度WUE, 分析了叶片和植株尺度碳水平衡在不同时间和空间位置上的变异特征, 指出叶片在植株冠层中所处位置不同和夜间蒸腾、呼吸效应是造成叶片与植株尺度WUE差异的两大主要因素。叶片到植株的尺度传递需深入两个生理过程: 叶片所处位置对其光能截获量和光合能力的影响, 夜间蒸腾和呼吸造成的碳水消耗情况。
相比于叶片尺度WUEi或WUET, 气孔导度s更适合作为植株尺度WUE的估算指标。在轻度水分亏缺条件下, 气孔导度与单株WUE相关性尤其显著[35,50]。但当水分胁迫较为严重时, 气孔效应不再是作物自身调控的主导因素, 叶片s也不再具有指示植株尺度WUE的功能。气孔导度也是实现叶片向植株尺度传递的重要媒介。
3.2 叶片/植株向群体尺度的WUE拓展与传递
叶片尺度和植株尺度WUE向较大尺度的田间尺度拓展时受到诸多因素限制, 如冠层内部阻力、边界层阻力、土壤蒸发、作物夜间蒸腾、作物呼吸耗水以及植株内同化物分配机制等。这两者之间缺乏相关性是叶片尺度理论研究拓展到群体并应用于指导田间生产的主要限制。Linderson等[51]通过对森林群落进行叶片气体交换测定, 研究叶片尺度WUE的变异性和环境依赖性, 并根据获得的相关关系进行从叶片到冠层的尺度拓展, 证明叶片WUE借助VPD进行归一化后获得归一化WUE(WUEnormleaf), 可成功用于冠层WUE评估, WUEnormleafWUET∙VPD, WUEnormleaf依赖低于500 μmol·m-2·s-1的光合有效辐射, 而不依赖冠层形态和环境因子。因此在均匀冠层作物系统中, 可以依据归一化WUE和模拟冠层内部PAR分布进行从叶片到冠层的尺度拓展。若冠层的VPD和PAR已知, 即可根据WUET评估冠层尺度的WUE。该方法尚未在严重干旱胁迫条件下试用。
叶片和植株尺度WUE主要由叶片蒸腾生理特性影响, 群体尺度WUE受冠层形态学差异影响, 因此叶片尺度与植株尺度WUE正相关关系较为明显, 而两者与群体尺度WUE的相关关系往往不明确。基于此, 黄桂荣等[52]提出猜想: 当作物品种的形态学特征相似、冠层特性差异较小时, 蒸腾和光合等生理特性即为各尺度WUE的主控因素, 此时叶片尺度或植株尺度WUE即可作为表征群体水分利用状况, 或作为该品种大田条件下水分利用状况的评价指标。由于尚存在与此结论和猜想相悖的研究报道, 如Condon等[53]认为作物的蒸腾特性本身即为各个尺度的WUE的主控因素。因此, 以上结论和猜想仍需进一步研究验证。
4 作物各尺度WUE尺度传递方法
作物WUE的尺度传递与拓展研究主要依靠模型模拟和观测手段的提高。叶片尺度是生理生态模型的基本尺度[54]。群体尺度的光合模型或蒸腾模型多是在叶片尺度已有模型或理论的基础上拓展而来, 如大叶模型、双层模型、SMPT-SB模型等。
4.1 叶片到植株尺度传递研究方法
叶片到植株尺度传递的难点在于明确引起其差异的两个关键点——夜间呼吸与蒸腾、作物各部位光合与耗水特性的差异[4], 所以尺度传递相关研究可从以下方面入手。对作物夜间和日间的水分利用状况进行分离, 利用桶栽称重法对植株夜间生物量和蒸腾蒸发量进行测定, 从而研究单株作物在较长时间尺度上的累积WUE与叶片瞬时WUE的差异与联系; 根据水平衡方程对单株作物的水分收支进行量化, 评估作物单株尺度的蒸散耗水、有效水分输入等各水平衡要素及相关环境条件, 为尺度拓展与传递确定边界条件。
此外, 碳同位素技术的发展给单株尺度WUE研究及尺度传递提供了新的思路。作物不同部位可直接取样测定其同位素组成, 进而分析其水分利用状况。13C可反映作物光合作用过程中CO2固定的综合信息, 其表征WUE的生理机制是作物在光合作用过程中, 因同位素扩散效应和光合酶系统对13C的分馏作用, 会导致作物体内的13C明显比空气中偏低。
Farquhar等[54]建立了联系13C值与C间的量化方程:
13C=4.4+22.6(C/C) (2)
WUE=n/r=(C-a)/1.6Δ(3)
式中: Δ指叶片与空气的水蒸气浓度差, 1.6为气孔对水蒸气的传导性转为对CO2的传导性的转换因子。
碳同位素判别值为作物WUE定性和定量评价提供了新的途径, 有望成为叶片到植株尺度WUE尺度传递的连接点或验证指标[55-57]。
4.2 叶片/植株向群体尺度拓展的研究方法
叶片或植株尺度耗水向群体尺度拓展可借助多种观测仪器。借助多点的微型蒸发器和包裹式茎流计方法, 结合涡度相关系统的观测对田块蒸散中土壤蒸发与作物蒸腾进行分离, 通过与植株尺度蒸散结构的对比, 结合叶面积指数明确蒸散结构在尺度拓展中的作用。另一方面以气孔导度与冠层导度的关系作为切入点, 探究单株尺度到群体尺度的WUE的传递机制。
大量冠层阻力模拟基于其与各环境因子间的相关关系, 利用经验系数计算得到, 如Jarvis阶乘模型等。Yu等[58]以叶片尺度气孔导度对光合作用的调控机制为理论基础, 拓展出了冠层阻力计算的具有生理意义的参数化模型。

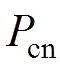
此外还有诸多学者致力于探索利用机理模型实现尺度传递。如Yu等[42]开发了基于气孔行为的光合-蒸腾耦合模型(SMPT-SB模型), 已在玉米()、大豆()等多种作物的叶片尺度进行了应用, 且适用性较好。部分学者尝试了将该模型拓展到冠层尺度。如任传友等[59]尝试构建了适用于温带阔叶林冠层尺度的SMPT-SB模型, 且模拟效果理想。张宝忠等[60]以夏玉米为研究对象, 在叶片和冠层两个尺度率定了SMPT-SB模型相关参数, 并分别以两个尺度的实测数据验证了模型估算结果, 证明该模型在叶片、冠层尺度的适用性均较高, 可为作物WUE的尺度传递提供科学思路。但该模型在表层土壤含水量较高时, 对冠层尺度的蒸散速率模拟偏低, 故仍需将土壤含水量作为影响因素之一耦合进该模型。Zhang等[43]以光合有效辐射作为尺度转换因子, 构建了气孔导度向冠层导度转换的尺度拓展估算模型, 并以所建模型估算冠层ET, 效果较好, 也为WUE的尺度传递提供了方法和依据。Ding等[45]在双源Shuttleworth-Wallace模型的基础上将阳生叶与阴生叶气孔对光照的非线性响应纳入考量范畴, 建立了一个“双源双叶”模型(dual-source dual-leaf model), 可更准确地实现由叶片气孔导度向冠层导度的传递。诸多改良模型均是对作物WUE尺度传递的有效探索, 但相关模型仍存在参数复杂、精度不足、生理意义不明确等缺陷, 尚需深入探索。
5 展望
目前针对各尺度WUE的观测手段、模型评估、调控机制等研究均已有较为成熟的理论成果, 但各尺度间的传递与拓展研究尚不深入。随着碳同位素技术、涡度相关技术、遥感观测技术等观测手段的发展, 植株尺度, 群体尺度, 甚至区域尺度的WUE均可进行直接观测研究。相关学者研究发现, 叶片尺度观测的瞬时WUE并不能直接代表较大空间尺度或较长时间尺度上的水分利用状况, 单叶水平和植株水平的WUE也不能反映农田耗水状况。叶片尺度测定的WUE与植株尺度WUE相关关系的变异性给大田条件下的控制试验和品种筛选等研究带来了严重限制。弥补尺度传递和拓展上的研究空白对于深入了解作物对环境变化的耦合响应机制, 提高农业用水效率意义重大。但由于作物WUE受诸多外界环境因子和内部生理机制的直接或间接影响, 尺度传递需考虑的因素错综复杂, 目前相关领域内仍缺少被广泛认可的权威拓展方法。若要利用各尺度作物WUE研究成果相互表征和验证, 并应用于指导实际生产, 还需在尺度拓展与传递方向进行深入研究。
深化作物WUE尺度传递研究还需完善WUE测定方法, 先进的观测手段是作物WUE尺度传递与拓展研究的基础。例如将双同位素标定法与茎流计相结合。双同位素标定法(13C与18O)不但可精确测定作物WUE, 而且能确定作物固定的水分来源, 反映作物用水格局; 茎流计则是测定作物单株WUE的当今较为成熟和稳定的重要工具。另有遥感数据与地面涡度相关观测相结合等手段。此外, 建立不同尺度WUE联系机理模型是尺度拓展与传递的有效途径。在野外站点设立长期多尺度定位观测, 整合作物各尺度WUE对环境交互变化的响应规律和适应机理, 在试验观测的基础上建立作物叶片、植株和群体多尺度联系的机理模型将是未来作物WUE尺度传递研究的发展方向。
[1] ZHONG H L, SUN L X, FISCHER G, et al. Mission impossible? Maintaining regional grain production level and recovering local groundwater table by cropping system adaptation across the North China Plain[J]. Agricultural Water Management, 2017, 193: 1–12
[2] JONES M M, OSMOND C B, TURNER N C. Accumulation of solutes in leaves of sorghum and sunflower in response to water deficits[J]. Australian Journal of Plant Physiology, 1980, 7(2): 193–205
[3] 曹生奎, 冯起, 司建华, 等. 植物叶片水分利用效率研究综述[J]. 生态学报, 2009, 29(7): 3882–3892 CAO S K, FENG Q, SI J H, et al. Summary on the plant water use efficiency at leaf level[J]. Acta Ecologica Sinica, 2009, 29(7): 3882–3892
[4] MEDRANO H, TOMÁS M, MARTORELL S, et al. From leaf to whole-plant water use efficiency (WUE) in complex canopies: Limitations of leaf WUE as a selection target[J]. The Crop Journal, 2015, 3(3): 220–228
[5] MEDRANO H, FLEXAS J, RIBAS-CARBÓ M, et al. Measuring water use efficiency in grapevines[M]//DELROT S, MEDRANO H, OR E, et al. Methodologies and Results in Grapevine Research. Dordrecht: Springer, 2010: 97–107
[6] 沈芳芳, 樊后保, 吴建平, 等. 植物叶片水平13C与水分利用效率的研究进展[J]. 北京林业大学学报, 2017, 39(11): 114–124 SHEN F F, FAN H B, WU J P, et al. Review on carbon isotope composition (13C) and its relationship with water use efficiency at leaf level[J]. Journal of Beijing Forestry University, 2017, 39(11): 114–124
[7] MINER G L, BAUERLE W L, BALDOCCHI D D. Estimating the sensitivity of stomatal conductance to photosynthesis: A review[J]. Plant, Cell & Environment, 2017, 40(7): 1214–1238
[8] SUN Q, WANG Y S, CHEN G, et al. Water use efficiency was improved at leaf and yield levels of tomato plants by continuous irrigation using semipermeable membrane[J]. Agricultural Water Management, 2018, 203: 430–437
[9] ELLSWORTH P, FELDMAN M, BAXTER I, et al. A genetic link between whole-plant water use efficiency and leaf carbon isotope composition in the C4grass[J]. bioRxiv, 2018: 285676
[10] WANG Y Z, ZHANG X Y, LIU X W, et al. The effects of nitrogen supply and water regime on instantaneous WUE, time-integrated WUE and carbon isotope discrimination in winter wheat[J]. Field Crops Research, 2013, 144: 236–244
[11] MEI X R, ZHONG X L, LIU X Y. Improving water use efficiency of crops by exploring variety differences[J]. Acta Agronomica Sinica, 2013, 39(5): 761–766
[12] TANG X G, MA M G, DING Z, et al. Remotely monitoring ecosystem water use efficiency of grassland and cropland in China’s arid and semi-arid regions with MODIS data[J]. Remote Sensing, 2017, 9(6): 616
[13] 孙志刚, 王勤学, 欧阳竹, 等. MODIS水汽通量估算方法在华北平原农田的适应性验证[J]. 地理学报, 2004, 59(1): 49–55 SUN Z G, WANG Q X, OUYANG Z, et al. Validation of the feasibility of MOD16 algorithm for estimating crop field vapor flux in North China Plain[J]. Acta Geographica Sinica, 2004, 59(1): 49–55
[14] RAHMAN T, LIU X, HUSSAIN S, et al. Water use efficiency and evapotranspiration in maize-soybean relay strip intercrop systems as affected by planting geometries[J]. PLoS One, 2017, 12(6): e0178332
[15] 李东晓, 王红光, 张迪, 等. 水分亏缺对不同小麦品种矿质元素吸收分布及水分利用的影响[J]. 中国生态农业学报, 2017, 25(10): 1475–1484 LI D X, WANG H G, ZHANG D, et al. Effect of water deficit on mineral element absorption, distribution and water utilization by different wheat varieties[J]. Chinese Journal of Eco-Agriculture, 2017, 25(10): 1475–1484
[16] 王建林, 温学发, 赵风华, 等. CO2浓度倍增对8种作物叶片光合作用、蒸腾作用和水分利用效率的影响[J]. 植物生态学报, 2012, 36(5): 438–446 WANG J L, WEN X F, ZHAO F H, et al. Effects of doubled CO2concentration on leaf photosynthesis, transpiration and water use efficiency of eight crop species[J]. Chinese Journal of Plant Ecology, 2012, 36(5): 438–446
[17] 孟凡超, 张佳华, 郝翠, 等. CO2浓度升高和不同灌溉量对东北玉米光合特性及产量的影响[J]. 生态学报, 2015, 35(7): 2126–2135 MENG F C, ZHANG J H, HAO C, et al. Effects of elevated CO2and different irrigation on photosynthetic parameters and yield of maize in Northeast China[J]. Acta Ecologica Sinica, 2015, 35(7): 2126–2135
[18] KNAUER J, ZAEHLE S, REICHSTEIN M, et al. The response of ecosystem water-use efficiency to rising atmospheric CO2concentrations: Sensitivity and large-scale biogeochemical implications[J]. New Phytologist, 2017, 213(4): 1654–1666
[19] ROGERS H H, RUNION G B, KRUPA S V. Plant responses to atmospheric CO2enrichment with emphasis on roots and the rhizosphere[J]. Environmental Pollution, 1994, 83(1/2): 155–189
[20] 李伏生, 康绍忠, 张富仓. CO2浓度、氮和水分对春小麦光合、蒸散及水分利用效率的影响[J]. 应用生态学报, 2003, 14(3): 387–393 LI F S, KANG S Z, ZHANG F C. Effects of CO2enrichment, nitrogen and water on photosynthesis, evapotranspiration and water use efficiency of spring wheat[J]. Chinese Journal of Applied Ecology, 2003, 14(3): 387–393
[21] LOADER N J, SWITSUR V R, FIELD E M. High-resolution stable isotope analysis of tree rings: Implications of ‘microdendroclimatology’ for palaeoenvironmental research[J]. The Holocene, 1995, 5(4): 457–460
[22] NICOTRA A B, COSGROVE M J, COWLING A, et al. Leaf shape linked to photosynthetic rates and temperature optima in South Africanspecies[J]. Oecologia, 2008, 154(4): 625–635
[23] 韩凡香, 常磊, 柴守玺, 等. 半干旱雨养区秸秆带状覆盖种植对土壤水分及马铃薯产量的影响[J]. 中国生态农业学报, 2016, 24(7): 874–882 HAN F X, CHANG L, CHAI S X, et al. Effect of straw strip covering on ridges on soil water content and potato yield under rain-fed semiarid conditions[J]. Chinese Journal of Eco-Agriculture, 2016, 24(7): 874–882
[24] 靳新红, 王百田, 郭红艳, 等. 黄土半干旱区枣、榆水分利用效率的比较研究[J]. 中国生态农业学报, 2009, 17(1): 90–93 JIN X H, WANG B T, GUO H Y, et al. Comparison of water use efficiency ofandin semi-arid zones of the Loess Plateau[J]. Chinese Journal of Eco-Agriculture, 2009, 17(1): 90–93
[25] GULÍAS J, SEDDAIU G, CIFRE J, et al. Leaf and plant water use efficiency in cocksfoot and tall fescue accessions under differing soil water availability[J]. Crop Science, 2012, 52(5): 2321–2331
[26] PONTON S, FLANAGAN L B, ALSTAD K P, et al. Comparison of ecosystem water-use efficiency among Douglas-fir forest, aspen forest and grassland using eddy covariance and carbon isotope techniques[J]. Global Change Biology, 2006, 12(2): 294–310
[27] GREAVES G E, WANG Y M. Yield response, water productivity, and seasonal water production functions for maize under deficit irrigation water management in southern Taiwan[J]. Plant Production Science, 2017, 20(4): 353–365
[28] 胡中民, 于贵瑞, 王秋凤, 等. 生态系统水分利用效率研究进展[J]. 生态学报, 2009, 29(3): 1498–1507 HU Z M, YU G R, WANG Q F, et al. Ecosystem level water use efficiency: A review[J]. Acta Ecologica Sinica, 2009, 29(3): 1498–1507
[29] 刘莹, 李鹏, 沈冰, 等. 采用稳定碳同位素法分析白羊草在不同干旱胁迫下的水分利用效率[J]. 生态学报, 2017, 37(9): 3055–3064 LIU Y, LI P, SHEN B, et al. Effects of drought stress onwater-use efficiency based on stable carbon isotope[J]. Acta Ecologica Sinica, 2017, 37(9): 3055–3064
[30] TANG B, YIN C Y, YANG H, et al. The coupling effects of water deficit and nitrogen supply on photosynthesis, WUE, and stable isotope composition in[J]. Acta Physiologiae Plantarum, 2017, 39: 148
[31] FARQUHAR G D, BUCKLEY T N, MILLER J M. Optimal stomatal control in relation to leaf area and nitrogen content[J]. Silva Fennica, 2002, 36(3): 625–637
[32] HETHERINGTON A M, WOODWARD F I. The role of stomata in sensing and driving environmental change[J]. Nature, 2003, 424(6951): 901–908
[33] 张喜英. 提高农田水分利用效率的调控机制[J]. 中国生态农业学报, 2013, 21(1): 80–87 ZHANG X Y. Regulating mechanisms for improving farmland water use efficiency[J]. Chinese Journal of Eco-Agriculture, 2013, 21(1): 80–87
[34] LIANG Y P, GAO Y, WANG G S, et al. Luxury transpiration of winter wheat and its responses to deficit irrigation in North China Plain[J]. Plant Soil and Environment, 2018, 64(8): 361–366
[35] LAVOIE-LAMOUREUX A, SACCO D, RISSE P A, et al. Factors influencing stomatal conductance in response to water availability in grapevine: A meta-analysis[J]. Physiologia Plantarum, 2017, 159(4): 468–482
[36] RAKOCEVIC M, MATSUNAGA F T, MÜLLER M, et al. Stress-induced DREB1A gene changes heliotropism and reduces drought stress in soybean plants under greenhouse conditions[C]//Proceedings of 2016 IEEE International Conference on Functional-Structural Plant Growth Modeling, Simulation, Visualization and Applications. Qingdao, China: IEEE, 2016: 183–188
[37] MEI X R, ZHONG X L, VINCENT V, et al. Improving water use efficiency of wheat crop varieties in the North China Plain: Review and analysis[J]. Journal of Integrative Agriculture, 2013, 12(7): 1243–1250
[38] DEVI M J, SINCLAIR T R, VADEZ V. Genotypic variation in peanut for transpiration response to vapor pressure deficit[J]. Crop Science, 2010, 50(1): 191–196
[39] KHAZAEI H, MONNEVEUX P, SHAO H B, et al. Variation for stomatal characteristics and water use efficiency among diploid, tetraploid and hexaploid Iranian wheat landraces[J]. Genetic Resources and Crop Evolution, 2010, 57(2): 307–314
[40] BALL J T. An analysis of stomatal conductance[D]. California: Stanford University, 1988: 35–59
[41] YU Q, WANG T D. Simulation of the physiological responses of C3plant leaves to environmental factors by a model which combines stomatal conductance, photosynthesis and transpiration[J]. Acta Botanica Sinica, 1998, 40(8): 740–754
[42] YU G R, ZHUANG J, YU Z L. An attempt to establish a synthetic model of photosynthesis-transpiration based on stomatal behavior for maize and soybean plants grown in field[J]. Journal of Plant Physiology, 2001, 158(7): 861–874
[43] ZHANG B Z, LIU Y, XU D, et al. Evapotranspiraton estimation based on scaling up from leaf stomatal conductance to canopy conductance[J]. Agricultural and Forest Meteorology, 2011, 151(8): 1086–1095
[44] WALDEN-COLEMAN A E, RAJCAN I, EARL H J. Dark-adapted leaf conductance, but not minimum leaf conductance, predicts water use efficiency of soybean (L. Merr.)[J]. Canadian Journal of Plant Science, 2013, 93(1): 13–22
[45] DING R S, KANG S Z, DU T S, et al. Scaling up stomatal conductance from leaf to canopy using a dual-leaf model for estimating crop evapotranspiration[J]. PLoS One, 2014, 9(4): e95584
[46] MEDLYN B E, DE KAUWE M G, LIN Y S, et al. How do leaf and ecosystem measures of water-use efficiency compare?[J]. New Phytologist, 2017, 216(4): 758–770
[47] ALBASHA R, OURNIER C, PRADAL C, et al. HydroShoot: a new FSPM model for simulating hydraulic structure and gas-exchange dynamics of complex plants canopies under water deficit[C]//IEEE International Conference on Functional-Structural Plant Growth Modeling, Simulation, Visualization and Applications (FSPMA). Qingdao, 2016.
[48] SENBAYRAM M, TRÄNKNER M, DITTERT K, et al. Daytime leaf water use efficiency does not explain the relationship between plant N status and biomass water-use efficiency of tobacco under non-limiting water supply[J]. Journal of Plant Nutrition and Soil Science, 2015, 178(4): 682–692
[49] ASSOULINE S, OR D. Plant water use efficiency over geological time-evolution of leaf stomata configurations affecting plant gas exchange[J]. PLoS One, 2015, 10(4): e0127015
[50] MEDRANO H, TOMÁS M, MARTORELL S, et al. Improving water use efficiency of vineyards in semi-arid regions. A review[J]. Agronomy for Sustainable Development, 2015, 35(2): 499–517
[51] LINDERSON M L, MIKKELSEN T N, IBROM A, et al. Up-scaling of water use efficiency from leaf to canopy as based on leaf gas exchange relationships and the modeled in-canopy light distribution[J]. Agricultural and Forest Meteorology, 2012, 152: 201–211
[52] 黄桂荣, 梅旭荣, 严昌荣, 等. 干旱条件下冬小麦不同尺度水分利用效率及其之间的关系[J]. 麦类作物学报, 2017, 37(4): 528–534 HUANG G R, MEI X R, YAN C R, et al. Water use efficiency of winter wheat near isogenic lines at leaf plant and population levels and their relationship under drought condition[J]. Journal of Triticeae Crops, 2017, 37(4): 528–534
[53] CONDON A G, RICHARDS R A, REBETZKE G J, et al. Breeding for high water-use efficiency[J]. Journal of Experimental Botany, 2004, 55(407): 2447–2460
[54] FARQUHAR G. 水分利用效率与用水有效性: 基于气孔视角的稳定同位素应用研究[J]. 张玉翠, 译. 中国生态农业学报, 2014, 22(8): 886–889 FARQUHAR G. Water-use efficiency and water use effectiveness: A stomatal perspective using stable isotopes[J]. ZHANG Y C, trans. Chinese Journal of Eco-Agriculture, 2014, 22(8): 886–889
[55] EHDAIE B, HALL A E, FARQUHAR G D, et al. Water-use efficiency and carbon isotope discrimination in wheat[J]. Crop Science, 1991, 31(5): 1282–1288
[56] 谭巍, 陈洪松, 王克林, 等. 桂西北喀斯特坡地不同演替阶段典型植物碳同位素组成差异[J]. 中国生态农业学报, 2010, 18(6): 1223–1227 TAN W, CHEN H S, WANG K L, et al. Leaf δ13C of plants in different vegetation succession stages on karst hillslope of Northwest Guangxi, China[J]. Chinese Journal of Eco-Agriculture, 2010, 18(6): 1223–1227
[57] LLOYD J, FARQUHAR G D.13C discrimination during CO2assimilation by the terrestrial biosphere[J]. Oecologia, 1994, 99(3/4): 201–215
[58] YU Q, LIU J D, LUO Y. Applicability of some stomatal models to natural conditions[J]. Acta Botanica Sinica, 2000, 42(2): 203–206
[59] 任传友, 于贵瑞, 王秋凤, 等. 冠层尺度的生态系统光合-蒸腾耦合模型研究[J]. 中国科学 D辑: 地球科学, 2004, 34(S2): 141–151REN C Y, YU G R, WANG Q F, et al. Photosynthesis-transpiration coupling model at canopy scale in terrestrial ecosystem[J]. Science in China Series D: Earth Sciences, 2004, 34(S2): 141-151
[60] 张宝忠, 刘钰, 许迪, 等. 夏玉米叶片和冠层尺度的水碳耦合模拟[J]. 科学通报, 2013, 58(12): 1121–1130 ZHANG B Z, LIU Y, XU D, et al. Water-carbon coupling modeling of summer maize at the leaf and canopy scales[J]. Chinese Science Bulletin, 2013, 58(12): 1121-1130
A review of progress in research and scaling-up methods of crop water use efficiency*
JIANG Hanbing1,2, ZHANG Yucui1, REN Xiaodong1,3, YAO Jiawei1,2, SHEN Yanjun1**
(1. Key Laboratory of Agricultural Water Resources, Chinese Academy of Sciences / Hebei Laboratory of Water-Saving Agriculture / Center for Agricultural Resources Research, Institute of Genetics and Developmental Biology, Chinese Academy of Sciences, Shijiazhuang 050022, China; 2. University of Chinese Academy of Sciences, Beijing 100049, China; 3. College of Biological and Geographical Sciences, Qinghai Normal University, Xining 810000, China)
Increasing crop water use efficiency (WUE) is an effective way of alleviating agricultural water scarcity. The scaling up of water use efficiency is the basis for mutual representation, verification and application of achievements at various scales. This paper summarized the main observation technologies of leaf-scale, plant-scale and plantation-scale WUE. At present, the widely used methods include leaf gas exchange measurement, carbon isotopic discrimination, pot weighing method, eddy covariance system, etc. Carbon isotope discrimination provides a new idea for the study of long-term cumulative effects of crop water use conditions which is also available at every scale. We reviewed the impacting factors and the related physiological mechanisms of crop water use at multi-scale WUE. Crop WUE at each scale was regulated by stomatal conductance and crops usually regulated stomatal aperture to response to temperature, humidity, CO2and other interactive environmental factors. Stomatal optimization theory essentially sought optimal state of stomata under complex environmental conditions to coordinate the process of photosynthesis and transpiration of crops. Instantaneous WUE at leaf scale cannot directly represent water use status at larger spatial and temporal scales. Thus we also discussed the feasibility of scaling up WUE from leaf to plant to plantation scales and analyzed the main limiting factors at each scale transfer. We pointed out the difficulties in transfer from leaf to plant in terms of WUE. It mainly focused on three points — uncertainty in leaf and light distributions, plant nighttime respiration and transpiration, and plant physiological adjustment mechanisms. Research on leaf to plant to plantation scale transfer was mainly influenced by canopy internal resistance, boundary layer resistance, soil evaporation, night transpiration of crops, crop water use and assimilates partitioning mechanism. Finally, existing research achievements on scale transfer were summarized. At present, WUE scale transfer depended mainly on improvement of models and observation methods. The transfer from leaf to plant focused on separation of water use during day and night and photosynthetic characteristics of each part of the crop. For transfer from leaf to plant to plantation scale, studies explored efficient ways. First, studies understood the structure of evapotranspiration and confirmed the characteristics of water use. Second, studies used the relationship between stomatal and canopy conductance as breakthrough point via models to explore transfer mechanisms. Actually, several models had already been established and applied in this respect.
Water use efficiency; Scaling up; Carbon isotopic discrimination; Stomatal conductance
, E-mail: yjshen@sjziam.ac.cn
May 25, 2018;
Sep. 2, 2018
S181
A
2096-6237(2019)01-0050-10
10.13930/j.cnki.cjea.180501
姜寒冰, 张玉翠, 任晓东, 要家威, 沈彦俊. 作物水分利用效率研究方法及尺度传递研究进展[J]. 中国生态农业学报(中英文), 2019, 27(1): 50-59
JIANG H B, ZHANG Y C, REN X D, YAO J W, SHEN Y J. A review of progress in research and scaling-up methods of crop water use efficiency[J]. Chinese Journal of Eco-Agriculture, 2019, 27(1): 50-59
* 国家重点研发计划课题(2016YFC0401403)、国家自然科学基金面上项目(31870422)、河北省自然科学基金项目(D2016503001)和中国科学院青年创新促进会项目(2017138)资助
沈彦俊, 主要从事生态水文过程的研究。E-mail: yjshen@sjziam.ac.cn
姜寒冰, 主要从事作物水分利用效率和农田水平衡过程研究。E-mail: jianghanbing16@163.com
2018-05-25
2018-09-02
* This study was supported by the National Key Research and Development Plan of China (2016YFC0401403), the National Natural Science Foundation of China (31870422), the Natural Science Foundation of Hebei Province (D2016503001), and the Youth Innovation Promotion Association of Chinese Academy of Sciences (2017138).
猜你喜欢
农业工程学报(2022年8期)2022-08-08
作物学报(2022年10期)2022-07-21
作物杂志(2022年3期)2022-07-06
中国农业信息(2022年1期)2022-05-25
农业工程学报(2022年3期)2022-04-16
河南水利与南水北调(2022年1期)2022-03-16
农业机械学报(2021年11期)2021-12-07
农业工程学报(2021年3期)2021-04-15
干旱地区农业研究(2020年4期)2020-09-22
节水灌溉(2020年9期)2020-09-16
