
MiR-222对山羊副流感病毒3型复制的影响
2019-01-02 07:55:34钟纯燕李基棕李文良刘茂军嵇辛勤杨蕾蕾张纹纹
畜牧兽医学报 2018年12期
钟纯燕,李基棕,毛 立,李文良,郝 飞,孙 敏,刘茂军,主 性,嵇辛勤,肖 芳,杨蕾蕾,张纹纹
(1. 江苏省农业科学院 兽医研究所,农业部兽用生物制品工程技术重点实验室,南京210014;2. 临沂大学 药学院,临沂 276000; 3. 贵州大学 动物科学学院,贵阳 550025)
山羊副流感病毒3型(caprine parainfluenza virus type 3, CPIV3)属于副黏病毒科呼吸道病毒属的新成员,为有囊膜的单股负链RNA病毒。同属成员还包括人副流感病毒1型和3型(human PIV1、human PIV3)、仙台病毒(Sendai virus, SV)和牛副流感病毒3型(bovine PIV3)[1]。2013年起在江苏、安徽等地的山羊养殖场中检测到CPIV3,感染羊出现不同程度的呼吸道疾病,RT-PCR扩增测序其N、M、F和HN基因发现,这些基因与HPIV1和HPIV3的相似性仅为76.9%~83.5%[2-4]。随后本实验室将分离鉴定的CPIV3 JS2013株进行山羊致病性试验,结果得出,CPIV3感染的山羊出现咳嗽、流鼻涕、发热和精神沉郁等临床症状,并出现持续性的病毒血症和排毒,剖检可见肺和气管出现较严重的病理损伤,并且CPIV3能通过气溶胶水平传播,使相邻圈舍健康羊出现呼吸道症状,并可检出病毒血症和排毒[5],这给养羊业健康发展带来了新的威胁。本实验室应用建立的荧光定量RT-qPCR方法[6]和阻断ELISA方法[7]对采集的鼻拭子和血清进行流行病学调查发现,CPIV3病原核酸检出率达44.7%,抗体检出率达39.3%。因此,该病原对养羊业的危害应该引起足够的重视。
miRNA(microRNA)是一类长度为19~24 nt的非编码RNA,细胞内源性miRNA与RNA诱导的沉默复合物(RNA-induced silencing complex, RISC)结合,通过互补结合靶mRNA的5′非编码区(5′ untranslated regions, 5′UTR)、3′UTR或编码区碱基,可降解mRNA或抑制mRNA翻译成蛋白质,从而参与细胞分化、增殖、代谢及凋亡的各个环节,与动物机体生殖发育、肿瘤形成及病毒感染过程密切相关[8-9]。近期研究发现,miRNA广泛参与宿主体内的抗病毒作用,miR-122可与丙型肝炎病毒(hepatitis C virus, HCV)基因组的5′UTR互补结合来调控病毒增殖,因此miR-122在肝细胞中的表达能有效降低HCV复制水平[10-11]。miR-130在体内外具有抗猪繁殖与呼吸综合征病毒(porcine reproductive and respiratory syndrome virus, PRRSV)感染作用,双荧光素酶报告系统揭示PRRSV的5′UTR是miR-130调控的靶位点[12]。金属蛋白酶m41 ftsh是西尼罗河病毒(West Nile virus, WNV)复制的必需因子,aae-miR-2940-5p能增强该蛋白酶的基因转录,提高病毒的复制效率[13]。
在前期研究中,本实验室使用CPIV3感染MDBK细胞,通过高通量测序技术筛选出感染细胞和正常细胞中差异表达的miRNA,结果发现miR-222在CPIV3感染细胞样品中出现显著下调、GO注释和KEGG富集,miR-222调控的靶基因(interferon regulatory factor 2, IRF2)参与天然免疫通路,且IRF2是IFN信号通路的负调控因子[14]。因此过表达的miR-222可能会促进IFN的表达,抑制病毒增殖。为挖掘具有抗CPIV3增殖作用的miRNA,本研究将miR-222在细胞内进行过表达和抑制表达,探讨miR-222对CPIV3复制的影响及其机制,以期为CPIV3的防控提供重要的理论依据。
1 材料与方法
1.1 病毒与主要试剂
CPIV3 JSHA2014-1 MDBK细胞适应株为本实验室分离并保存;MDBK细胞购自中国兽医药品监察所;293FT细胞由本实验室保存;One Step PrimeScriptTMRT-PCR Kit购自TaKaRa公司;EasyScript One-Step gDNA Removal and cDNA Synthesis SuperMix、EasyScript One-Step RT-PCR SuperMix及其他PCR试剂购自北京全氏金生物公司;胶回收试剂盒和质粒提取试剂盒购自Axygen公司;IRF2抗体购自Abcam公司;Lipofectamine 2000转染试剂购自Invitrogen公司;双荧光素酶报告基因载体pRL-CMV、pGL3-control和Dual Luciferase Reporter Gene Assay Kit均购自Promega公司。
1.2 miR-222 mimics和miR-222 inhibitor的设计与合成
MiR-222 mimics和miR-222 inhibitor是根据miR-222的序列(AGCUACAUCUGGCUACU-GGGU)设计,并通过化学合成方法制备。MiR-222 mimics能显著提高成熟miR-222的表达;miR-222 inhibitor能特异性与成熟miR-222靶向结合从而抑制其发挥作用。两种miRNA均由上海吉玛制药技术有限公司合成。
1.3 MiR-222 对CPIV3复制的影响
在24孔板中准备MDBK单层细胞,待细胞长至60%汇合度时,分别将miR-222 mimics和miR-222 inhibitor转染MDBK细胞,转染方法按照Lipofectamine 2000说明书进行。转染后24 h,每孔接种1 000 TCID50的CPIV3 JSHA2014-1株,1.5 h后弃去病毒液,更换成500 μL 2% DMEM培养基,置37 ℃继续培养,分别在接毒后24和48 h收获细胞上清,用RT-qPCR和TCID50法测定CPIV3病毒含量。同时设立转染NC对照组。
1.4 RT-qPCR和TCID50检测CPIV3滴度
用Transzol UP试剂提取上清中的总RNA后,使用TaKaRa公司的One Step PrimeScriptTMRT-PCR Kit检测上清中CPIV3核酸含量。使用ABI Step One荧光定量PCR仪进行扩增,RT-qPCR的反应体系和反应条件参照试剂盒说明书进行。所用的引物qCPIV3F、qCPIV3R和探针qCPIV3-probe序列见表1。
表1相关引物序列表
Table1Thesequenceofprimersusedinthisstudy
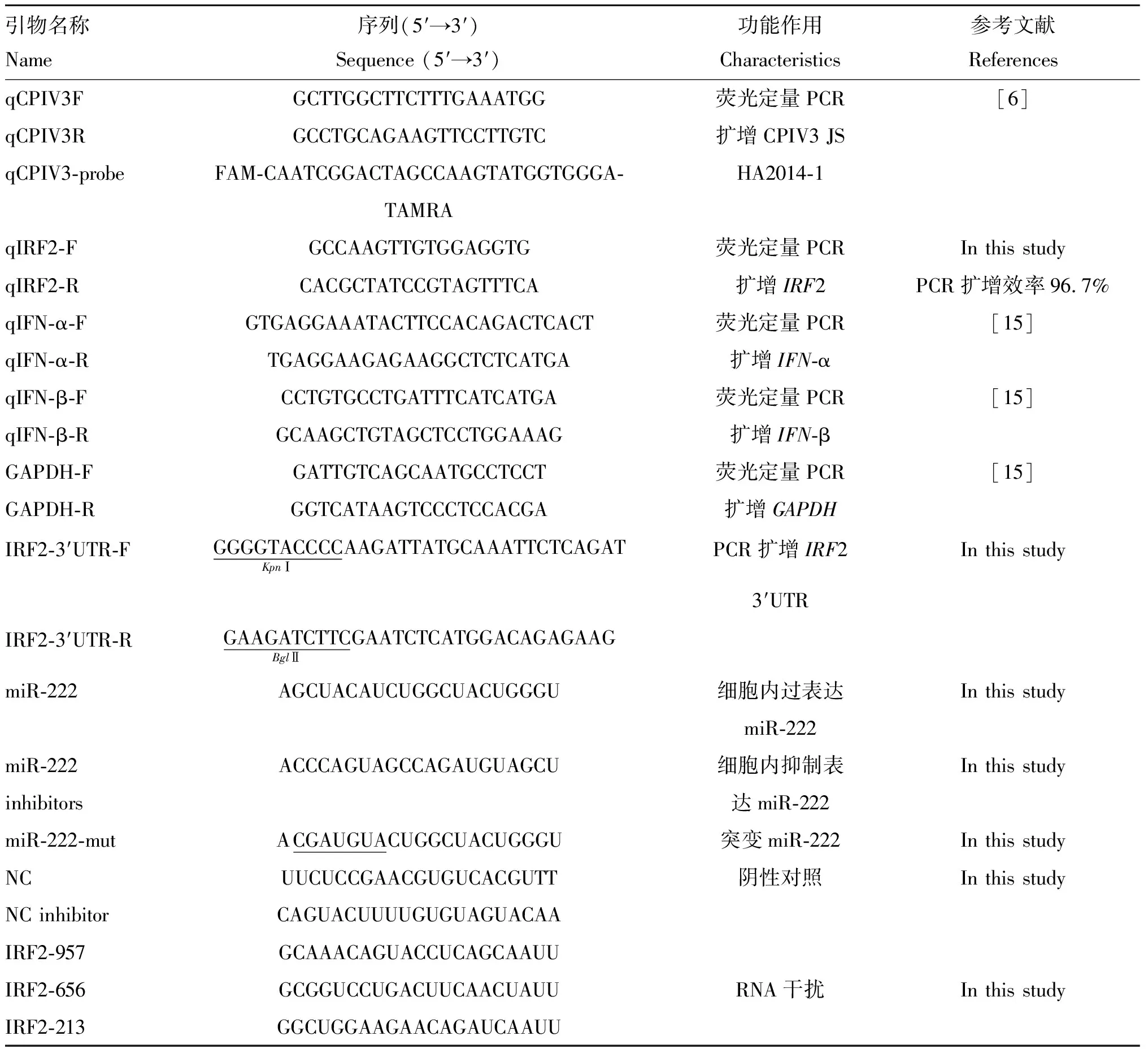
引物名称Name序列(5′→3′)Sequence (5′→3′)功能作用Characteristics参考文献ReferencesqCPIV3FGCTTGGCTTCTTTGAAATGG荧光定量PCR[6]qCPIV3RGCCTGCAGAAGTTCCTTGTC扩增CPIV3 JSqCPIV3-probeFAM-CAATCGGACTAGCCAAGTATGGTGGGA- HA2014-1TAMRAqIRF2-FGCCAAGTTGTGGAGGTG荧光定量PCRIn this study qIRF2-RCACGCTATCCGTAGTTTCA扩增IRF2PCR扩增效率96.7%qIFN-α-FGTGAGGAAATACTTCCACAGACTCACT荧光定量PCR[15]qIFN-α-RTGAGGAAGAGAAGGCTCTCATGA扩增IFN-αqIFN-β-FCCTGTGCCTGATTTCATCATGA荧光定量PCR[15]qIFN-β-RGCAAGCTGTAGCTCCTGGAAAG扩增IFN-βGAPDH-FGATTGTCAGCAATGCCTCCT荧光定量PCR[15]GAPDH-RGGTCATAAGTCCCTCCACGA扩增GAPDHIRF2-3′UTR-FGGGGTACCCCKpnⅠAAGATTATGCAAATTCTCAGATPCR扩增IRF2In this study 3′UTR IRF2-3′UTR-RGAAGATCTTCBglⅡGAATCTCATGGACAGAGAAGmiR-222 AGCUACAUCUGGCUACUGGGU细胞内过表达In this study miR-222miR-222ACCCAGUAGCCAGAUGUAGCU细胞内抑制表In this studyinhibitors达miR-222miR-222-mutACGAUGUACUGGCUACUGGGU突变miR-222In this studyNCUUCUCCGAACGUGUCACGUTT阴性对照In this studyNC inhibitorCAGUACUUUUGUGUAGUACAAIRF2-957GCAAACAGUACCUCAGCAAUUIRF2-656GCGGUCCUGACUUCAACUAUURNA干扰In this studyIRF2-213GGCUGGAAGAACAGAUCAAUU
将MDBK细胞接种96孔板,待细胞长至90%汇合度,用维持液将收获的病毒液进行10倍倍比稀释,然后接种细胞,每孔100 μL,重复4个孔,37 ℃培养3 d,然后观察每个稀释度出现细胞病变的孔数。按Reed-Muench方法计算病毒TCID50。
1.5 miR-222调控CPIV3复制的分子机制
1.5.1 miR-222对IRF2和IFN产生的影响 在24孔板中准备MDBK单层细胞,待细胞长至60%汇合度时,将miR-222 mimics转染MDBK细胞,转染方法按照Lipofectamine 2000说明书进行。转染24 h后收获细胞,用RT-qPCR检测细胞中IFN的mRNA水平。同时设立转染NC对照组。
提取收获细胞的总RNA,使用北京全氏金生物公司的EasyScript One-Step gDNA Removal and cDNA Synthesis SuperMix和EasyScript One-Step RT-PCR SuperMix进行反转录和RT-qPCR检测。使用ABI Step One荧光定量PCR仪进行扩增,反应体系和反应条件均参照试剂盒说明书设置。以GAPDH为内参计算转染miRNA组和转染NC对照组的IFN转录量变化,结果以-ΔΔCt表示。所用的引物qIFN-α-F、qIFN-α-R,qIFN-β-F、qIFN-β-R和GAPDH-F、GAPDH-R见表1。
1.5.2 miR-222靶基因预测 用生物信息学软件TargetScan预测miR-222的靶基因及其结合位点,并选择与干扰素信号通路相关的靶基因进行功能验证。
1.5.3 双荧光素酶报告基因系统分析miR-222靶基因 根据IRF2 mRNA的3′UTR设计引物,见表1。进行RT-PCR扩增,将扩增产物通过KpnⅠ和BglⅡ酶切位点克隆于荧光素酶报告载体pGL3-control中,构建重组荧光素酶报告载体pGL3-IRF2 3′UTR。将pGL3-IRF2 3′UTR、pRL-CMV和miR-222 mimics共转染293FT细胞,利用双荧光素酶报告基因检测试剂盒分析荧光素酶的变化。同时设立转染NC对照组。
1.5.4 Western blot分析 在24孔板中准备MDBK单层细胞,待细胞长至60%汇合度时,将miR-222 mimics转染MDBK细胞,转染方法按照Lipofectamine 2000说明书进行。转染24 h后收获细胞,用Western blot分析IRF2蛋白的表达水平。同时设立转染NC对照组。
1.5.5IRF2基因敲除对CPIV3复制的影响 针对IRF2的保守区域设计并合成3条siRNA(si-IRF2),同时设计并合成siRNA阴性对照,序列见表1。siRNA由上海吉玛制药技术有限公司合成。
在24孔板中准备MDBK单层细胞,待细胞长至60%汇合度时,将3个siIRF2同时转染MDBK细胞,转染方法按照Lipofectamine 2000说明书进行。转染后24 h,每孔接种1 000 TCID50的CPIV3 JSHA2014-1株,1.5 h后弃去病毒液,更换成500 μL 2% DMEM培养基,置于37 ℃继续培养,分别在接毒后24和48 h收获细胞上清,用RT-qPCR和TCID50法测定CPIV3病毒含量。同时设立转染siRNA阴性对照组。
1.6 统计分析
2 结 果
2.1 MiR-222 对CPIV3复制的影响
将合成的miR-222 mimics和miR-222 inhibitor,转染导入MDBK细胞内,通过RT-qPCR技术和TCID50方法来评价CPIV3的复制能力,发现转染miR-222 mimics在24和48 h均能有效抑制CPIV3的增殖(P<0.01),如图1所示;转染miR-222 inhibitor在24和48 h均能有效促进CPIV3的增殖(P<0.01),如图2所示。
2.2 MiR-222对Ⅰ型干扰素转录的影响
使用miR-222 mimics转染MDBK细胞,24 h后收集MDBK细胞,RT-qPCR检测细胞中的IFN-α和IFN-β转录水平。结果发现,IFN-α(P<0.01)和IFN-β(P<0.001)转录均极显著上调,如图3所示。
2.3 MiR-222的靶基因预测及对靶基因的调控作用
用TargetScan预测miR-222的靶基因及其结合位点发现,IRF2基因的3′UTR中存在miR-222的潜在靶序列(3′UTR:977—984 bp),如图4所示。用双荧光素酶报告基因检测试剂盒分析荧光素酶变化,结果显示转染miR-222 mimics组的荧光素酶活性与对照组相比降低55%左右,差异极显著(P<0.01),但转染miR-222-mut mimics组的荧光素酶活性与对照组相比无显著差异(P=1),见图5A。用RT-qPCR检测IRF2 mRNA转录水平,结果显示,转染miR-222 mimics组IRF2 mRNA量显著低于对照组(P<0.05),见图5B。用Western blot分析IRF2蛋白的表达水平,结果显示,转染miR-222 mimics组IRF2蛋白表达量显著低于对照组,见图5C。

图1 MiR-222 mimics对CPIV3增殖的抑制作用Fig.1 Bta-miR-222 mimics inhibited CPIV3 replication in MDBK cells

图2 MiR-222 inhibitor对CPIV3增殖的促进作用Fig.2 Bta-miR-222 inhibitor promoted CPIV3 replication in MDBK cells
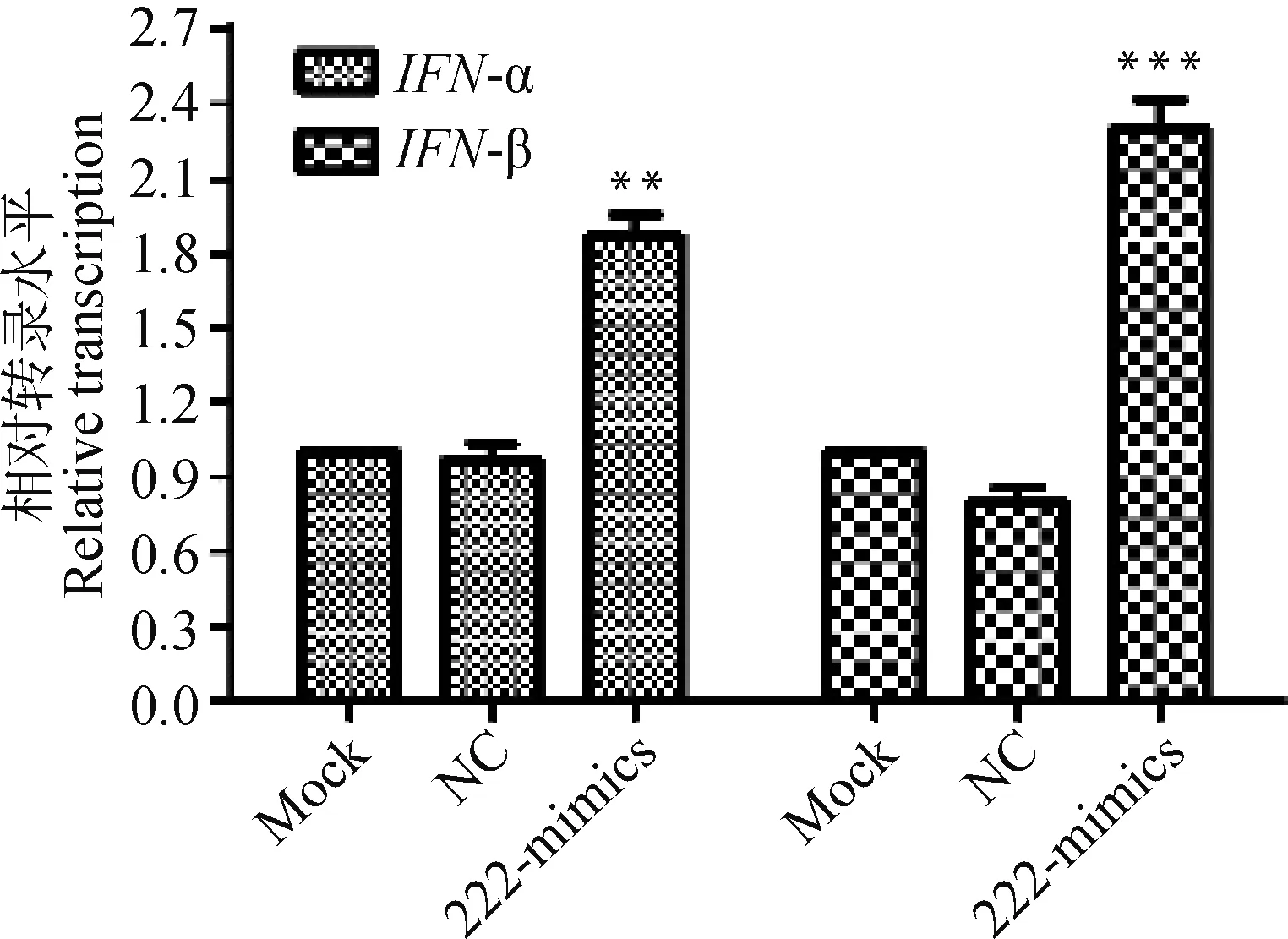
图3 IFN-α和IFN-β的转录变化Fig.3 Bta-miR-222 mimics increase typeⅠinterferon transcription in MDBK cells
2.4 IRF2基因敲除对CPIV3复制的影响
将3条si-IRF2同时转染于MDBK细胞,24 h后收获细胞,RT-qPCR检测IRF2基因转录水平,结果发现,转染si-IRF2后IRF2基因表达水平显著降低(P<0.05),见图6A。Western blot检测IRF2蛋白的表达水平,结果显示,转染si-IRF2后IRF2蛋白的表达水平显著降低,见图6B。同时在转染si-IRF2 24 h后接种CPIV3病毒液,24 h后收获病毒液,RT-qPCR技术和TCID50检测病毒含量,结果显示,转染si-IRF2组其CPIV3病毒含量极显著低于NC对照组(P<0.001),见图6C、D。
3 讨 论
MiR-222家族分布广泛且高度保守,其序列在脊椎动物中具有极高的同源性。近期学者们发现,miR-222在不同的宿主细胞中通过作用于不同的靶基因而发挥调控功能。Zeng等[16]、Dai等[17]和Galardi等[18]的研究证实miR-222能抑制肿瘤细胞的凋亡,其靶基因包括细胞凋亡诱导剂(BCL-2 modifying factor, BMF)基因、蛋白磷酸酶2A和肿瘤抑制因子p27等;Yan等[19]发现miR-222参与先天性免疫应答,调控的靶基因包括肿瘤坏死因子受体相关因子6和白介素1β。Cheng等[20]在稳定表达戊型肝炎病毒ORF3的细胞系中发现,miR-222表达量出现显著下调,ORF3蛋白是启动病毒增殖的调节因子,但该团队没有进一步探究miR-222是否参与机体免疫防御。因此,miR-222调节病毒增殖作用尚无人报道。

图4 MiR-222的靶基因预测Fig.4 Bta-miR-222 directly targets the 3′UTR of IRF2
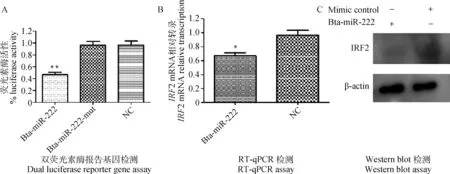
图5 MiR-222对IRF2的调控作用Fig.5 Bta-miR-222 downregulates IRF2 expression in MDBK cell
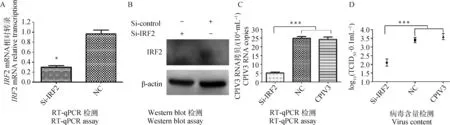
图6 转染si-IRF2对CPIV3复制的影响Fig.6 Bta-miR-222 suppress CPIV3 replication through targeting IRF2
基于此,为了证实MDBK细胞内源性miR-222抗CPIV3免疫作用,本研究通过合成miR-222类似物和抑制剂,分别转染导入细胞内,24 h后收获细胞,以5S rRNA为内参,相对荧光定量RT-PCR检测细胞内miR-222的表达量,得出细胞内miR-222含量分别提高或降低2~3倍。接种CPIV3病毒液,24 h后收毒,荧光定量RT-PCR和TCID50的方法来评价CPIV3的复制能力,发现转染miR-222类似物在24和48 h均能有效抑制CPIV3的增殖;同时,转染相同剂量的miR-222抑制剂在24和48 h均能有效促进CPIV3的增殖。结果说明,miR-222具有调控CPIV3复制的能力。随后在细胞中转染miR-222,荧光定量RT-PCR显示,IFN-α和IFN-β转录水平出现不同程度的上调。IFN能诱导细胞产生多种抗病毒蛋白的细胞因子,具有抑制病毒增殖和调节机体免疫反应的生物学功能,通过检测细胞内IFNmRNA转录水平,可以用来评估机体免疫系统活化程度[21]。以上结果说明,miR-222通过诱导Ⅰ型干扰素的产生,抑制CPIV3增殖。
为了进一步揭示miR-222调控Ⅰ型干扰素影响CPIV3复制的分子机制,本研究用TargetScan预测miR-222的靶基因及其潜在的结合位点发现,IRF2基因的3′UTR中存在miR-222的潜在靶序列,随后用双荧光素酶报告系统、RT-qPCR、Western blot和RNAi等方法证实了miR-222对IRF2的靶向调节作用。IRF2是IFN途径中发挥抗病毒作用的重要节点蛋白,其作用是负向调节IFN产生和信号传导[22]。而细胞因子的稳定产生在维持机体免疫反应平衡中起关键作用,Hida等[23]揭示了IRF2缺陷型小鼠体内IFN产生失调,从而出现渐进性皮炎症状。Li等[14]证实通过将辛德比斯病毒(Sindbis virus)脑内注射IRF2缺陷型小鼠后发现,与正常组小鼠相比,IRF2缺陷型小鼠IFN调节功能紊乱,更利于病毒在体内复制,造成严重的组织损伤和小鼠死亡。在本研究中,miR-222通过抑制IRF2 mRNA转录和蛋白表达,从而促进IFN的产生,提高细胞天然免疫功能,抑制CPIV3在MDBK细胞中增殖。
综上所述,本研究揭示了miR-222影响CPIV3增殖的分子调控机制,发掘了miR-222调控的Ⅰ型干扰素分子通路,这是研究CPIV3致病机制新的切入点,也为抗CPIV3分子药物的研发提供科学基础。
4 结 论
miR-222通过靶向降解IRF2 mRNA提高IFN表达水平来抑制CPIV3复制。
猜你喜欢
天津医科大学学报(2021年4期)2021-08-21 02:14:50
中日友好医院学报(2021年1期)2021-04-14 01:58:32
世界科学技术-中医药现代化(2020年2期)2020-07-25 02:06:06
山东医药(2020年9期)2020-05-20 01:12:16
学苑创造·A版(2020年12期)2020-01-07 14:07:23
中国外汇(2019年15期)2019-10-14 01:00:34
中成药(2018年12期)2018-12-29 12:25:44
中成药(2017年6期)2017-06-13 07:30:35
作文教学研究(2016年1期)2016-07-05 12:22:47
医学研究杂志(2015年8期)2015-06-22 14:00:57
