
马铃薯与蚕豆、荞麦间作对土壤的影响
2018-12-06 01:49刘亚军何文寿
江苏农业科学 2018年21期
刘亚军, 李 越, 马 琨, 何文寿
(1.宁夏大学西北土地退化与生态恢复省部共建国家重点实验室培育基地,宁夏银川 750021; 2.宁夏大学农学院,宁夏银川 750021;
3.商丘市农林科学院,河南商丘 476000)
马铃薯是西部地区的重要农作物,栽培历史悠久,具有稳定的、较高的经济效益[1]。近年来,马铃薯主粮化促进了宁夏南部山区马铃薯产业化发展,使得其栽培面积不断扩大,致使马铃薯轮作倒茬困难。马铃薯多年连续不断地种植导致马铃薯产量下降、土壤质量下降、病虫害频繁不断地发生,而农作物间作是解决这一问题的关键。间作是一种集约化生产模式,在我国具有悠久的栽培历史[2]。合理的间作模式不仅能够利用作物对养分需求特性的不同而促进养分的有效吸收,也能够通过其时间、空间等方面的互补性高效利用水肥气热等资源[3]。已有研究指出,土壤酶活性和微生物群落结构组成影响着土壤养分的吸收和转化,土壤酶活性的降低与微生物结构的失衡是导致土壤质量下降、作物减产的主要原因[4]。而作物合理的间作不仅能够提高土壤中的养分含量、改善土壤的微环境,还能够增加作物的产量,改善作物的品质,提高作物的安全性[5]。傅佳等在重茬种植西洋参对土壤养分和微生物的研究中发现,土壤酶活性与根际微生物类群呈显著正相关,可改善土壤中养分的转化和吸收、根际微生物群落结构的组成和数量[6]。汪春明等在马铃薯间作蚕豆的研究中认为,间作模式有利于改善马铃薯连作栽培的根际微生态环境[7]。吴红英等在沙地梨园间作研究中指出,间作可以增加梨树各时期和各土层中土壤微生物群落的数量,改变土壤微生物群落中细菌、放线菌和真菌的比例[8]。刘均霞等在对玉米间作大豆的体系研究认为,间作体系中玉米、大豆的土壤酶活性和根际土壤微生物数量显著高于相应的单作对照组,间作大豆处理的脲酶活性和根际土壤细菌数量显著高于单作大豆对照组[9]。Li等在间作玉米和蚕豆的研究中采用测定磷脂脂肪酸(PLFA)含量的方法,证实间作模式对玉米和豆科作物根际微生物群落的结构和功能有影响[10]。可见,合理的间作模式可以改善土壤酶活性和微生物结构组成,提升土壤质量,对作物的生长发育有着重要影响。本研究以单作马铃薯、蚕豆、荞麦,马铃薯间作蚕豆、荞麦为处理,通过研究马铃薯不同间作模式对土壤养分含量、土壤酶活性和土壤微生物群落的影响,揭示马铃薯不同间作模式下酶活性和微生物群落结构变化的特点,旨在为马铃薯合理间作提供科学依据。
1 材料与方法
1.1 试验地点概况
该试验于2015年4—10月在宁夏农林科学院固原头营科研基地徐河村进行,地处东经106°44′、北纬36°10′,海拨 2 341 m。初霜期9月上、中旬,终霜期5月上、中旬,无霜期140 d左右,降水291~415 mm,属半干旱半湿润地区。供试土壤为黄绵土,土壤基础理化性状为:有机质含量 10.63 g/kg,全氮含量0.258 g/kg,全磷含量0.645 g/kg,碱解氮含量20.51 mg/kg,速效磷含量45.5 mg/kg,速效钾含量170.5 mg/kg,土壤pH值9.00。
1.2 试验设计与样品采集
试验采用单因素区组设计,种植方式分为:处理1,马铃薯单作(MP);处理2,蚕豆单作(MF);处理3,荞麦单作(MW);处理4,马铃薯+蚕豆(P+F);处理5,马铃薯+荞麦(P+W)。小区面积为6 m×4 m,4次重复。供试马铃薯为青薯9号,蚕豆(faba bean)为青蚕11号,荞麦(buckwhea)为宁荞1号。种植2行马铃薯,间种2行蚕豆或荞麦,马铃薯株行距为40 cm×60 cm,荞麦行距为30 cm,蚕豆株行距为 20 cm×20 cm。马铃薯与蚕豆间距30 cm,马铃薯与荞麦间距35 cm。
试验地前茬作物为玉米,秋翻,返春后耙耱整地,播种前施用优质农家肥30 000 kg/hm2、尿素255 kg/hm2,磷酸二铵170 kg/hm2,磷酸二氢钾90 kg/hm2,其中70%氮肥作基肥,30%氮肥作追肥。4月25日播种蚕豆、马铃薯,5月23日播种荞麦,8月12日收获蚕豆,9月20日收获荞麦、马铃薯。
在马铃薯花期,选择毗邻马铃薯的1行,通过土钻利用五点取样法随机取五处土壤,把取到的土样混合后放入冰盒中带回实验室。土壤样品一部分存于4 ℃冰箱,用于土壤微生物群落结构与生物量的测定;一部分自然风干,用于土壤理化性质和酶活性的分析。
1.3 测定指标及方法
1.3.1 土壤理化性状 土壤有机质含量采用重铬酸钾容量法-外加热法;全氮含量采用半微量凯氏法;碱解氮含量采用碱解扩散法;全磷含量采用HClO4-H2SO4法;速效磷含量采用0.5 mol/L NaHCO3法;速效钾含量采用NH4OAc浸提火焰光度法;土壤pH值(1 ∶5土水比)的测定参照文献[11]。
1.3.2 土壤酶活性测定方法 土壤脲酶活性采用苯酚钠-次氯酸钠比色法测定;土壤碱性磷酸酶活性采用磷酸苯二钠比色法测定;土壤过氧化氢酶活性采用高锰酸钾滴定法测定[12]。
1.3.3 土壤微生物群落的测定 土壤微生物群落结构组成采用稀释平板法测定。其中,细菌培养采用牛肉膏蛋白胨琼脂培养基;真菌培养采用马铃薯-蔗糖琼脂(PDA)培养基;放线菌培养采用改良高氏一号培养基[13]。
以土壤微生物磷脂脂肪酸(PLFA)含量反映土壤微生物生物量,其测定步骤为:称取相当于4 g干土质量的新鲜土样于棕色培养瓶中,加入磷酸缓冲液 30 mL 和甲醇 ∶三氯甲烷(2 ∶1)溶液105 mL后振荡2 h,取出分别加入三氯甲烷和无菌超纯水各36 mL,振荡混匀,在黑暗中培养18~24 h;吸掉试剂瓶上层的水和甲醇,用无水硫酸钠除去未吸净的水,然后进行过滤、旋转蒸发浓缩。其中利用固相萃取技术,通过SPE固相萃取小柱进行脂肪酸分离。最后加内标液(十九碳酸甲酯)0.5 mL,用GC-MS分析。经甲酯化后采用Agilent 6850型气相色谱仪测定,在进样口为 300 ℃、H2流量为 30 mL/min、色谱柱为Agilent HP-5MS,30 μm×250 μm×0.25 μm,二阶程序升高柱温(170 ℃时按照5 ℃/min升至260 ℃,而后40 ℃/min升至300 ℃,维持 2 min),测定标准样品成功后再分析待检样品。
1.4 数据处理与分析
利用Microsoft Excel 2007进行数据处理,DPS 7.05进行方差分析(LSD法),Canoco 4.5进行多元分析。
2 结果与分析
2.1 马铃薯不同间作模式对土壤化学性状的影响
由表1可知,马铃薯收获期间作土壤养分含量与单作相比发生了较大变化。其中,马铃薯间作蚕豆土壤养分含量与单作马铃薯相比均有不同程度的增加,其全磷、碱解氮、有机质含量分别显著增加9.10%、14.54%、17.87%(P<0.05);与单作蚕豆相比,除全氮、全磷含量显著增加外,其余养分含量显著下降。经分析,在茄科与豆科的间作中,由于豆科作物根瘤菌具有固定空气氮素作用和地下根系交互作用,从而会促进氮素的转化与吸收,蚕豆收获后由于间作系统的恢复作用促进马铃薯对其他养分的吸收和转化,较单作马铃薯可以提高土壤中的养分含量。

表1 马铃薯不同间作模式对土壤化学性状的影响
与单作马铃薯相比,马铃薯间作荞麦土壤的全磷、有机质含量显著提高,其余无显著变化;与单作荞麦相比,其土壤速效磷含量显著增加,其余养分含量无显著性变化,说明在茄科与蓼科的间作系统中由于根系之间的交互作用,增加了磷肥的吸收与转化,并提高了磷的利用效率。马铃薯是喜钾作物,2种作物间作时可促进对钾的吸收,与单作荞麦相比减少土壤速效钾含量。而间作蚕豆、荞麦的土壤pH值与单作马铃薯相比分别下降1.27%、1.39%。有研究认为,间作栽培会改变土壤的pH值,进而影响土壤中养分的转化和吸收[14]。而本研究中间作模式下马铃薯土壤各养分含量较单作马铃薯处理均有不同程度的增加,可见间作系统中由于根系交互作用以及蚕豆收获后对马铃薯的恢复作用促进了土壤中的营养吸收以及养分的转化,说明合理的间作可以影响土壤pH值以及改善土壤的养分质量。
2.2 马铃薯不同间作模式对土壤酶活性的影响
土壤酶活性反映了土壤中各种生物化学过程的强度和方向,它与土壤肥力状况、理化特性和农业措施有着较为显著的相关性,是土壤肥力评价的关键指标之一[15]。由表2可知,与单作马铃薯相比,马铃薯间作蚕豆的土壤脲酶和碱性磷酸酶活性分别显著增加22.05%、20.00%,过氧化氢酶无显著变化;而与单作蚕豆相比,土壤脲酶活性显著增加,碱性磷酸酶显著下降,过氧化氢酶活性显著性下降。经分析发现,脲酶对尿素的转化和作用具有重要的影响,碱性磷酸酶可加速有机磷的脱磷速度,磷酸酶的积累对有效磷具有重要的作用,豆科作物根瘤菌的固氮作用促进了其对氮素的吸收和利用,蚕豆根系可分泌磷酸酶,从而提高了土壤中磷酸酶的活性。在马铃薯间作蚕豆的模式中蚕豆收获后,由于恢复作用后期马铃薯增加了对有效磷的吸收,从而提高了土壤中碱性磷酸酶的活性。

表2 马铃薯不同间作模式对土壤酶活性的影响
马铃薯间作荞麦与单作马铃薯相比土壤酶活性无显著性变化;与单作荞麦相比,土壤脲酶活性显著下降19.54%,碱性磷酸酶和过氧化氢酶无显著性变化。在茄科与蓼科的兼间作系统中增加了对土壤氮素的吸收,降低了脲酶的活性。过氧化氢酶是一种氧化还原酶,其作用在于破坏对生物体有毒的过氧化氢,该间作模式并没有表现出显著的差异。因此,合理的间作模式可显著提高土壤中的酶活性,对土壤中养分的吸收转化起较大的影响。
2.3 马铃薯不同间作模式对土壤微生物的影响
由表3可知,单作蚕豆的微生物总群落数最低,马铃薯间作蚕豆的微生物总群落数最高。马铃薯间作蚕豆、荞麦较单作马铃薯的真菌、细菌数量上升,放线菌数量下降,其中马铃薯间作荞麦处理中的放线菌数量显著下降36.5%(P<0.05)。而从微生物各结构所占的比例可以看出,马铃薯间作蚕豆、荞麦处理中的细菌所占比例上升,放线菌所占比例下降。不同作物根际土壤都有其特定的微生物区系,而不同间作模式对探究根际土壤中微生物结构、研究马铃薯根际土壤养分的转化过程具有重要的意义[16]。由以上结果可以推测,马铃薯与豆科、蓼科作物间作时细菌数量的累计增长会抑制放线菌的转化,从而导致放线菌数量下降,对土壤中养分的转化和吸收产生影响。

表3 马铃薯不同间作模式对土壤微生物群落结构组成的影响
磷脂脂肪酸(PLFA)是土壤经甲基化提取磷脂成分后得到的脂肪酸产物,其中磷脂是活细胞膜的基本成分,具有多样性和生物学特异性,且脂肪酸只存在于活细胞中,因此PLFA适用于微生物群落的动态监测,常被用于土壤微生物群落结构的测定[17-18]。由表4可知,单作马铃薯土壤微生物主要类群的PLFA含量最高,单作荞麦主要类群的PLFA含量最低,其中单作荞麦革兰氏阳性菌(G+)中的含量与单作马铃薯相比差异达到显著性水平(P<0.05)。间作模式中土壤微生物群落的PLFA含量与单作马铃薯相比发生了较大的变化,其中间作荞麦的菌根真菌(AMF)中的含量显著下降55.6%(P<0.05)。
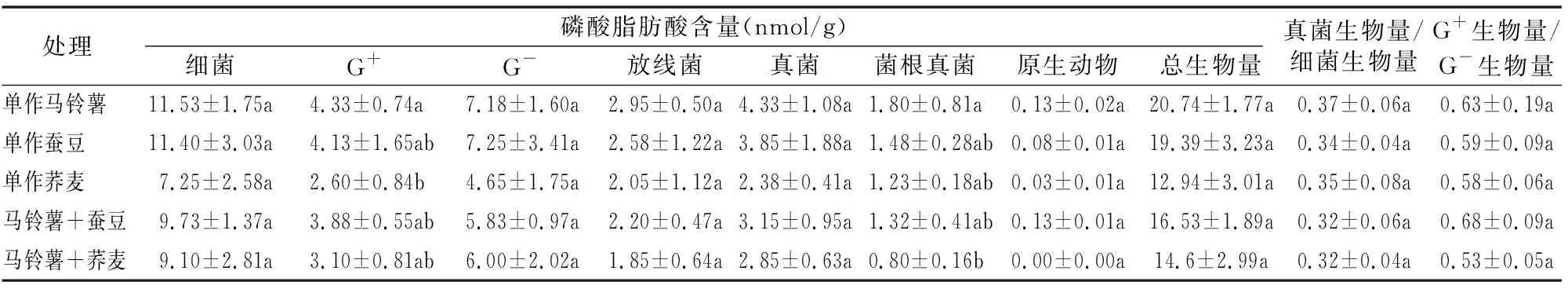
表4 马铃薯不同间作模式对土壤微生物群落生物量的影响
2种间作模式中革兰氏阳性菌(G+)和革兰氏阴性菌(G-)的生物量以及细菌的生物量与单作马铃薯相比出现不同程度的下降趋势,马铃薯间作荞麦的细菌生物量较单作荞麦有所增加,但差异不显著。马铃薯间作荞麦的总生物量较单作荞麦升高,其余间作模式中的土壤微生物总生物量均低于各自单作对应的总微生物量,但均无显著性差异。马铃薯间作荞麦中的G+生物量/G-生物量以及马铃薯间作蚕豆、间作荞麦中的真菌生物量/细菌生物量较单作马铃薯有所降低,间作蚕豆的G+生物量/G-生物量的值升高,但均无显著性差异。
由以上结果可知,茄科与豆科、蓼科间作会减少土壤中细菌、放线菌、真菌等微生物主要类群的生物量,说明茄科与豆科、蓼科间作改变了土壤微生态的环境,在一定程度上削弱了马铃薯根际土壤微生物主要类群PLFA的生物量;而真菌生物量/细菌生物量的值降低和间作蚕豆中G+生物量/G-生物量的值升高,均表明间作有利于促进土壤向高肥效的“细菌型”土壤类型转变。
2.4 土壤微生物群落结构组成与土壤因子之间的相互关系
多元分析结果(图1-A、图1-B)显示,各排序轴都能在超过56%的累计贡献率上解释土壤微生物结构及微生物生物量变化与环境因子间的关系。排序图(图1-A、图1-B)反映了各根际土壤微生物群落结构最适宜生活的土壤环境,各处理点分散较为明显,说明马铃薯不同间作模式下形成了不同的土壤微生态环境,各处理间有差异。由图1-A可知,排序轴累积解释的信息量达70.1%。因此,基于特征向量载荷因子相联系的排序轴1和轴2最大程度地解释了不同处理之间土壤环境因子对土壤微生物群落结构的影响。土壤细菌、真菌数量除与全氮含量成锐角外,与其他环境因子都成钝角;而放线菌数量与全磷含量、速效氮含量、pH值成锐角,与其他环境因子呈钝角。由于图1-A中2条射线代表其相关性,锐角代表呈正相关,其夹角越小,相关性越强,钝角与之相反,钝角越大负相关性越大。说明细菌、真菌数量与全氮含量呈正相关性,受全氮含量环境因子影响较大,与其他环境因子呈负相关性。放线菌数量与土壤全磷含量、速效氮含量、pH值呈正相关性,与其他环境因子呈负相关性。
图1-B显示了不同处理下微生物生物量与环境因子的制约关系,土壤微生物生物量与环境因子也表现出一定的相关性。其中,菌根真菌生物量与全氮含量呈负相关性,而与其他环境因子都呈正相关性,原生动物、细菌、放线菌、真菌、G+、G-生物量与有机质、全磷、速效磷、速效钾的含量有良好的相关关系,与其他环境因子呈负相关性。由图1可以看出,5种种植模式分别位于不同的象限,说明种植模式不同,土壤微生物结构和土壤微生物生物量不同,受环境因子的影响也不相同。综合可知,马铃薯不同间作模式下土壤微生物结构和微生物生物量的变化与土壤环境因子有着密切联系,受多种环境因子制约。

3 讨论
3.1 不同间作模式对土壤理化与生物学性状的影响
土壤养分是作物摄取养分的主要来源之一,在作物养分吸收总量中占有很高的比例,土壤养分含量的高低关系着植株能否健康生长或生存。已有大量的研究表明,合理的间作可以提升土壤中的养分含量。郝艳如等在玉米、小麦间作的研究中表明,间作模式提高了土壤中的有效养分含量,改善了土壤根部供肥能力和养分吸收环境[19];代会会等在豆科间作试验中发现,间作可以增加土壤中全氮、速效氮、速效钾和速效磷含量,并且提高作物产量[20]。本研究结果表明,在茄科与蓼科、豆科的间作中,不同的间作模式与单作马铃薯相比不同程度地提高了土壤中的养分含量,降低了pH值,其中间作蚕豆的碱解氮、有机质含量,间作荞麦的速效磷、全氮、有机质含量增加较多,且差异达到显著水平(P<0.05),这与代会会等的研究结果[20]较为一致。经分析发现,茄科在与豆科、蓼科的间作系统中由于空间、根系和生理等方面的需求差异以及互补性,使得土壤中的养分和自然资源得到最大的利用,从而提高土壤中的养分含量。
土壤酶是生态系统的能量流动和物质交换等生态过程中活跃的生物活性物质,且土壤碱性磷酸酶、过氧化氢酶、脲酶的活性能够分别反映出土壤磷素供应状况、有机质的转化速度和土壤氮素水平状况,对农作物的生长起着至关重要的作用[12]。柴强等在研究间甲酚对间作模式中土壤微生物和酶活性的影响中指出,通过玉米+蚕豆发现间作模式对土壤磷酸酶和过氧化氢酶活性具有极显著影响[21];Zhou等在黄瓜间作洋葱、大蒜的研究中表明,间作模式下土壤脲酶和过氧化氢酶活性高于黄瓜单作[22]。本研究结果表明,间作模式下磷酸酶、脲酶、过氧化氢酶活性升高,其中间作蚕豆的磷酸酶和脲酶活性与单作对照相比差异显著,这与姜莉等的研究结果[23]较为一致。由于豆科作物根瘤菌的固氮作用以及收获后马铃薯的恢复作用对间作马铃薯养分的吸收有较大影响,并且间作模式下作物之间对土壤养分的相互作用和利用促进了土壤酶活性的变化,使得酶活性与土壤环境因子联系变化更为敏感。
3.2 不同间作模式下土壤理化性状对土壤微生物群落结构的影响
土壤微生物参与土壤的物质交换和能量转化,土壤酶参与土壤中许多重要的生物化学过程,二者共同成为评价土壤肥力的重要指标[24]。而合理的间作可以改变土壤的微环境,由于根系分泌物、作物残体和根系残体在土壤中的累积,供给土壤微生物的营养物质增加,因此增强了土壤微生物的活性,增加了土壤微生物的群落结构多样性,进而形成与之相适应的微生物区系[25]。很多研究结果表明,合理的间作可以提高作物根际土壤微生物的数量[26]。其中宋亚娜等对小麦+蚕豆、玉米+蚕豆和小麦+玉米间作体系进行研究时发现,间作能改变根际细菌群落结构的组成,且证明了间作体系地上部多样性与地下部多样性存在着紧密联系[27];马琨等在间作栽培对土壤微生物群落的影响中表明,间作栽培显著地改变了根际土壤微生物主要功能群落的结构[28];吴娜等在马铃薯间作燕麦的研究中指出,马铃薯+燕麦(4 ∶2)的种植比例能改善根际土壤微生态环境,优化根际土壤微生物群落结构[29]。Artursson等研究认为,根系分泌物中某种物质促进了根际土壤中革兰氏阳性细菌、AMF真菌的累积,从而相应地增加了微生物的生物量[30]。
本试验结果表明,不同间作模式下间作处理增加了真菌、细菌的数量,减少了放线菌的数量,但差异均不显著。真菌比例变化较小、细菌比例增加,使土壤向着更高效的细菌型土壤转化,而微生物群落总数的增加增大了微生物环境的多样性。间作处理中土壤微生物群落细菌、放线菌、真菌的生物量较对照组减少,但细菌中革兰氏阳性菌生物量比例增加、真菌生物量/细菌生物量的值降低,表明间作模式能改变马铃薯根际土壤微生物群落的组成及环境的对应关系。
由以上结果说明间作条件下,茄科与豆科或蓼科作物之间的根系横向交叉,其根系分泌物改变了间作模式下土壤微生物的区系结构,进而影响了马铃薯根际土壤微生物的数量和活性,改变了土壤微生物群落的结构组成。由Canoco多元分析表明,土壤养分的转化和吸收与微生物群落结构和微生物生物量的变化呈紧密的相关关系,说明间作模式不仅能提高土壤养分含量、酶活性,还能改变土壤中的微生态环境,产生与环境因子相适应的微生物区系。
4 结论
马铃薯间作蚕豆提高了土壤碱解氮、有机质的含量,间作荞麦提高了土壤速效磷、全氮、有机质的含量,且差异达到显著性水平(P<0.05);马铃薯间作蚕豆显著地提高了土壤磷酸酶和脲酶活性,而马铃薯间作荞麦土壤酶活性与对照相比无显著差异;间作模式降低了土壤中放线菌的比例,提升了土壤中细菌的比例,并且改变了土壤中微生物主要类群的PLFA含量;马铃薯不同间作模式下土壤养分与微生物群落结构的变化较为紧密。
猜你喜欢
经济林研究(2022年4期)2023-01-12
广西林业科学(2022年2期)2022-05-09
小读者(2021年2期)2021-03-29
河北果树(2020年4期)2020-11-26
公民与法治(2020年13期)2020-07-27
山东农业科学(2019年11期)2019-12-24
学生天地(2018年9期)2018-09-06
干旱地区农业研究(2017年5期)2017-12-18
现代园艺(2017年11期)2017-06-28
新疆农垦科技(2016年2期)2016-08-21
