
猪伪狂犬病病毒HB-11株的分离鉴定及gC基因的分析
2018-12-05 08:59马晶晶吴碧清高俊锋郑良益舒银辉
中国兽医杂志 2018年8期
马晶晶,吴碧清,高俊锋,郑良益,舒银辉,赖 志
(1. 上海创宏生物科技有限公司,上海 松江 201619;2. 武汉中博生物股份有限公司,湖北 武汉 430070)
猪伪狂犬病(Porcine pseudorabies,PR)是甲型疱疹病毒亚科猪疱疹病毒I型引起的一种急性传染病[1-2]。此病可引起妊娠母猪繁殖障碍、流产、死胎、呼吸症状;新生仔猪除出现神经症状外,还可侵害消化系统,临床症状主要表现为腹泻、呕吐、生长不良等。猪既是本病原的原发感染宿主,又是病毒的长期贮存者和排毒者,在猪伪狂犬病的传播上起着重要作用。
猪伪狂犬病病毒(PRV)属于疱疹病毒科,α-疱疹病毒亚科,基因组为双链DNA,全长150 kb,其中gC含量在基因组中最高达73%,至少含有70个基因,可编码100多种蛋白质[3-4]。gC是PRV的一种重要糖蛋白,虽然gC不是PRV复制所必需的,但作为PRV最主要的保护性抗原之一,它不仅能在病毒对靶细胞的吸附、释放和病毒毒力方面产生作用,还能诱导细胞免疫,产生中和抗体,还能激活补体系统,产生防御作用[5-7]。
自2011年以来,我国华北、华东、华中、华南等地区出现猪伪狂犬病流行,造成母猪流产、产死胎、木乃伊胎,仔猪、保育猪群出现大量死亡。最近研究表明,与以往的流行株相比,新流行株的抗原性已发生了变异,gC基因存在抗原飘移现象,从而导致现有疫苗不能提供完全保护[8-9]。我们于2015年9月,在河北某猪场大批死亡育肥猪群的脑及肺脏中分离到1株病毒,并进行了全面鉴定,并对其进行了gC基因的扩增和序列分析,以期获得流行毒株PRV分子变异趋势,为进一步鉴别PRV疫苗株和野毒株提供理论依据,为丰富PRV的分子流行病学和疫苗的研制提供参考依据。
1 材料
1.1 细胞 BHK-21细胞、IBRS-2细胞、HeLa细胞、Vero细胞,均购自中国典型培养物保藏中心(武汉大学)。原代鸡胚成纤维细胞由武汉中博生物股份有限公司按常规方法进行制作。
1.2 病毒 猪伪狂犬病病毒闽A株、Bartha K61株,购自中国兽医药品监察所。
1.3 血清 猪伪狂犬病病毒标准阴性、阳性血清由武汉中博生物股份有限公司制备。新生犊牛血清,购自杭州新锐生物工程有限公司,使用前56 ℃灭活30 min。
1.4 DMEM培养基 购自北京清大天一生物技术有限公司,货号:MD202。
1.5 实验动物
1.5.1 BALB/c小鼠 清洁级,体重20~22 g,购自武汉生物制品研究所。
1.5.2 家兔 普通级,体重1.8~2.0 kg,购自武汉市万千佳禾实验动物养殖有限公司。
1.5.3 断奶仔猪 6~7周龄,来自健康猪群,PRV中和抗体效价不高于1∶2,PCR抗原检测为阴性,购自武汉市江夏区鲁湖猪场。
2 方法
2.1 病料处理 对猪场发病死亡的仔猪采集脑和肺组织,混合后在灭菌的研钵内剪碎,用DMEM作1∶5倍稀释成悬液,-70 ℃反复冻融3次。经3 000 r/min 离心30 min,取上清液经0.22 μm微孔滤膜过滤后,加入适量青、链霉素,-70 ℃保存作为接种材料。
2.2 病毒分离 将上述处理的样品接种于已长成单层的BHK-21细胞的培养瓶中(100 mL),每瓶接种2 mL,接种3瓶,设正常细胞对照1瓶。置于37 ℃恒温箱吸附1 h,然后加入DMEM维持液8 mL。培养并观察3日,盲传3代,有细胞病变效应(CPE)的继续传代,直至稳定。并用具有PRV典型CPE特征的培养物分别接种IBRS-2细胞、HeLa细胞、Vero细胞和原代鸡胚成纤维细胞。同时用PRV闽A株接种上述各细胞,比较两者所致CPE情况。
2.3 分离病毒的克隆纯化和培养 用维持液将病毒液作10倍系列稀释,取10-3、10-4、10-5、10-6、10-7、10-86个稀释度接种已长成BHK-21细胞单层的平皿,每个稀释度接种4个平皿,每个平皿接种0.1 mL。经37 ℃培养箱孵育1 h后,加入44 ℃营养琼脂,每个平皿10 mL,待凝固后翻转平皿,置于5%二氧化碳培养箱内培养。48 h后加入0.01%中性红5 mL覆盖,培养72 h后选取产生空斑的平皿,选择小而孤立的空斑挑出,加入含0.5 mL营养液的离心管中,经冻融后以3 000 r/min 离心10 min,取上清进行扩大培养。
2.4 病毒含量的测定 纯化后,取在BHK-21细胞适应的病毒液,采用微量法测定其在BHK-21细胞上的感染力。同样方法测定PRV闽A株在BHK-21细胞上的TCID50。
2.5 病毒的鉴定
2.5.1 电镜形态的观察 取病毒悬液反复冻融3次,以3 000 r/min离心30 min,取上清液以 40 000 r/min 离心3 h,沉淀物用2%磷钨酸钠负染,观察病毒粒子形态。
2.5.2 中和试验 将分离株用灭菌生理盐水稀释成为每0.1 mL含有200 TCID50的病毒悬液,与等量的抗猪伪狂犬病病毒特异性血清充分混合,37 ℃作用1 h后,接种形成良好单层BHK-21细胞培养板,接种6孔,置37 ℃培养。同时设立不中和病毒悬液为阳性对照组,置同条件下培养。5日后观察各细胞孔是否出现CPE。
2.6 病毒的血清学特异性
2.6.1 制备PRV HB-11抗血清 将HB-11病毒纯化、培养后,通过甲醛灭活制备抗原,将制备好的抗原与弗氏完全佐剂按1∶1混合肌肉注射SPF鸡,第7天将抗原与不完全弗氏佐剂按照1∶1混合进行第2次免疫,第14天将抗原第3次免疫SPF鸡。第3次免疫7日后,采血,制备HB-11的阳性血清。把制备的血清置56 ℃水浴灭活30 min,分装后置-20 ℃冻存。
2.6.2 抗血清中和试验 采用固定血清稀释病毒法,将分离得到的14株分离病毒株和疫苗毒株闽A株、Bartha K61株作10倍系列稀释,使其稀释度为10-1~10-8,然后各个稀释度的病毒液与等量伪狂犬分离株HB-11的阳性血清混合,置于37 ℃作用1 h,再接种96孔细胞板,每个稀释度6孔,每孔100 μL,同时设未加抗血清的对照。48~96 h后观察CPE,判定结果,计算PRV HB-11抗血清对各PRV毒株的血清中和指数。
2.7 动物毒力试验
2.7.1 对家兔的毒力试验 将分离培养的病毒悬液F5代接种健康家兔2只,每只在臀部皮下注射2 mL,并设注射DMEM培养液的健康家兔2只作为对照。连续观察5日。记录临床反应观察结果。
2.7.2 对小鼠的毒力试验 将分离培养的病毒悬液F5代接种BALB/c小鼠4只,每只在腹部皮下注射0.1 mL,并设注射DMEM培养液的BALB/c小鼠4只作为健康对照。连续观察5日。记录临床反应观察结果。
2.7.3 对断奶仔猪的毒力试验 试验前筛选6~7周龄仔猪15头(PRV中和抗体效价不高于1∶2,PCR抗原检测为阴性),随机分为3组,每组5头。A组接种病毒原液(108.33TCID50/mL),B组接种病毒含量为106.0TCID50/mL,C组为空白对照,接种方式为每头肌肉注射3 mL同时滴鼻3 mL。每日测量体温,观察仔猪临床症状,并计算3 d平均值,绘制评分表。剖检病死猪,观察肺和脑部病理变化;攻毒后观察21日,剖检所有未死亡仔猪,观察肺部病变,记录肺部评分(临床评分法按照参考文献[12]方法进行。肺部评分按照参考文献[13]方法进行)。
2.8 gC基因测序
2.8.1 引物合成 根据GenBank中登录的PRV全基因序列设计gC基因段引物:gC-F:5′-ATGGCCTCGCTCGCGTGCGCTC-3′,gC-R:5′-TCACGGCCCCGCCCGGCGGTAGTAG-3′,扩增片段大小为 1 440 bp。引物由上海捷瑞生物工程有限公司合成。
2.8.2 病毒DNA提取及PCR鉴定 使用北京天根生化科技有限公司DP315试剂盒说明书提取PRV DNA,利用上述引物扩增PRV gC基因全序列。PCR反应总体积25 μL:DNA模板2 μL,2×gC Rich Buffer 12.5 μL,上下游引物各0.5 μL,ddH2O补至25 μL。PCR程序:94 ℃ 5 min;94 ℃ 1 min;53 ℃ 1 min,72 ℃ 2 min,35个循环;72 ℃ 10 min。PCR产物用1.5%琼脂糖凝胶电泳检测。
2.8.3 gC基因的克隆与序列分析 将目的基因胶回收克隆至pMD18-T载体中,送上海杰李生物技术有限公司进行序列测定,每个样品选择3个克隆,每个克隆测序3次。利用生物信息软件对此分离株及近年本实验室分离到的变异株和疫苗株Bartha株gC基因序列进行比对分析。
3 结果
3.1 病毒分离结果 将经过除菌处理的脑和肺脏组织悬液样品,接种BHK-21细胞,传至第5代,稳定在24~36 h可见细胞肿胀、出现颗粒、失去光泽,继而圆缩、脱落,并可见到许多巨细胞,与PRV闽A株所致细胞病变类似。将第5代细胞培养物接种猪肾传代细胞IBRS-2、HeLa细胞、Vero细胞以及原代鸡胚成纤维细胞,均产生典型细胞病变(见图1),表明分离株能在不同细胞上增殖。

图1 细胞病变图
注:从左往右依次为未接毒,接毒24 h,接毒36 h的细胞图
3.2 病毒克隆纯化 分离病毒在平皿上经过3日培养后,在产生空斑细胞单层(10-4稀释度)上可见单个的可数空斑,空斑直径在1 mm~4 mm之间,大小不一,挑出直径1 mm的空斑。重复克隆2次后接种BHK-21细胞,传代3次后,75%CPE出现时间稳定在24~36 h,表明获得一株纯病毒。
3.3 病毒含量测定 按Reed-Muench法计算,所分离病毒和PRV闽A株在BHK-21细胞上的病毒含量分别为108.33TCID50/mL和107.75TCID50/mL。
3.4 病毒的鉴定
3.4.1 电镜观察结果 病毒上清液经超高速离心,取沉淀物经磷钨酸钠负染后在电镜下见到典型的PRV颗粒。病毒粒子形状呈圆形,病毒颗粒大小为150 nm左右(见图2)。

图2 病毒电镜图片 (40 000×)
注:箭头所指处为PRV颗粒
3.4.2 中和试验结果 以200 TCID50的病毒液与倍比稀释的PRV标准阳性血清反应,按Reed-Muench法测得标准阳性血清的中和效价为1∶32。试验结果表明,该分离病毒为PRV。
3.5 PRV HB-11血清学特性试验结果 将6株伪狂犬病病毒分离株、闽A株、Bartha K61株按10倍比稀释后,与等量PRV HB-11的阳性血清混合后,接种细胞,37 ℃培养96 h后观察到CEF上无CPE,试验结果证明,15株伪狂犬病病毒分离株、闽A株以及Bartha K61株病毒能被PRV HB-11阳性血清中和,中和指数为832~6 761,结果见表1。

表1 PRV HB-11阳性血清对各毒株的中和指数
3.6 动物毒力试验结果
3.6.1 对小鼠和家兔的接种试验结果 以所分离的病毒悬液接种家兔和小鼠均产生了典型的伪狂犬病症状。接种36 h后家兔表现为奇痒,用嘴啃咬注射部位,使注射部位被毛脱落,皮肤受损,暴露出红色肌肉,头、颈、背部等多处擦伤,48 h后四肢出现麻痹而死亡,注射DMEM维持液的对照家兔均健活;小鼠于接种后24 h表现为狂躁不安,于接种后48 h自行啃咬注射部位,4只小鼠于攻毒60 h死亡,注射DMEM培养基的对照小鼠均健活。结果表明,分离病毒液对小鼠和家兔的毒力较强。
3.6.2 对断奶仔猪的接种试验 接种病毒原液组A组(108.33TCID50/mL),断奶仔猪全部死亡(5/5)。接种106.0TCID50/mL B组,仔猪均体温升高,并出现不同程度的食欲下降、精神不振、呼吸困难等临床症状,其中2头仔猪(H2、H5)出现了神经症状。H3在攻毒后第9天死亡,H5在攻毒后第11天死亡,共死亡2头仔猪(2/5)。剖检攻毒试验仔猪,肺部均表现出血、水肿、坏死,仔猪脑有出血、充血病变;空白对照组C组无任何临床症状,体温在38.5 ℃~39.5 ℃,剖检后观察,肺和脑部无任何病理变化,结果见临床症状评分(图3)。结果表明,新分离的HB-11株为猪伪狂犬病病毒强毒株。
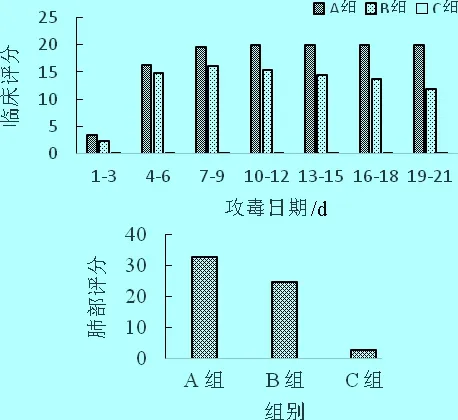
图3 攻毒后仔猪临床评分及肺部评分
注:临床评分法按照参考文献[12]方法进行;肺部评分按照参考文献[13]方法进行
3.7 gC基因PCR扩增结果 将此分离株和近年本实验室分离到的PRV-JX,PRV-X两株新流行毒株及疫苗株Bartha进行PCR扩增后,凝胶电泳结果显示,4株PRV毒株在1 440 bp均有扩增条带,见图4。
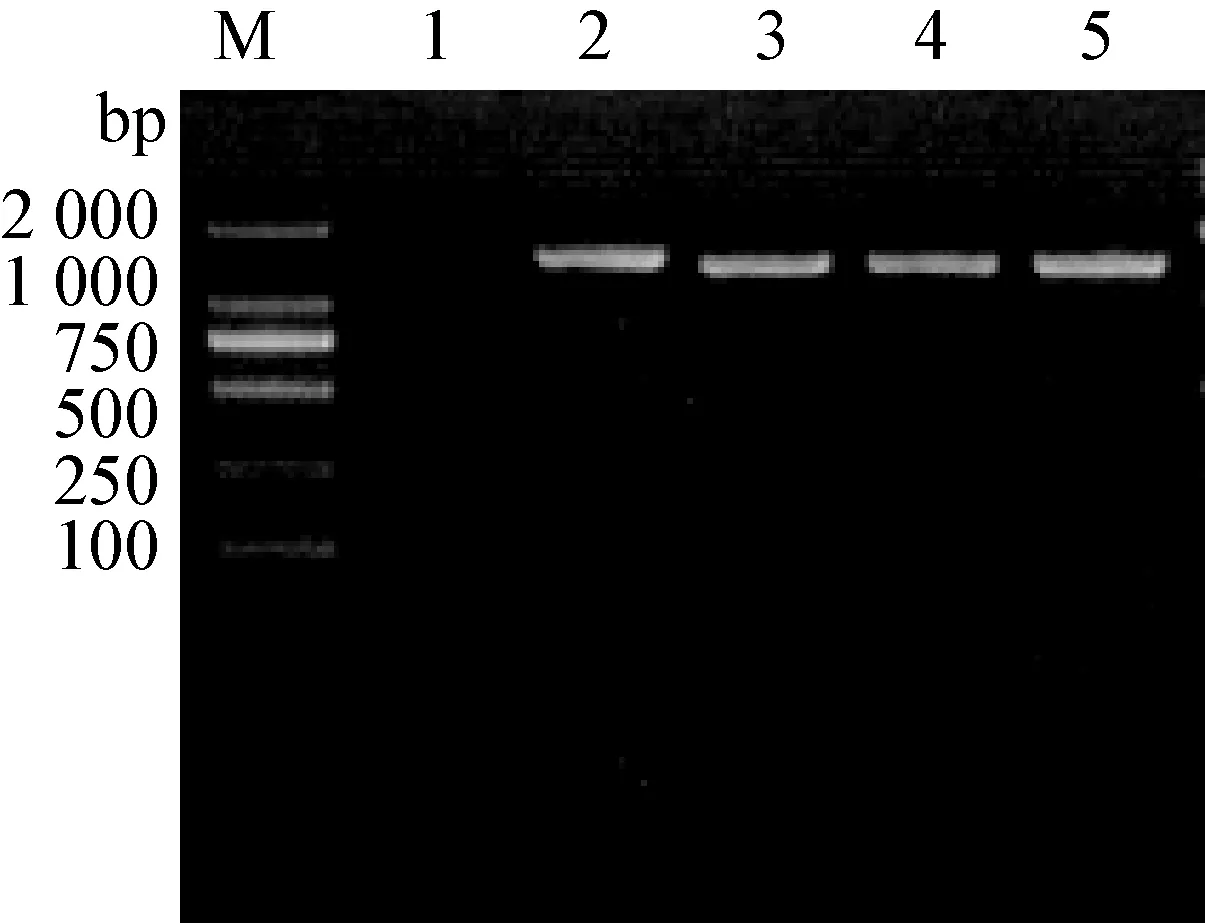
图4 PRV gC基因PCR扩增图
M:DNA分子质量标准; 1~5依次为阴性对照,PRV HB-11株,PRV-JX株,PRV-X株,Bartha株
3.8 gC基因序列分析 将该分离株测序结果与本实验室及近年本实验室分离到的变异株PRV-JX株,PRV-X株和疫苗株Bartha株gC基因核苷酸序列及推导的氨基酸序列进行比对分析。结果表明,PRV HB-11株与PRV-JX株,PRV-X株和疫苗株Bartha株gC基因序列的同源性分别为99.3%,99.4%和94.8%(见图5),说明该分离株与近年本实验室分离到的新流行毒株同源性较高,与Bartha疫苗株的同源性相对较低。氨基酸序列比对结果显示,分离株与PRV-JX株,PRV-X株相对于疫苗Bartha株的gC蛋白中有相同的7个氨基酸的插入(158AAASTPA164)(见图6)。

图5 分离株与PRV-JX株,PRV-X株和疫苗株Bartha株gC基因核苷酸序列的比对结果

图6 分离株与PRV-JX株,PRV-X株和疫苗株Bartha株gC基因氨基酸序列的比对结果
4 讨论
从河北某猪场的病死仔猪中分离到的一株病毒,在BHK-21细胞上产生CPE,并且能与PRV标准阳性血清发生中和反应。病毒在电镜下可观察到150 nm左右的典型的伪狂犬病病毒粒子,用所分离的病毒接种家兔、小鼠和仔猪均出现典型的伪狂犬病症状,这一结果证实所分离的毒株是猪伪狂犬病病毒毒株。
本试验中,测得的猪伪狂犬病病毒HB-11株在BHK-21细胞上的TCID50为108.33TCID50/mL,该病毒接种家兔2只、小鼠4只不仅均出现典型的伪狂犬病症状,而且均全部死亡,106.0TCID50/mL接种5头仔猪,每头猪颈部肌肉注射3.0 mL,同时滴鼻3.0 mL可导致仔猪5/5出现临床症状并出现2/5死亡,剖检发病猪的肺部病理变化,均表现出血、水肿、坏死等病变。说明分离的猪伪狂犬病病毒是一株毒力较强的野毒株,命名为HB-11株。
PRV-gC基因序列比对分析结果显示,猪伪狂犬病病毒HB-11株与近年本实验室分离到的PRV变异株JX株和X株同源性较高,与Bartha疫苗株亲缘关系较远,仅为94.8%;氨基酸序列比对结果显示,其gC基因序列相对于Bartha疫苗株序列有7个连续氨基酸的插入(158AAASTPA164),表明近年来PRV的分离株正向远离疫苗株的方向变异,这也进一步证明了2012年以来猪伪狂犬病的暴发可能由于现有的猪伪狂犬疫苗株Bartha株已不能对新流行的伪狂犬变异株提供完全保护。流行毒株PRV-gC基因的研究分析对于研究PRV多样性及分子进化具有重要意义,有助于实验室在分子水平上确定猪伪狂犬病的存在和发生原因,为猪场的免疫方案提供更准确的技术依据。gC基因序列之间的差异是否会影响猪伪狂犬病病毒HB-11的毒力和抗原性的变化,还需进行全基因测序分析PRV的其他基因组内编码的蛋白之间的相互作用,以期为新疫苗的研发及猪伪狂犬病的防控提供理论依据。
猜你喜欢
科学大观园(2022年2期)2022-01-23
文萃报·周二版(2021年47期)2021-12-14
今日农业(2021年5期)2021-11-27
今日农业(2021年13期)2021-11-26
今日农业(2021年21期)2021-11-26
今日农业(2021年20期)2021-11-26
动漫星空(兴趣百科)(2020年3期)2020-03-24
中国猪业(2017年11期)2017-12-11
中华实验和临床病毒学杂志(2017年4期)2017-01-15
中华实验和临床病毒学杂志(2017年5期)2017-01-13
