
17β-雌二醇/微小核糖核酸-21信号通路抑制低氧性肺动脉高压肺血管重构的机制研究
2018-11-24 05:58沈丽晓袁博云王丽袁雅冬
中国循环杂志 2018年11期
沈丽晓,袁博云,王丽,袁雅冬
肺动脉高压(PH)是一种临床高发的慢性肺循环疾病,低氧性肺动脉高压(HPH)属于PH中的第三大类,肺小动脉收缩与血管重建是其发生、发展的病理生理学基础[1]。雌激素在体内主要由卵巢成熟滤泡分泌,行卵巢切除术后自身分泌的雌激素水平会明显降低,而17β-雌二醇(E2)被认为是体内天然存在的三种雌激素(包括雌二醇、雌酮和雌三醇)中生物活性最强的[2],关于其对肺血管保护作用的研究也日益增多。微小核糖核酸(miRNA)是一组长约22个核苷酸的单链非编码小RNA[3],主要在转录后水平调控基因的表达。近期研究显示,miRNA介导肺动脉平滑肌细胞(PASMC)增殖的各种分子信号转导机制在HPH发生、发展中起着关键作用[4,5]。本研究旨在建立慢性去势HPH大鼠模型,研究经E2干预后miRNA-21(miR-21)、增殖细胞核抗体(PCNA)在肺动脉中表达的变化,并进一步体外培养人肺动脉平滑肌细胞(hPASMC),验证E2对肺血管的保护作用是否通过下调miR-21表达进而影响PASMC增殖而实现。
1 材料与方法
1.1 实验动物及细胞
6~7周龄SD雌性大鼠32只,体重(200±10)g,购于河北医科大学实验动物中心(合格证编号:1511341)。hPASMC购自美国Sciencell公司。
1.2 试剂及仪器
低氧环境动物试验箱(CJ-DO2245,长沙长锦科技有限公司);MCO-15AC二氧化碳(CO2)培养箱(三洋公司,日本);Thermo3131三气培养箱(赛默飞世尔科技公司,美国);双色近红外成像系统Odyssey CLx(LI-COR公司,美国);BioTek Cytation 3细胞成像酶标仪(BioTek仪器有限公司,美国);Powerlab生理记录仪(AD器械有限公司,澳大利亚);Gene Amp 聚合酶链式反应(PCR)系统9700(ABI公司,美国);7500 实时PCR 系统(ABI公司,美国);电泳、电转仪(北京六一仪器厂);E2(进口分析纯度, Sigma公司,美国);兔抗大鼠PCNA抗体(武汉proteintech公司);近红外荧光抗兔二抗(Abcam公司,英国);Trizol试剂(Invitrogen公司,美国);实时PCR试剂盒及反转录试剂盒(Promega公司,美国);引物(Invitrogen公司,美国)。平滑肌细胞培养基(SMCM)、平滑肌细胞生长因子(SMCGS)、胎牛血清(FBS)均购自美国Sciencell公司。
1.3 实验方法
1.3.1 模型制备
动物实验:大鼠适应性饲养一周后行卵巢切除术,术后1周随机分入常氧组、常氧+E2组、低氧组、低氧+E2组,每组8只。两个E2干预组大鼠每日皮下注射E2 20 μg/kg,余组大鼠注射等量生理盐水。两个低氧组大鼠置于氧气浓度为(10.0±0.5)%的低氧环境动物试验箱中,以钠石灰和无水氯化钙吸收多余的CO2和水分,控制CO2浓度<0.5%,每天持续低氧24 h,两个常氧组置于正常空气中饲养外,余饲养条件同低氧组。
细胞实验:将hPASMC在专用培养液(配方:2% FBS、0.5% SMCGS、0.5%青霉素/链霉素溶液和SMCM)中培养,隔天换液1次,第5~8代细胞用于实验。细胞生长达50%融合后,用不含FBS的无酚红培养基孵育24 h,使细胞生长同步化,传代后分为3组:常氧组、低氧组、低氧+E2(10-6mol/L)组。常氧组置于37℃、5% CO2、95%空气的MCO-15AC CO2培养箱中孵育24 h,两个低氧组置于37℃、5%CO2、92%氮气的Thermo3131三气培养箱中孵育24 h,收集细胞进行下游实验。
1.3.2 肺小动脉形态观察及平均肺动脉压(mPAP)、右心室肥厚指数(RVHI)测定
饲养8周后,大鼠腹腔内麻醉,将连接压力传感器的微导管插入钝性分离的右侧颈外静脉,连接Powerlab生理记录仪记录mPAP。测压结束后放血处死,留取左肺上叶制成石蜡标本,行苏木素伊红(HE)染色镜下观察肺小动脉形态;取出大鼠心脏,剪去心房及脂肪、血管等组织,沿室间隔边缘将右心室游离壁分离,吸干水分后用电子天平分别称取右心室(RV)和左心室+室间隔(LV+S)的重量,计算RVHI来反映右心室肥厚程度的变化,公式:RVHI=RV/(LV+S)×100%。留取肺组织并游离出肺动脉,待测miR-21及PCNA表达情况。
1.3.3 四甲基偶氮唑蓝比色法(MMT)检测细胞增殖
将细胞以5 000/孔接种于96孔板上,每孔设5个复孔,周围设只加培养液的空白对照孔。同步化处理后常氧组、低氧组、低氧+E2组干预24 h后每孔加入20 μl MTT溶液(0.5% MTT),继续培养4 h后,弃去孔内培养液,每孔加入150 μl二甲基亚砜(DMSO),水平摇床上低速避光震荡10 min,应用BioTek Cytation 3细胞成像酶标仪测定各孔在490 nm处的光密度(OD)值。空白孔调零,实验重复三遍。
1.3.4 实时PCR检测肺动脉及hPASMC中miR-21、PCNA信使RNA(mRNA)水平
用Trizol法分别提取肺动脉及细胞中的总RNA,应用NanoDrop2000超微量分光光度计测定总RNA浓度和纯度,理想RNA纯度为A260/A280在1.8~2.0之间。按反转录试剂盒(Promega Goscript A5001)在冰上进行操作,将2 μg总RNA加入到20 μl体系中合成cDNA,其中反转miR-21时需加入特异miRNA的茎环引物1 μl,应用Gene Amp PCR系统9700反转录成cDNA。将2 μl cDNA加入到含目的基因上下游引物各 0.5 μl、2×GoTaq qPCR Master Mix 10 μl的20 μl反应体系中,在7500 实时PCR系统上进行扩增。扩增程序为:95℃ 2 min预变性,(95℃15 s,60℃ 1 min)×40个循环。重复实验3次,按照F=2-△△ct分析结果。实时PCR检测中用到的引物序列见表1。
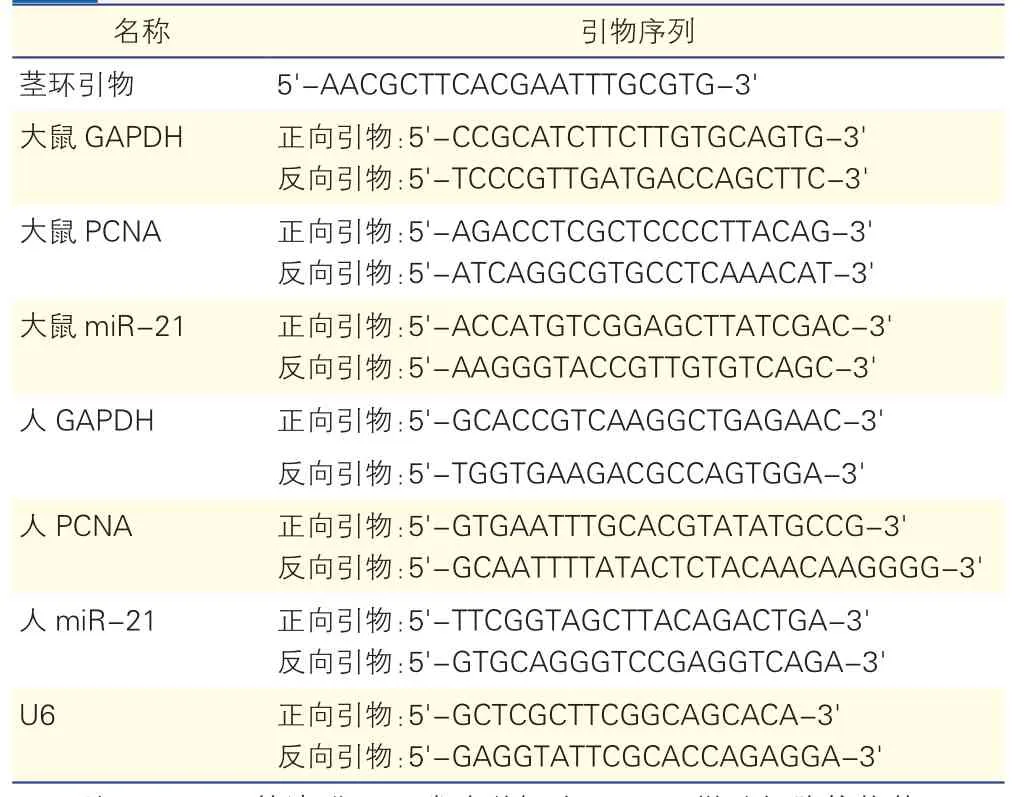
表1 实时聚合酶链式反应检测中的引物序列
应用SPSS 21.0统计软件进行数据处理,正态分布的计量数据以±s的形式表示,多个样本均数的两两比较采用方差分析中的LSD和SNK-q检验,P<0.05为差异有统计学意义。
1.3.5 免疫印迹法检测肺动脉及hPASMC中PCNA蛋白水平
用加入蛋白酶抑制剂的RIPA裂解液,按说明书分别提取肺动脉及细胞中的总蛋白,NanoDrop2000超微量分光光度计测定蛋白浓度。将蛋白样品与SDS-PAGE上样缓冲液混匀后95℃变性10 min。取等量蛋白样品,经10% SDS-PAGE,80 V恒压电泳,当预染蛋白marker出现2个条带时将电压调至120 V,所有条带均出现后结束电泳。恒定电流200 mA 90 min,转膜至聚偏二氟乙烯膜(PVDF)上。将PVDF膜5%脱脂奶粉的TBS中室温封闭1 h,分别放入TBST液稀释的一抗(PCNA 1:1 000,GAPDH 1: 2 000),4℃反应过夜,次日TBST洗膜三次,每次10 min,然后放入近红外荧光(680 nm)抗兔二抗工作液(1: 10 000)中室温避光孵育1 h,TBST洗膜三次,每次10 min,打开双色近红外成像系统Odyssey CLx ,将膜放在该仪器的成像面板上进行扫膜,并应用其中的Image Studio软件分析数据。
1.4 统计学方法
2 结果
四组大鼠肺动脉组织形态、mPAP和RVHI比较:8周后,光镜下常氧组大鼠肺动脉管壁结构正常,内皮细胞扁平连续分布均匀,常氧+E2组与常氧组无明显差异;低氧组大鼠肺动脉管壁均增厚,管腔狭窄,肺小动脉中膜肥厚,中膜内平滑肌细胞增殖明显,低氧+E2组管壁厚度明显低于低氧组(图1)。低氧组较常氧组mPAP、RVHI均明显升高(P均<0.01);常氧+E2组上述指标较常氧组无明显变化(P均>0.05);低氧+E2组上述指标明显低于低氧组(P均<0.01),见表2和图2。

图1 四组大鼠肺组织形态学比较(苏木素伊红染色,×40)
表2 四组大鼠平均肺动脉压和右心室肥厚指数比较(n=8,±s)

表2 四组大鼠平均肺动脉压和右心室肥厚指数比较(n=8,±s)
注:E2:17β-雌二醇。与常氧组比较*P<0.01;与低氧组比较△P<0.01。1 mmHg=0.133 kPa
?
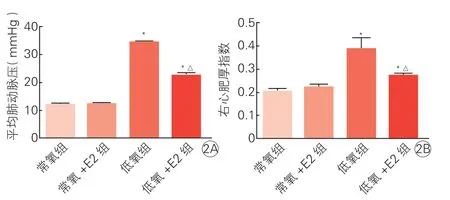
图2 四组大鼠平均肺动脉压(2A)和右心肥厚指数(2B)比较(n=8)
四组大鼠肺动脉中miR-21、PCNA mRNA和蛋白表达水平比较(表3、图3):与常氧组相比,低氧组miR-21及PCNA mRNA、PCNA蛋白的表达均明显上升(P均<0.01),常氧+E2组上述指标无明显变化(P均>0.05);低氧+E2组上述指标虽仍高于常氧组,但已明显低于低氧组(P均<0.01)。
表3 四组大鼠肺动脉中微小核糖核酸-21与增殖细胞核抗体表达水平比较(n=8,±s)

表3 四组大鼠肺动脉中微小核糖核酸-21与增殖细胞核抗体表达水平比较(n=8,±s)
注:E2:17β-雌二醇。与常氧组比较*P<0.01;与低氧组比较△P<0.01
组别 微小核糖核酸-21 增殖细胞核抗体信使核糖核酸 蛋白常氧组 0.0362±0.0010 0.1015±0.0043 0.4906±0.0242常氧+E2组 0.0358±0.0111 0.1025±0.0034 0.5155±0.0284低氧组 0.1206±0.1642* 0.2677±0.0071* 1.7818±0.1719*低氧+E2组 0.0711±0.0073*△ 0.1777±0.0079*△ 1.1688±0.0645*△

图3 四组大鼠肺动脉中微小核糖核酸-21(3A)、增殖细胞核抗体信使核糖核酸(3B)和蛋白表达水平(3C)及蛋白电泳情况(3D)比较(n=8)
三组hPASMC增殖情况比较(表4、图4):细胞培养24 h后,与常氧组相比,低氧组细胞增殖明显(P<0.01),低氧+E2组细胞增殖程度较低氧组明显减轻(P<0.01)。
三组hPASMC中miR-21、PCNA mRNA和蛋白表达水平比较(表4、图5):与常氧组相比,低氧组hPASMC中miR-21、PCNA mRNA和蛋白的表达明显上升(P均<0.01);低氧+E2组hPASMC中上述指标的表达虽仍高于常氧组,但较低氧组已明显下降(P均 <0.01)。
表4 三组hPASMC增殖情况及微小核糖核酸-21、增殖细胞核抗体mRNA和蛋白表达水平比较(±s)
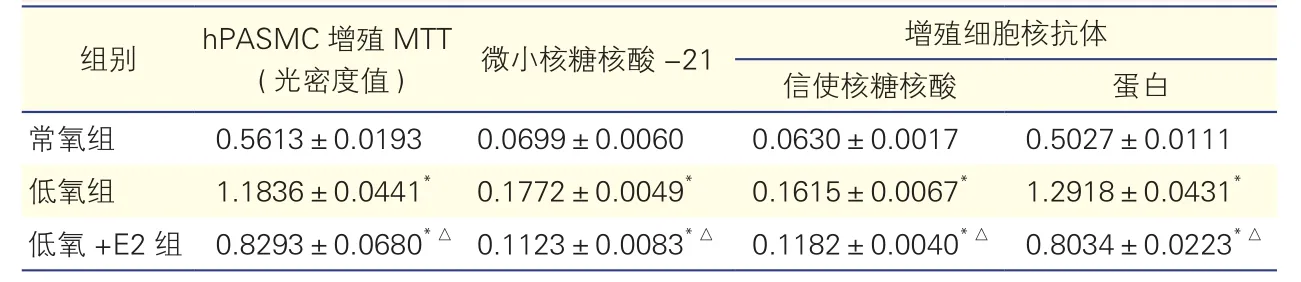
表4 三组hPASMC增殖情况及微小核糖核酸-21、增殖细胞核抗体mRNA和蛋白表达水平比较(±s)
注:hPASMC:人肺动脉平滑肌细胞;mRNA:信使核糖核酸;MTT:四甲基偶氮唑蓝比色法;E2:17β-雌二醇。与常氧组相比*P<0.01;与低氧组相比△P<0.01
组别 hPASMC增殖MTT(光密度值) 微小核糖核酸-21 增殖细胞核抗体信使核糖核酸 蛋白常氧组 0.5613±0.0193 0.0699±0.0060 0.0630±0.0017 0.5027±0.0111
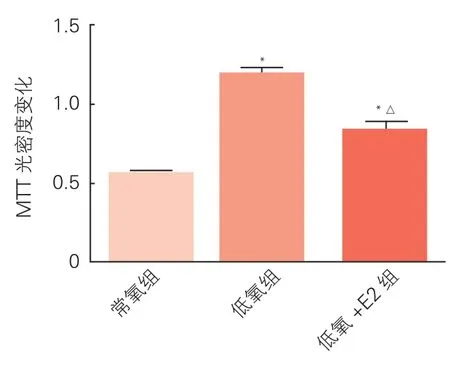
图4 三组hPASMC增殖情况比较
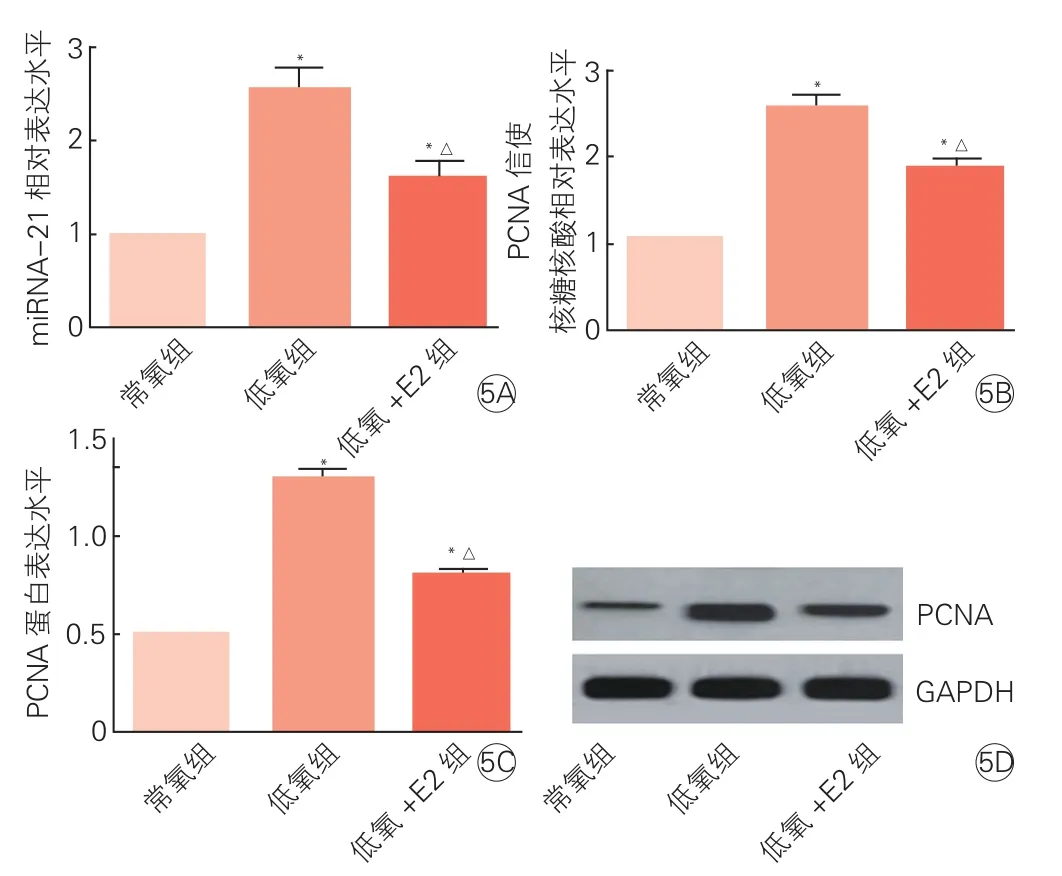
图5 三组hPASMC中微小核糖核酸-21(5A)、增殖细胞核抗体信使核糖核酸(5B)和蛋白表达水平(5C)及蛋白电泳情况(5D)比较
3 讨论
PH以肺血管重构、肺血管阻力进行性升高和右心功能进行性衰竭为特征,病因复杂,致死率高,目前缺乏有效治疗手段[6]。
miRNA是真核细胞中一组高度保守的内源性单链非编码小核苷酸片段,长约22个核苷酸,最早由Lee等[3]在秀丽新小杆线虫中发现。研究表明,多种miRNA与血管再生相关,如miR-17/92、miR-21、miR-143/145等,其中关于miR-21的研究最为广泛。
PH共同的病理改变:肺血管细胞异常增殖和抗凋亡,从而造成肺血管重构。低氧刺激、炎症反应、BMPR2基因突变等多种因素均可导致PH,Parikh等[7]运用生物信息网络方法证明miR-21在三者联系中是一个关键的miRNA,并在PH患者及多种啮齿类动物PH模型肺组织中表达上调。Yang等[8]研究表明在低氧暴露小鼠肺动脉远端miR-21表达升高,无论暴露于低氧前或低氧后,降低miR-21表达均可减弱低氧诱导的PH和肺血管重构,反之亦然。并发现当体外培养的hPASMC过表达miR-21时,可引起PCNA、细胞周期素D1、B淋巴细胞瘤基因(bcl)-xl等增殖相关蛋白的表达上调。本研究成功建立HPH大鼠模型,并在低氧暴露大鼠的肺动脉中观察到miR-21及与增殖相关的PCNA的表达上调,虽然动物模型有差异,但实验结果与Yang等[8]的研究一致,并进一步通过体外细胞学实验验证了miR-21的上调与PASMC增殖相关。Sarkar等[4]发现,hPASMC在低氧(3%的氧气)环境培养6 h后,miR-21表达增加了3倍,培养24 h后,只增加2倍。本研究体外细胞培养24 h后miR-21表达增加了大约2倍,动物实验中慢性低氧暴露使得肺动脉中miR-21表达上调达3倍多。对于动物水平与细胞水平表达不一致的现象我们考虑有以下几点原因:(1)低氧处理时间不同所致。我们在细胞水平采取的是低氧24 h处理后检测相应指标,而如前所述,随着低氧时间的延长miR-21表达会较峰值有所下降。(2)其他细胞成分中miR-21的表达增加参与其中。肺动脉血管壁分3层:含有成纤维细胞的外膜,以PASMC为主的中膜,只有一层内皮细胞的内膜。且目前认为miRNA参与肺血管收缩和重构,既调控PASMC增殖、凋亡及表型转化,又在血管内皮细胞增殖、凋亡起重要作用[9]。故推测这种组织与细胞水平增长程度不一致情况,可能与低氧同时上调了肺动脉内皮细胞中的miR-21表达有关。相关研究表明,miR-21在肺动脉内皮细胞(PAEC)中作用机制复杂[7,10,11],但无论在内皮细胞还是平滑肌细胞,低氧条件下miR-21表达均上调,这便能使我们的实验结果得到合理解释。
前期研究证明,慢性低氧导致去势的大鼠形成更为严重的PH、肺动脉重构及右心室的肥厚;进一步研究证实,E2干预可以缓解慢性低氧性肺动脉压力的升高及肺血管重构,在HPH中起保护作用[12-14]。此次研究结果与前期研究结果一致,需要说明的是,在常氧条件下,无论是单纯去势组还是去势后又补充E2组,两组无论是肺血管形态学、右心室肥厚程度还是mPAP均处于较一致水平,说明E2在低氧条件下才表现出显著的肺血管保护作用。
从动物实验水平上,已经证实miR-21参与了低氧诱导的肺血管重构,并且多项研究表明其参与hPASMC增殖,那么E2对肺血管的保护作用是否通过调节miR-21来实现的呢?为了验证这一点,我们在动物水平及细胞水平均添加了E2干预因素,结果表明E2对肺血管的保护作用可能是通过下调miR-21表达从而抑制PASMC增殖实现的。E2与miR-21之间存在相关性,E2通过抑制miR-21表达发挥作用早在2009年就被Wickramasinghe等[15]证实,他们的研究显示,在乳腺癌MCF-7细胞中,E2通过激活的雌激素受体(ER)来抑制miR-21的表达,同时miR-21的靶基因表达上调,其机制可能是雌激素同源受体ER α和ER β通过与miR-21启动子中具有雌激素效应元件的配体结合来实现的。后来Selcuklu等[16]也报道了此关系,但两者研究的均是乳腺癌细胞,雌激素直接通过结合配体依赖性转录因子ER α和ER β,间接激活与细胞膜相关的ER来调节基因表达,从而不断激活细胞内信号级导致基因的异常表达[17]。那么在肺血管细胞中E2与miR-21之间是否也通过ER α、ER β或者GPR30受体来实现相互作用,抑或存在其他信号通路?另外鉴于miR-21同时存在于PASMC及PAEC中,该实验未进一步体外培养PAEC检测相应指标变化,故不能确定E2是否也调节了PAEC中miR-21的表达,这一系列问题尚需进一步研究证实。
猜你喜欢
山东第一医科大学(山东省医学科学院)学报(2022年8期)2022-12-07
昆明医科大学学报(2021年6期)2021-07-31
天津医科大学学报(2021年3期)2021-07-21
现代临床医学(2021年2期)2021-03-29
心肺血管病杂志(2020年5期)2021-01-14
心肺血管病杂志(2020年5期)2021-01-14
中国特种设备安全(2019年4期)2019-05-20
西江月(2018年5期)2018-06-08
人大建设(2017年4期)2017-07-21
海峡姐妹(2017年5期)2017-06-05
