
Pilot study on the effects of elevated air temperature and CO2 on artificially defoliated silver birch saplings
2018-11-06 06:44:06ValdaAraminienIvetaVarnagirytKabainskienVidasStaknas
Journal of Forestry Research 2018年6期
Valda Araminienė•Iveta Varnagirytė-Kabašinskienė•Vidas Stakėnas
Abstract The impacts of elevated temperature and CO2on young silver birch(Betula pendula Roth)saplings after 0,25,50 or 75% artificial defoliation were assessed by measuring plant height and dry mass of aboveground compartments and roots and various morphological and physiological variables.Defoliation either increased or decreased plant growth depending on the severity of damage and the climatic treatment.At21°C and 400 mg L-1CO2,defoliated plants were not able to compensate for the lost foliage,but growth compensation and adaptation to the changed conditions were greater;growth of young defoliated silver birch saplings increased,which led to increased height and a tendency to enhance final aboveground and root biomass and leaf nitrogen and carbon content compared to the nondefoliated controls.Nevertheless,the short-term effect of the different climatic conditions did not result in a significant overgrowth of defoliated plants.A slight increase in temperature and CO2 were the most acceptable conditions for defoliated plants;however,a 4°C increase with correspondingly higher CO2 was more stressful as shown by less growth in height and biomass allocation to leaves,stems and roots.The findings from the pilot experiment are more applicable to young birch trees,but stress on young trees may be reflected in future tree growth.
Keywords Biomass·Betula pendula·CO2concentration·Elevated temperature·Photosynthesis
Introduction
Over the past century as interest in global warming has increased,tree biological responses to climate change have received considerable attention.Higher atmospheric temperature and CO2concentration impact forest tree species,climatic changes can induce specific reactions and alter forest dynamics such as tree growth,mortality and regeneration.The question is whether forest management needs to be modified as a result of the changing climate(Lawrence and Marzano 2014).
Recent studies have already illustrated that the growth of damaged trees can be improved by climate change(Huttunen et al.2007,2013;Reyer et al.2014).However,warmer seasons can also increase the risk of widespread foliage damage caused by insect herbivores(Volney and Fleming 2000;Dale et al.2001;Wolf et al.2008),in parallel functioning with other environmental factors.When temperature increases,insects may become more abundant,and the spread of new insect species is also expected.From one side,the predicted findings show that the damage risk for certain tree species will increase,i.e.,the trees can be seriously damaged,and the condition of forest stands deteriorate,leading to growth decline.Lindner et al.(2010)emphasised that long-lived trees do not easily adapt to environmental changes.Other studies have documented that an increase in atmospheric CO2and temperature stimulates photosynthetic activity(Long 1991;Lee et al.1998)and,consequently,enhances height and stem growth of deciduous species,including silver birch(Huttunen et al.2007).A number of researchers have reported that not only height and stem growth respond to changing environment but,initially,the higher concentrations of phenolic compounds in the tree leaves can also be produced as a reaction to the increased CO2(Kuokkanen et al.2001).More often,the higher temperature induces an increase in leaf size and number of newly burst leaves(Gunn and Farrar 1999).However,the opposite response was obtained under the current climatic conditions:foliage reduction caused by insect feeding reduced the growth of new leaves(Dormann et al.2000;Mattson et al.2004).It has been also suggested that the ability to tolerate the insect damage on silver birch saplings under the current climate depends on individual tree development:taller trees were more resistant than shaded trees(Prittinen et al.2003).Such examples outline the other side of tree responses to changing climate and leads to a discussion raised two decades ago by Ayres(1993),who reported that climate change(increased temperature)improves the ability to tolerate insect damage.
Huttunen et al.(2007)indicated that silver birch is tolerant to herbivore damage and more often is able to overcompensate in height and biomass growth.This compensational tree growth or response has been widely discussed by several authors(Agrawal 2000;Anttonen et al.2002;Huttunen et al.2007,2012),and could be used as a tool to interpret tree response under different climatic conditions.
Birch stands comprise more than a half of the deciduous forest area in Lithuania(Ministry of Environment/State Forest Service 2015),where the climate is predicted to become even more favourable to deciduous species,including birch,during the coming decades(Ozolinčius et al.2014).The vegetation,including woody plants,more often are destroyed by the short-term summer heat waves that are recorded during July–August in the territory.To simulate the situation for a short period,we elevated the growth temperature and CO2for plants with and without artificial defoliation.Because the evaluation of the effect of changes in climatic conditions on trees is quite challenging,we used young tree saplings for these simulations as an initial tool to interpret the adaptive capacity of the trees at a local scale.To that end,we initiated a pilot study to determine the short-term effects of artificially elevated air temperature and atmospheric CO2on young silver birch(Betula pendula Roth)saplings.
Materials and methods
Planting material,treatments and growing conditions
The planting material originated from a naturally regenerated silver birch(Betula pendula Roth)stand in the southwestern Lithuania(54°51′N,24°03′E).The birch saplings from this nursery are used for reforestation.One-year-old birch saplings were replanted into individual 4-L plastic base-perforated pots containing sand and neutralised peat mixture(one part of sand and five parts of peat)in early April 2015.The substrate contained the optimum nutrition for silver birch growth(pHKCl5.0–5.5;about 50 mg L-1of each N,P and K;and 30 mg L-1of Mg),which was determined by the Agrochemical Laboratory of the Lithuanian Research Centre for Agriculture and Forestry.No additional fertilizers were added during the study.The saplings were watered as needed during the experiment.
The potted saplings were grown in an open field for 2.5 months until the start of the simulation experiment.In June 2015,the canopy of the saplings was defoliated(25,50 or 75%of leaf area)using scissors,cutting each leaf perpendicular to the midrib near its base.Nondefoliated saplings served as the control(0%defoliation).
After a few days,the saplings were placed in three closed walk-in indoor chambers(2.5×2×2 m)with controlled climate for 6 weeks to simulate present atmospheric temperature and CO2concentration and projected elevated temperatures and CO2in the laboratory of Vytautas Magnus University,Lithuania.The photoperiod was 14 h light/10 h dark,air humidity was 75%,and photosynthetic photon flux density was about 200 μmol m-2s-1in all chambers.The light was provided by six natural fluorescent lamps(Philips,Waterproof OPK Natural Daylight LF80 Wattage 2×58 W/TL-D 58 W)and one high pressure sodium lamp(Philips MASTER GreenPower CG T 600 W).
The potted saplings were exposed to the following temperatures and CO2concentration: first chamber,21°C/14°C day/night,400 mg L-1CO2(present conditions):second chamber 23 °C/16 °C,550 mg L-1CO2;third chamber,25 °C/18 °C,750 mg L-1CO2.The photoperiod,humidity and photosynthetic photon flux density were automatically adjusted,temperature and CO2concentration were controlled by the PC-based Environmental Control System(computer software IGSS 9-13175).Despite uniform conditions within each chamber,plants were moved to different locations in the chamber periodically to avoid possible variations in the growing environment.There were six saplings per defoliation treatment(0,25,50 and 75%defoliation)in each chamber,totalling 72 saplings.
Measurements
The height of potted birch saplings was measured twice:at the beginning of the climate simulation experiment(30 June)and at the end,i.e.,after 6 weeks of growth in the chambers(mid August).
The photosynthetic rate(μmol CO2m-2s-1)of six of the remaining leaves per treatment was measured with a photosynthetic system LI-COR 6400(LI-COR,Lincoln,NE,USA)on August 10(after 6 weeks in the chamber)between 10:00 and 15:00 h.In each treatment,the mature and visually healthy leaves were measured in the middle part of the canopy.The transpiration rate(mmol H2-O m-2s-1)was also estimated.
Chlorophyll a and b and carotenoids in leaves were measured spectrophotometrically (UV/VIS Spectophotometer,PG INSTRUMENTS LTD)method after standard ethanol extraction.The leaves were sampled from the nondefoliated control,25-,50-and 75%-defoliated plants after 6 weeks in the different climatic chambers.Three leaves from each sapling were sampled from the middle crown,yielding 18 leaves per treatment.The leaf samples were combined into three samples.Chlorophyll a,b and total chlorophyll(a+b)concentrations(mg L-1)were calculated by the following formulas:

where D is the sample absorbance at 662 or 644 nm.
The chlorophyll content of leaves was expressed in milligrams per gram of fresh mass of leaves:

where A is the chlorophyll content of the fresh mass of leaves(mg g-1),C is the chlorophyll concentration(mg L-1),V is the volume of chlorophyll extract(mL)and P is the mass of plant material(g).
The total content of phenolic compounds was measured spectrophotometrically according to the method described by Slinkard and Singleton(1977)and Makkar et al.(2007).Two leaves from each sapling were sampled from the middle crown,yielding 12 leaves per treatment.The leaf samples were combined into three samples.
Six weeks after birch saplings growing in the climatic chambers,in the middle of August,above-and below ground biomass measurements were conducted.For the determination of total dry mass,samples were removed from the following tree compartments:leaves,living and dead shoots,branches,stems and roots.Roots were rinsed free of soil and oven-dried to determine the total belowground biomass.The samples were transported to the laboratory in paper bags and stored in a ventilated room.The oven drying took place within a few days after sampling.The removed biomass from each treatment,as well as the final live biomass,was collected and oven-dried to a constant mass at 60°C.
The leaf areas of three leaves,taken randomly from each plant per each treatment,were estimated using a scanner and leaf area analysis software WinFOLIA 2004a(Regent Instruments,Quebec City,Quebec,Canada).The diameter of air-dried main stems was measured 2 cm above the stump base for each individual plant using an electronic digital calliper.The length of the roots of each plant was measured from the root base to the bottom,straightening the root before measurement.
In the leaf samples,total N was analysed using the Kjeldahl method(ISO 11261:1995),and organic carbon(C)was determined by dry combustion at 900°C with a ECS 4010 CNS analyzer(ISO 10694:1995).
Calculations and statistical analysis
Leaf and total mass was calculated by summing the dry mass at harvest plus the leaf mass removed during the defoliation events.The total mass of each sapling was calculated by the summation of the leaf,shoot,stem and root dry masses,in grams.The process of growth compensation,that is whether birch saplings partially or fully compensated for the removal of leaf biomass,was evaluated when the height increment,photosynthetic rate and final biomass of defoliated saplings were compared to those of the control saplings during the 6-week period.
The normality of the variables was checked using Lilliefors and Kolmogorov–Smirnov tests.The Kruskal–Wallis analysis of variance(ANOVA)test was used to ascertain the significant differences in dry mass between the different treatments.This test is used as a nonparametric alternative to between-groups one-way ANOVA in cases when the data are notnormally distributed.Throughout the study,the means are presented with the standard error of the mean(±SE).Statistical analyses were conducted using the Statistica 7.0 software at p<0.05 in all cases.
Results
Morphometrical variables of silver birch saplings
During the limited 6-week period,the growth rate of the nondefoliated control silver birch saplings was very similar and did not vary among the different environmental conditions.The height increment of the plants varied between 10.6 and 13.7 cm,irrespective of climatic conditions(Fig.1).The slightly defoliated saplings grew more in the warmer environment with higher CO2concentration.The height increment of birch saplings grown at 23°C with 550 mg L-1CO2and at 25°C with 750 mg L-1CO2increased by 1.7–1.9 times compared to those at the current conditions(21°C day,400 mg L-1CO2).The height increment of 75%defoliated saplings was 1.5 times higher in the warmest environment with 750 mg L-1CO2compared to the milder conditions.
Defoliation either increased or slowed growth in height depending on the severity of damage and the climatic treatment(Table 1).The height growth was reduced in response to 50 and 75%removal of total leaf area in some climatic treatments.The 25%defoliation increased height growth compared to the nondefoliated controls in the saplings affected by elevated temperature and CO2.The plants in these treatments were the tallest(about 71 cm),7%taller than the nondefoliated plants,and over 15%taller than the plants grown at 21°C and 400 mg L-1CO2.Under elevated temperature(25°/18 °C)and 750 mg L-1CO2,a 25%defoliation caused a 6%increase in stem diameter over the nondefoliated controls(Table 1).
Throughout the growing season,the largest longest root(29 cm)was found on the 25%defoliated plants compared to the nondefoliated controls treated with the elevated temperature and CO2.Defoliation affected the total leaf area at harvest(Table 1).The effect of foliage loss was dependent both on climatic treatment and the magnitude of foliage removal.Compared to the nondefoliated controls,the total leaf area increased in the defoliated plants.At the lowest temperature and CO2,the increase ranged between 17 and over 80%depending on the severity of damage.Under the elevated temperature and CO2,the total leaf area increased nonlinearly with the increase in defoliation severity.In most cases,the leaf area increase ranged between 44 and 67%compared to the nondefoliated controls.A 50%defoliation,on the other hand,increased the leaf area by two times compared with the nondefoliated controls at 23°C and 550 mg L-1CO2.
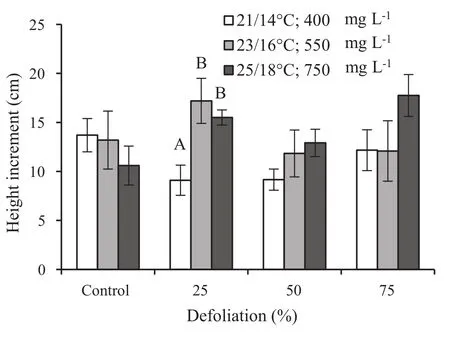
Fig.1 Height increment of silver birch saplings after different levels of defoliation and 6 weeks at different temperatures and CO2levels.Different letters indicate significant difference between treatments
Above-and belowground biomass and biomass allocation
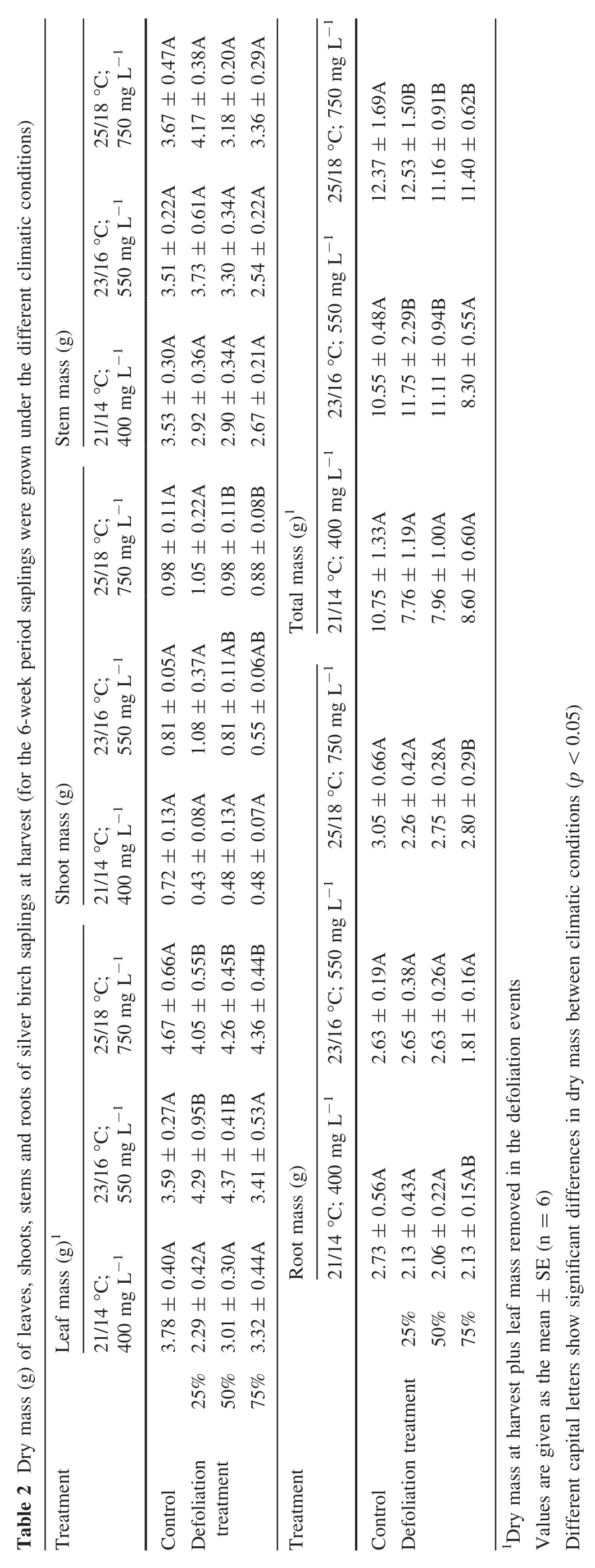
No significant differences were found between final leaf mass(dry leaf mass at harvest plus leaf mass lost in defoliation)of defoliated and nondefoliated plants at the different temperatures and CO2concentrations(Table 2).On a per plant basis,leaf mass was 2.3–3.8 g at 21 °C and 400 mg L-1CO2;3.4–4.3 g at 23 °C and 550 mg L-1CO2and 4.1–4.3 g at 25 °C and 750 mg L-1CO2(Table 2).
The results for the other aboveground biomass compartments—stems and shoots—revealed no relation to the different climatic conditions(Table 2).The lowest dry mass for shoots was determined for the plants at 21°C and 400 mg L-1CO2.Elevated temperatures and CO2levels caused more shoot growth;however,there were no differences between defoliated plants and the nondefoliated controls.
The 75%-defoliated plants under the conditions when temperature was elevated by 4 °C (at25 °C)and 750 mg L-1CO2produced the most aboveground mass compared to the lower temperature and CO2.At both elevated temperatures and CO2levels,plants produced 1.5–1.7 times more aboveground mass compared to the plants grown under the milder climatic conditions.
Elevated temperature and CO2intensified plant growth.Defoliated plants produced similar aboveground biomass at harvest compared to the nondefoliated controls.However,foliage loss did not significantly affect the root mass at harvest,i.e.no significant differences showed complete root growth compensation of 25–75%defoliated plants compared to the nondefoliated controls.
However,specific combinations of climatic treatments and foliage removal caused some variations in the general trend of leaf biomass(Fig.2A).For example,21°C with 400 mg L-1CO2caused a decreasing trend in leaf production,which could be defined as partial compensation or under compensation.However,plants with more defoliation differed less from the nondefoliated control.When plants lost more foliage at the initial stage,they were able to stimulate their growth and produce more biomass.At 23°C and 550 mg L-1CO2,the trend was the opposite.Compared to nondefoliated controls,removal of 25 and 50%of the total leaf area increased leaf production at harvest,but with the removal of 75%of foliage,leaf production remained at the nondefoliated control level.However,25°C with 750 mg L-1CO2caused a slight decrease in final leaf production without clear differences between defoliations.
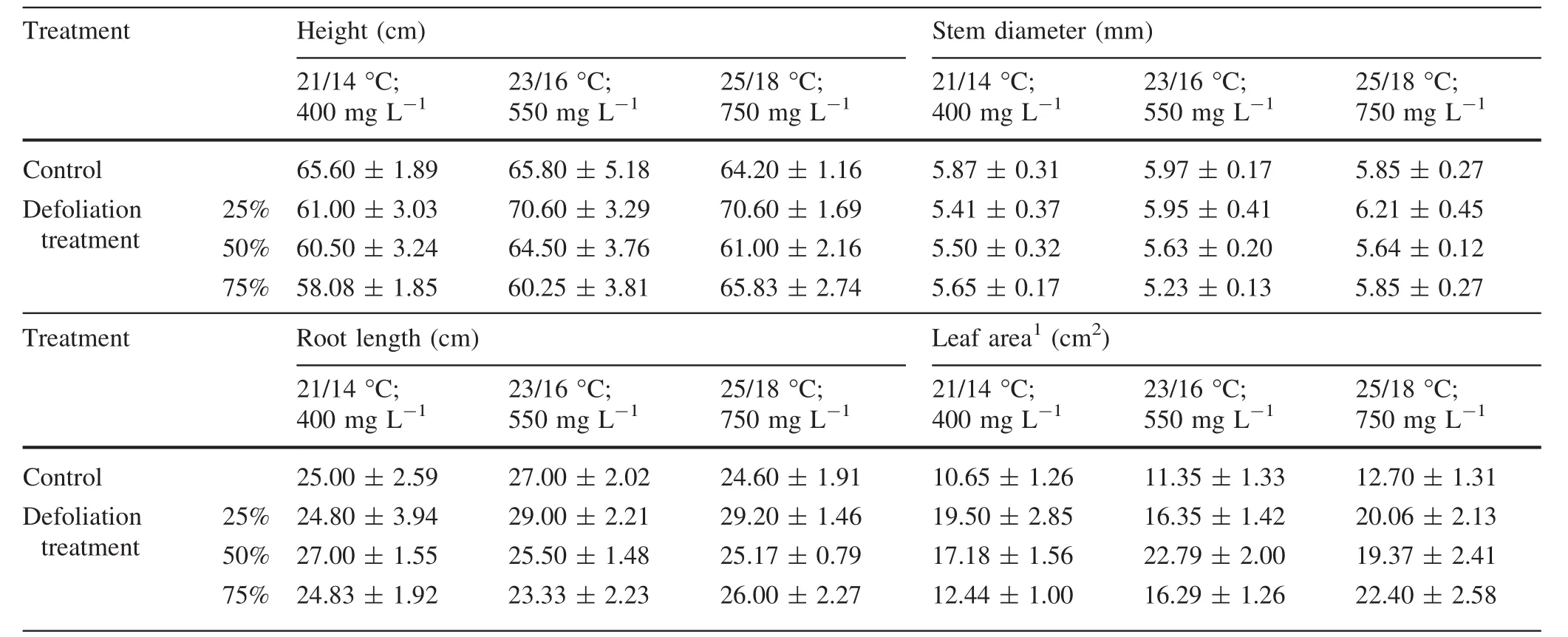
Table 1 Some morphometrical parameters of silver birch saplings at harvest under the different climatic conditions(temperature day/night;CO2 concentration)
The results of other aboveground biomass compartments—stems and shoots—revealed no dependence on different climatic conditions.The lowest dry mass of shoots was determined under the 21°C temperature and 400 mg L-1CO2condition,i.e.the estimated values were in the range of 0.4–0.7 g per plant.While both elevated temperatures with corresponding CO2concentrations caused more intensive shoot growth followed by 1.5–1.8 times higher final shoot mass but no differences between defoliated and nondefoliated controls.
The elevated temperature slightly increased stem biomass at harvest but did not result in an overgrowth of defoliated plants compared to the nondefoliated controls.Defoliations caused a decreasing trend in stem biomass under the tested climatic conditions,except for the 25%defoliation,which resulted in stem biomass increase at elevated temperatures and CO2(Fig.2B).
Elevated temperature and CO2intensified plant growth,especially for the aboveground plant compartments.Defoliated plants,at elevated temperatures and CO2,increased their aboveground biomass by 1.3–1.6 times compared to the lower temperature and CO2.However,defoliated plants produced similar aboveground biomass at harvest compared to the nondefoliated controls in each climatic treatment.The response trend of aboveground biomass of defoliated plants compared to the nondefoliated controls well reflected the trend for leaf biomass at all climatic treatments and defoliations(Fig.2A,C).At 21°C and 400 mg L-1CO2,25–75%defoliations increased the aboveground biomass compared to the nondefoliated controls(Fig.2C).However,when air temperature was elevated to 23°C and CO2up to 550 mg L-1,the aboveground biomass changes from the nondefoliated controls depended on the defoliation treatment.Under the latter climatic treatment,the 25 and 50%defoliation increased the aboveground biomass(defined as the compensation),but 75%defoliation decreased the aboveground biomass(defined as undercompensation).The defoliated plants were able to reach the aboveground biomass of nondefoliated controls at temperatures to 25°C with 750 mg L-1CO2.
Foliage loss did not significantly affect root biomass at harvest.Final root biomass was estimated as 2–3 g per plant but did not depend on climatic treatment and defoliations.Evaluating the response trend,defoliations resulted in less root biomass compared to the nondefoliated controls t 21°C and 400 mg L-1CO2(Fig.2D).The root biomass of the 25 and 50%defoliated plants remained at the nondefoliated controls level when the temperature was elevated to 23°C with the corresponding CO2concentration.
The different climatic conditions did not result in a significant overgrowth,but the defoliated plants tended to compensate for the lost foliage.The largest impact on the final biomass production or full compensation was determined for the plants grown at the highest temperature(current atmospheric temperature plus 4°C) and 775 mg L-1CO2.
?
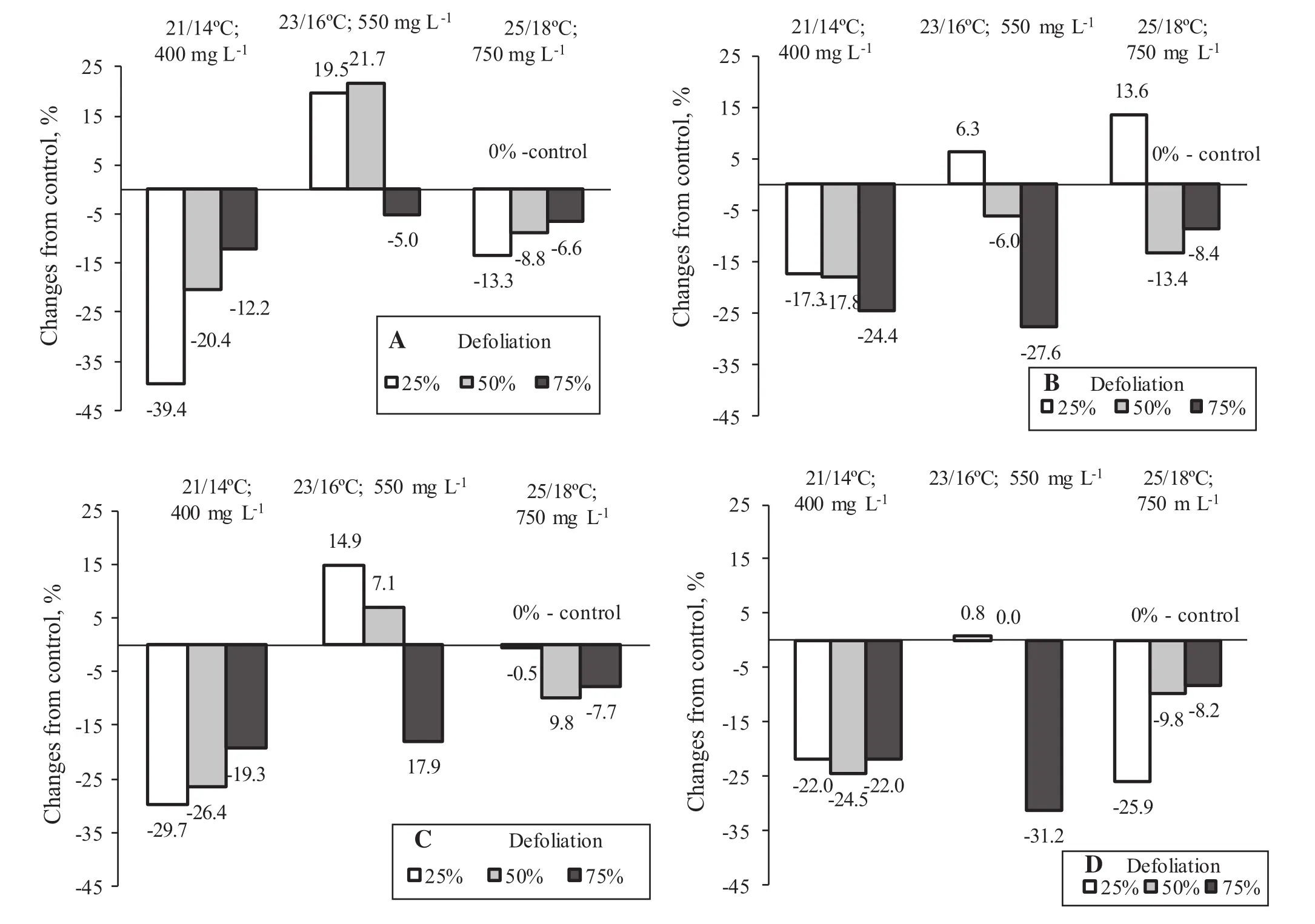
Fig.2 Percentage change in leaf(A),stem(B),aboveground(C)and root(D)dry mass from control(0%is equal to control)at harvest.Values equal or close to the control mean full or equal compensation,obviously higher values indicate overcompensation,obviously lower values,undercompensation or no compensation
Biomass allocation to stems tended to increase,but allocation to leaves decreased in plants with 25%defoliation compared to the nondefoliated controls at 21 and 25°C with corresponding CO2levels(Fig.3).However,the opposite pattern was observed at23°C and 550 mg L-1CO2:defoliated plants allocated more biomass to leaves.For example,the 75%defoliation increased the biomass allocation to leaves by 7%.
Physiological,biochemical and chemical leaf properties
When photosynthesis and transpiration rate were measured in plants grown at 21 and 23°C with 400 and 550 mg L-1CO2at different levels of defoliation,the 25,50 and 75%defoliation treatments increased the leaf photosynthetic rate over that in the undefoliated controls(Table 3).Leaf transpiration rate was reduced for 50–75%defoliated plants compared to the nondefoliated controls at 21°C and 400 mg L-1CO2.However,the transpiration rate slightly increased under the elevated temperatures.
Defoliation led to a decrease in chlorophyll a and b concentrations compared to the nondefoliated controls at 21°C and 400 mg L-1CO2(Table 4);the decrease depended on the severity of the damage.For example,concentrations with the 25–50%defoliation were reduced by 1.5–1.6 times compared with the nondefoliated plants,but 75%defoliation only reduced it by 1.2 times.The defoliation caused an increase in the ration of chlorophyll a to b compared to the nondefoliated controls,except for the 75%-defoliated plants.
The elevated temperature up to 25°C at 400 mg L-1CO2caused an increase in the chlorophyll a and b concentrations but a decrease in their ratio compared to levels in the nondefoliated control(Table 4).At 23°C and 550 mg L-1CO2,chlorophyll levels in the defoliated plants and nondefoliated controls did not differ significantly.Photosynthetic rate did not differ for the defoliated and nondefoliated plants under the 25°C and 750 mg L-1CO2.
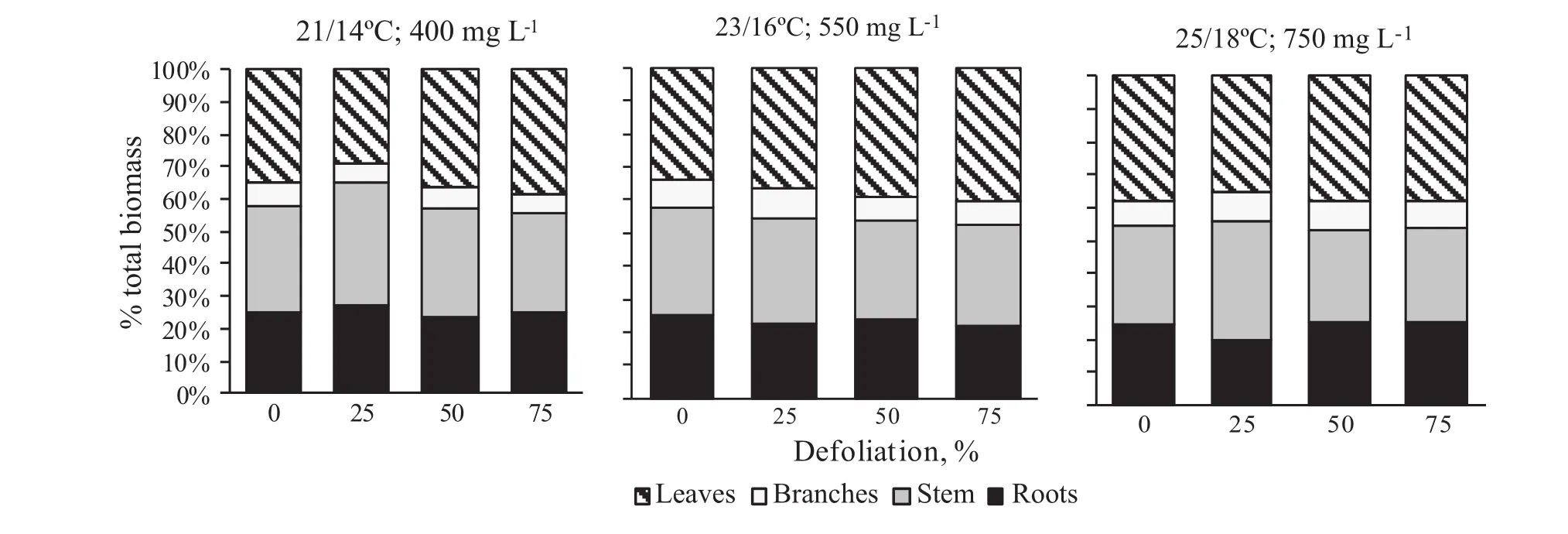
Fig.3 Biomass allocation to stem,leaves,and branches of silver birch saplings after different levels of defoliation and 6 weeks at different temperatures and CO2levels
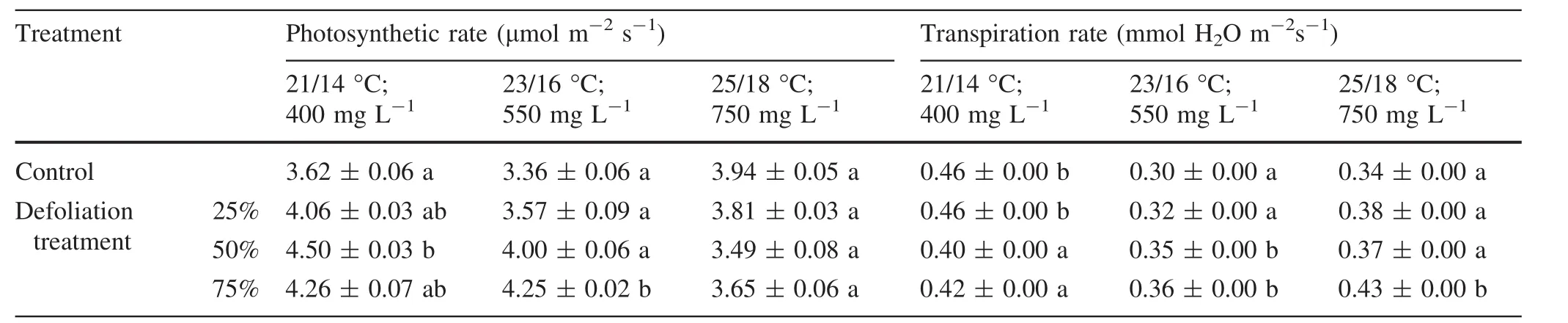
Table 3 Some physiological parameters in the silver birch leaves under the different climatic conditions(temperature day/night;CO2 concentration)
In the case of the nondefoliated controls,the elevated temperature caused the slight decrease of total phenolics compared to the plants grown at 21°C and 400 mg L-1CO2(Table 5).The defoliations caused either an increase or no change in total phenolics compared to the nondefoliated controls at the elevated temperature and higher CO2levels than at the milder conditions(21°C,400 mg L-1CO2).
The leaf nitrogen and carbon contents of nondefoliated saplings was 1.3 times higher at elevated temperatures up to 25°C and 750 mg L-1CO2compared with the milder conditions(Table 6).Compared to the nondefoliated controls,the 25 and 50%defoliations tended to decrease leaf nitrogen and carbon.The opposite patterns were recorded for the saplings grown at 23°C with 550 mg L-1CO2:the higher defoliation tended to increase the nitrogen and carbon levels compared to the nondefoliated controls.
Discussion
The results of this study indicate that in general the higher atmospheric temperature in combination with increased CO2tended to intensify the growth of silver birch saplings,which subsequently led to higher final biomass.The assessment of air temperature and carbon dioxide concentration impact on silver birch sapling growth showed 22%higher dry mass in saplings with 25–75%defoliation at the elevated CO2concentration up to 550 mg L-1and increased air temperature up to 23°C.Meanwhile,when CO2concentration was increased to 750 mg L-1and air temperature increased to 25°C,the total biomass of defoliated saplings was 30%higher than in the controls.However,the plants at 21°C and 400 mg L-1CO2were not able to fully compensate for the lost foliage,which could be interpreted as undercompensation.The capacity of defoliated plants for compensatory growth depends on a variety of combinations between environmental conditions and internal plant peculiarities(Mutikainen et al.2000;Anttonen et al. 2002; Huttunen et al.2007,2008,2012,2013).Huttunen et al.(2007)highlighted that overcompensation of young and light-demanding birch is important for species’competitive ability,especially if the plants were damaged by herbivores.
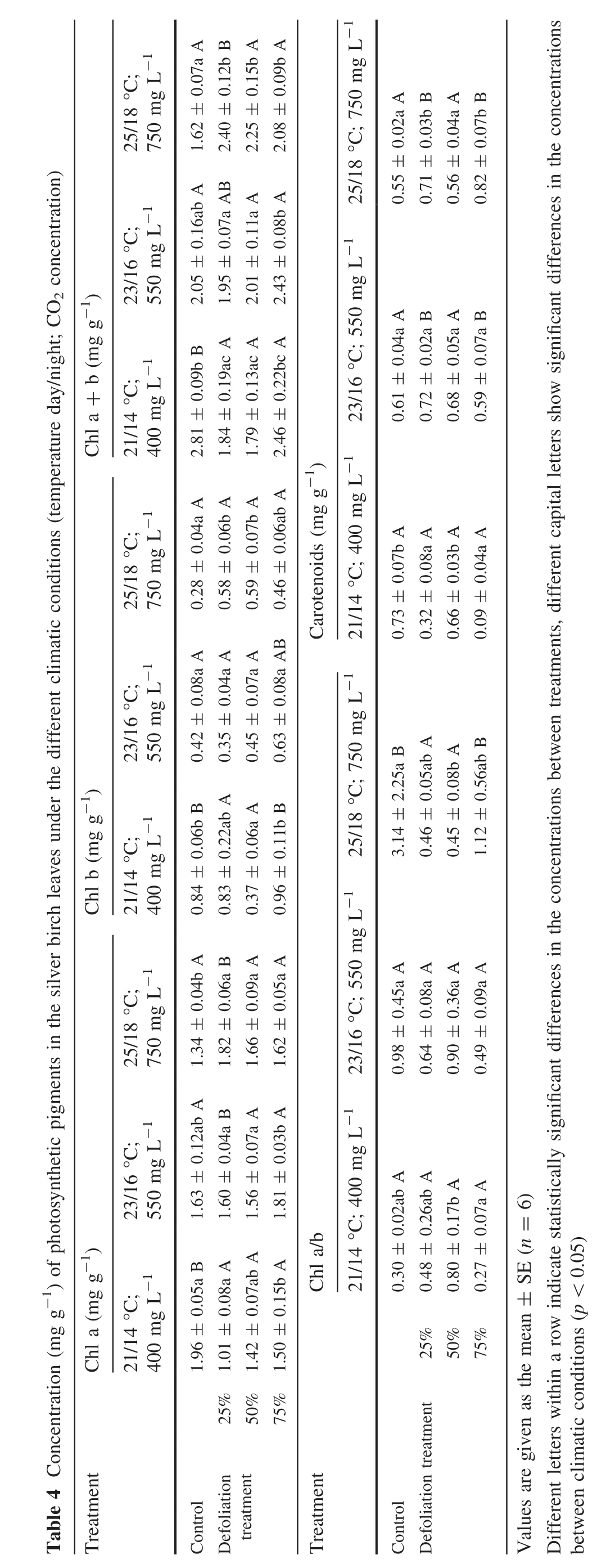
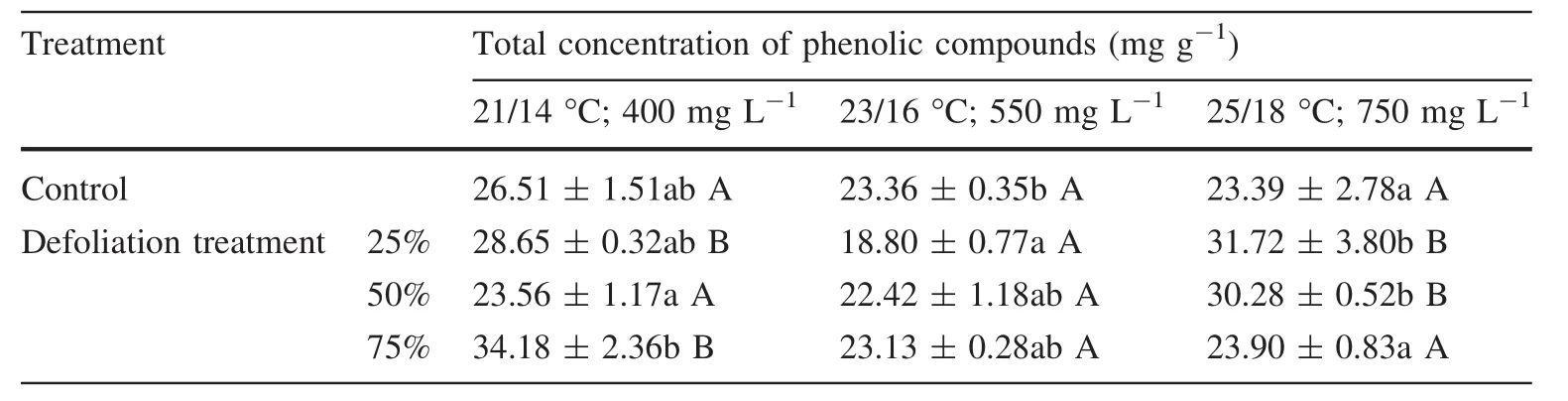
Table 5 Total concentration of phenolic compounds(mg g-1)in the silver birch leaves under the different climatic conditions(temperature day/night;CO2 concentration)
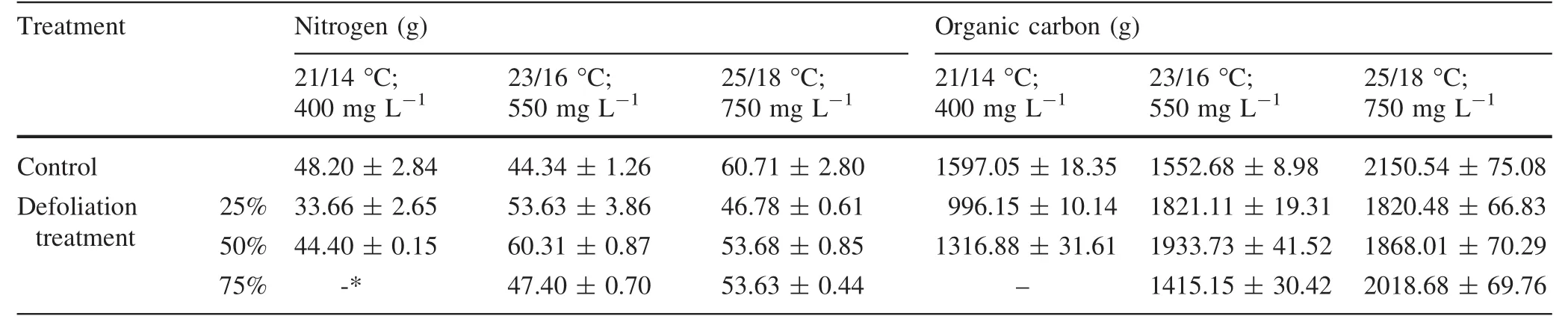
Table 6 Nitrogen and carbon content in the silver birch leaves(grams per leaf)
Both negative and positive plant response to defoliations have been reported in the literature.Our findings are in accord with the recent studies indicating that not only growth in height was affected by the defoliations,but also the dry mass increased in defoliated plants.All the main factors affected the dry mass of shoots and roots.The defoliation either increased or decreased dry mass depending on the severity of the defoliation.The magnitude of the effect was greater for aboveground biomass compartments than for root biomass.Our results showed that birch tended to be able to compensate for the loss of foliage during the first growing season.
In a prior study,only complete(100%)defoliation affected biomass allocation to stem,branches and leaves(Anttonen et al.2002),and 1 year after the defoliation,biomass allocation to the stem increased in defoliated saplings compared to controls.On the whole,our findings indicated more sensitive and faster plant response;however,in some cases,this response may have resulted from differences in the growing environment.
The silver birch saplings,with the different defoliation levels and grown at the elevated temperature and higher CO2,were able to adapt to the changed conditions,and their photosynthetic rates did not differ much compared to those in the milder climatic conditions.Similar photosynthetic levels were obtained for the control and defoliated saplings that were grown at the 750 mg L-1CO2and 25°C,but they were more intensive at lower temperature and CO2.Elevated air temperature and higher CO2concentration induced higher chlorophyll a and b ratios in the control saplings,while the climatic conditions did not significantly affect the chlorophyll a/b in defoliated saplings.Photosynthesis is regulated by different factors such as leaf temperature and CO2availability,which were included in the photosynthesis model by Karlberg et al.(2006).Elevated atmospheric CO2tends to increase the rate of photosynthetic carbon fixation by leaves.For example,Ainsworth and Rogers(2007)reported that leaf photosynthetic rates in plants grown at elevated CO2(475–600 mg L-1)increased by 40%.The defoliations combined with elevated temperature and CO2in the present study enhanced the photosynthetic rate.The results of this study con firm that partial defoliation tended to increase the photosynthetic rate of leaves and compensated for the loss of foliage as shown in earlier studies of Bassman and Dickman(1982),Reichenbacker et al.(1996)and Hart et al.(2000).
The decrease in N concentration in aboveground biomass as a response to elevated CO2has been widely studied(Ainsworth and Long 2005;Taub and Wang 2008),but the results among these studies have also varied.For example,Yin(2002)found that the effect of elevated CO2was greatest for deciduous species,which had more pronounced N decreases under elevated CO2when temperatures were also high.Other data showed that foliage regrowth on trees after defoliations was accompanied by increased leaf nitrogen and photosynthesis(Heichel and Turner 1983).Very little was found in the literature on plant organic carbon content at different defoliation levels and temperatures.We found slightly more organic carbon in the leaf biomass in silver birch saplings at 25°C and 750 mg L-1CO2,while organic carbon was significantly higher than in birch saplings grown at 21°C with 400 mg L-1CO2.This ability to accumulate higher organic carbon at elevated atmospheric temperature and CO2could help mitigate the negative effects of climate change.
The obtained data did not show a significantly positive impact of changed climatic conditions on young birch saplings growth,but the plants tended to increase leaf biomass,photosynthetic rate and organic carbon content when the ambient temperature was higher than 21°C,even for a short time.Thus,we would expect that young,healthy,defoliated trees,growing naturally,would respond similarly if air temperature increased by 2 or 4°C.However,these results cannot be directly applied to older trees because their growth potential,physiological processes,root system development and nutrient reserves differ from saplings.Older trees also tend to have higher adaptability to environmental factors,but any stress incurred at a young age(stand establishment stage)may be reflected in later growth processes in older stands.
Since tree growth responded to several factors(damage intensity,air temperature and CO2concentration)simultaneously,it was difficult to distinguish the predisposing factor.We recognise that our data were obtained from a limited experiment and may therefore not necessarily represent the behaviour of similar species throughout the region.Also,the experimental design of this study does not allow statistical inferences about the influences of individual environmental disturbances on birch trees.Nevertheless,our results start to fill in gaps in our knowledge about potential responses of healthy and defoliated silver birch trees to future global changes.
Conclusions
The simulation of short-term warmer weather conditions and higher CO2concentration led to more intensive growth of silver birch saplings,they tended to produce more biomass.The 25–75%-defoliated saplings produced 22–30%more dry mass under the elevated air temperature and CO2compared with the controls.The short-term elevated air temperature and higher CO2concentration did not significantly increase the photosynthetic rate but induced a higher ratio of chlorophyll a to b in the control seedlings.The climatic conditions did not significantly change either photosynthetic rate or the concentration of chlorophyll a and b in defoliated seedlings compared to the nondefoliated controls.The leaf nitrogen and carbon content in the nondefoliated saplings was 1.3 times higher at the elevated temperature up to 25°C and 750 mg L-1CO2compared to the milder conditions.On the basis of this pilot study birch saplings are expected to respond similarly to natural increases in air temperature.These results should be interpreted and applied to the older trees with caution while considering that stress incurred at a young may be reflected in the future tree growth.
AcknowledgementsWe acknowledge the valuable assistance of the colleagues from the Institute of Forestry and Vytautas Magnus University during the experiments.
 Journal of Forestry Research2018年6期
Journal of Forestry Research2018年6期
- Journal of Forestry Research的其它文章
- Temperature characteristics of wood during microwave treatments
- Tin-based metal bath heat treatment:an efficient and recyclable green approach for Pinus massoniana wood modification
- Decay resistance and dimensional stability improvement of wood by low melting point alloy heat treatment
- A novel image segmentation approach for wood plate surface defect classification through convex optimization
- Ectomycorrhizal fungus enhances drought tolerance of Pinus sylvestris var.mongolica seedlings and improves soil condition
- First report of leaf-spot disease caused by Sphaeropsis visci on Asian mistletoe[Viscum coloratum(Kom.)Nakai]in China