
牛分枝杆菌mpb51-mpb70融合基因的原核表达与抗原活性初步鉴定
2018-11-02 10:23姜秀云周步丹包艳红陈敬蕊王春芳徐博文马红霞
中国人兽共患病学报 2018年9期
姜秀云,周步丹,董 阳,包艳红,陈敬蕊,刘 磊,王春芳,徐博文,马红霞
牛结核病(Bovine tuberculosis)主要是由牛分枝杆菌(Mycobacteriumbovis)引起的以牛、鹿等动物为主的动物和人慢性细菌性疫病,世界动物卫生组织(Office International Des Epizooties,OIE)将其列为B类传染病。目前,牛结核菌素(Purified Protein Derivative,PPD)变态反应是OIE指定的唯一检测牛结核病的方法[1],但是该方法特异性差,并且对阳性牛行隔离或捕杀所带来经济损失在许多国家无力承担。另因卡介苗(Bacillus of Calmette and Guerin,BCG)无法用于牛结核病的预防。因此,研发牛结核病的新型诊断试剂和预防疫苗。
MPB51和MPB70皆是结核分枝杆菌的主要分泌蛋白,且只存在于结核分枝杆菌复合群中,两者在临床检测中具有良好的敏感性和特异性[2-3]。MPB51中存在CD4+T和CD8+T 细胞抗原表位,可诱导免疫小鼠脾细胞分泌高水平的IFN-γ[4]。MPB70中存在多个产生IFN-γ 的Th1型细胞抗原表位,mpb70重组质粒DNA能激发免疫动物产生Th1型应答反应,但MPB70在不同BCG菌株中表达水平较低或缺失[5],影响了BCG的临床效果。可见,MPB51和MPB70均是结核分枝杆菌的保护性抗原,可作为新型诊断抗原和疫苗的候选蛋白。据报道,抗体检测能提高牛结核病诊断的敏感性[6-8],融合蛋白或多重抗原可以提高牛结核病检测的敏感性和特异性[9-11],并具有良好的免疫原性[12-14]。为开发牛结核病敏感、特异的诊断抗原和高效重组疫苗,利用重叠拼接 (Splicing by Overlapping Extension,SOE) PCR技术实现牛分枝杆菌mpb51和mpb70基因相融合,在大肠杆菌中进行表达,并明确其抗原性。为进一步研究MPB51-MPB70作为新型检测抗原及疫苗奠定基础。
1 材料与方法
1.1菌株、质粒与载体EscherichiacoliJM109、E.coliDH5ɑ、E.coliBL21(DE3)、pET28a(+)由笔者所在实验室保存。pGEM-T-51[15]、pGEM-T-70[16]由笔者构建与保存。T-Vector pMD19购于TaKaRa公司。
1.2主要试剂 Pyrobest DNA Polymerase、LATaqDNA聚合酶、dNTPs、BamHⅠ、EcoRⅠ、T4 DNA连接酶及DNA Markers、DNA回收纯化试剂盒、质粒小量提取试剂盒为TaKaRa公司产品;琼脂糖为Spanish公司产品;溶菌酶、牛血清白蛋白(BSA)为Bebco公司产品;氨苄青霉素(Amp)、卡那霉素(Kan)、异丙基硫代半乳糖苷(IPTG)、RNase A为Sigma公司产品;酵母浸出粉、胰蛋白胨为Oxoid公司产品;牛结核血清自剖检结核病变牛血液分离;HRP标记兔抗牛IgG二抗为北京鼎国生物工程有限公司产品。Ni-NTA Agarose为克劳宁(北京)生物科技有限公司产品。
1.3引物的设计与合成 根据mpb51、mpb70的基因序列设计PCR引物,序列如下:P151: 5′ TAGGATCCATGGCCCCATACGAGAA 3′,P251: 5′CGATCCGCCACCGCCAGAGCCTCCACCTCCTGAACCGCCTCCACCGCGGATCGCACC 3′,P170: 5′GGTGGAGGCGGTTCAGGAGGTGGAGGCTCTGGCGGTGGCGGATCGGGCGATCTGGTGGGC3′,P270: 5′GCGGAATTCTTACGCCGGAGGCATTAG 3′, P151、P270的斜体部分分别是BamHⅠ和EcoRⅠ内切酶酶切位点。上述引物由北京三博远志生物技术有限公司合成。
1.4目的基因的扩增及产物的纯化 以pGEM-T-51、pGEM-T-70为模板,以P151和P251、P170和P270为引物,利用Pyrobest DNA Polymerase分别对mpb51与mpb70基因进行扩增。以mpb51和mpb70基因的PCR纯化产物为模板,以P151和P270为引物,以LATaqDNA 聚合酶进行扩增,反应条件为:98 ℃ 5 min;98 ℃ 30 s、60 ℃ 1 min、72 ℃ 90 s,35个循环。产物经1%琼脂糖凝胶电泳后切胶,并利用DNA凝胶回收纯化试剂盒纯化。
1.5mpb51-mpb70融合基因的克隆和序列测定 用 Solution I 对纯化的mpb51-mpb70 PCR产物和T-Vector pMD19进行连接,构建重组质粒pMD-51-70,转化至E.coliJM109感受态细胞中,37 ℃ 180 r/min培养1.5 h,涂布于含Amp(100 mg/L)的LB固体培养基中,37 ℃过夜培养。挑取转化菌接种于5 mL含Amp 的LB液体培养基中,37 ℃ 180 r/min过夜培养。利用小量提取试剂盒提取质粒,通过BamHⅠ、EcoRⅠ进行双酶切鉴定。选取阳性质粒送至南京金斯瑞生物科技有限公司测序。
1.6原核表达质粒的构建与鉴定 对重组质粒pMD-51-70和pET28a(+)均以BamHⅠ、EcoRⅠ进行双酶切,纯化的产物利用T4 DNA Ligase连接,构建重组表达质粒pET-51-70,转化至E.coliBL21(DE3)感受态中,涂布于含Kan(50mg/L)的LB固体培养基中,37 ℃培养过夜。挑取转化菌,提取质粒,进行双酶切鉴定。
1.7融合基因的表达与分析 将重组表达菌株接种于5 mL含Kan的LB液体培养基中,37 ℃过夜培养。取2 mL菌液接种至50 mL含Kan 的LB液体培养基中,37 ℃ 200 r/min培养至OD600nm≈0.6-0.7,用终浓度为1 mmol/L的IPTG进行诱导7 h,期间每隔1 h留取少许菌液。将留取的菌液OD600nm调至0.6,取菌液1.2 mL,离心,于沉淀中加蒸馏水100 μL。样品处理后,进行SDS-PAGE分析。
1.8重组蛋白的纯化 表达MPB51[15]、MPB70[16]及MPB51-MPB70的重组菌株经IPTG 于25 ℃下诱导,收集菌体,低温超声破碎后,按产品说明书进行Ni-NTA纯化。纯化的蛋白于-20 ℃保存。
1.9融合蛋白的鉴定 纯化的融合蛋白经SDS-PAGE后,通过半干式碳板转印槽转移至PVDF膜上,1%BSA封闭后,以牛结核血清为一抗,HRP标记兔抗牛IgG抗体为二抗,联苯胺为底物,进行Western blotting鉴定。
1.10融合蛋白抗原性鉴定与比较 以不同浓度的MPB51-MPB70为抗原,不同稀释倍数的牛结核阴、阳性血清为一抗,1∶2 000 HRP标记兔抗牛IgG为二抗,邻甲苯胺为底物,通过棋盘法确定间接ELISA最适抗原包被浓度和血清稀释倍数。以阳性血清OD492nm>1,P/N比值最大者的抗原浓度和血清稀释倍数作为MPB51-MPB70的最适包被浓度和最适血清稀释倍数。以MPB51-MPB70最适包被浓度和血清最适稀释倍数对10头份牛结核阴、阳性血清进行间接ELISA检测,并与MPB51(包被浓度为3/5的MPB51-MPB70)、MPB70(包被浓度为2/5的MPB51-MPB70)为抗原的间接ELISA比较抗原性。
2 结 果
2.1目的基因的PCR扩增产物mpb51、mpb70及mpb51-mpb70扩增产物的电泳结果如图1所示。从图中可见,扩增的mpb51、mpb70和mpb51-mpb70 DNA片段分别是849 bp、555 bp、1359 bp,均和预期DNA片段大小相一致。
2.2mpb51-mpb70 融合基因的克隆与鉴定 mpb51-mpb70 融合基因PCR产物与pMD19 连接,获得了重组质粒pMD-51-70,经BamHⅠ、EcoRⅠ酶切,电泳可见2 692 bp的pMD19线性片段和1 359 bp的插入片段(图2)。序列测定表明,mpb51-mpb70 融合基因的大小为1359 bp,与目的片段大小一致,且没有序列插入、缺失及改变的现象,成功地获得了牛分枝杆菌mpb51-mpb70融合基因。
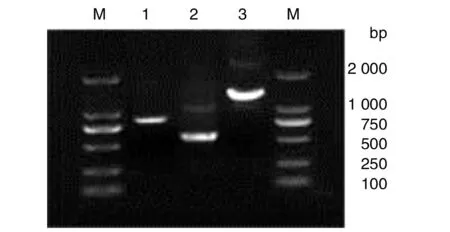
1-3: 分别为mpb51、mpb70和mpb51-mpb70基因的 PCR产物;M: DNA markerLane M, DNA marker /DL2000; Lanes 1-3, PCR products of mpb51, mpb70 and mpb51-mpb70 genes, respectively图1 mpb51、mpb70和mpb51-mpb70的PCR 扩增Fig.1 PCR amplification of mpb51, mpb70 and mpb51-mpb70 genes
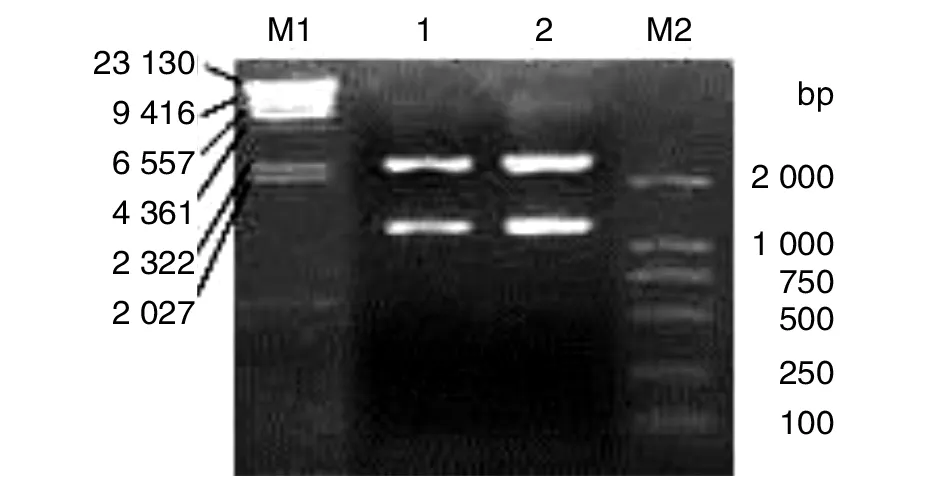
1-2: pMD-51-70 质粒的酶切产物; M1: DNA marker λDNA/Hind III; M2: DNA marker/DL2000Lanes 1-2, products of pMD-51-70 digested by enzyme; Lane M1, DNA marker λDNA/Hind III; Lane M2,DNA marker/DL2000图2 重组质粒的酶切鉴定Fig.2 Restriction analysis map of pMD-51-70
2.3原核表达重组质粒的构建与鉴定 在T4 DNA Ligase 的作用下,mpb51-mpb70和pET28a(+)连接,经BamHⅠ、EcoRⅠ酶切,电泳后见到5367 bp pET28a(+)载体片段和1 359 bp 插入片段(图3)。成功地构建了mpb1-mpb70重组原核表达质粒pET-51-70。
2.4融合蛋白的表达分析 经IPTG诱导,E.coliBL21(DE3)中pET-51-70在各时间表达蛋白的SDS-PAGE结果如图4所示。从图可见,融合基因mpb51-mpb70 获得了表达,其表达量随诱导时间的延长而增加,诱导3 h达到高峰,MPB51-MPB70融合蛋白的相对分子量约为45 ku。而pET28a(+)E.coliBL21(DE3)中则未见该融合蛋白表达。
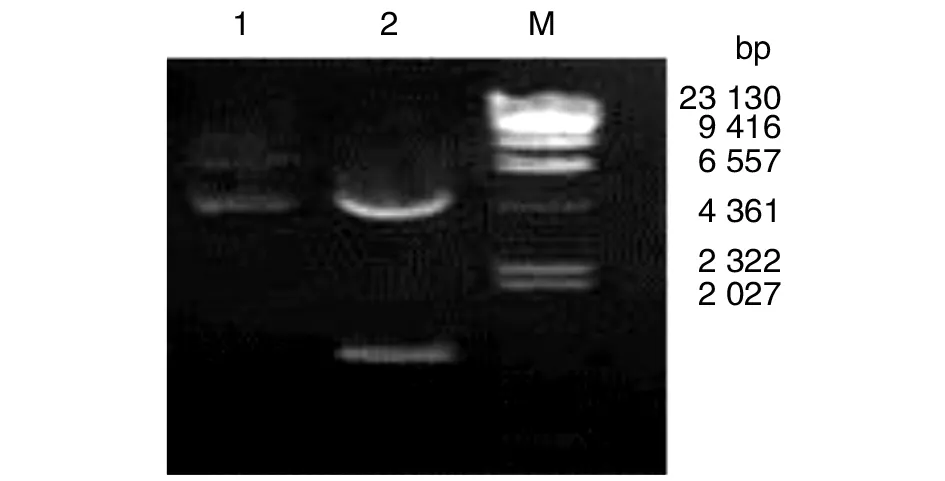
1: pET-51-70; 2: pET-51-70 的酶切产物; M1:DNA 分子量标准; M: DNA MarkerLane 1, pET-51-70; Lane 2, Products of pET-51-70 digested by enzyme; Lane M, λ DNA digested with Hind Ⅲ图3 原核表达质粒的酶切鉴定Fig.3 Restriction analysis map of pET-51-70
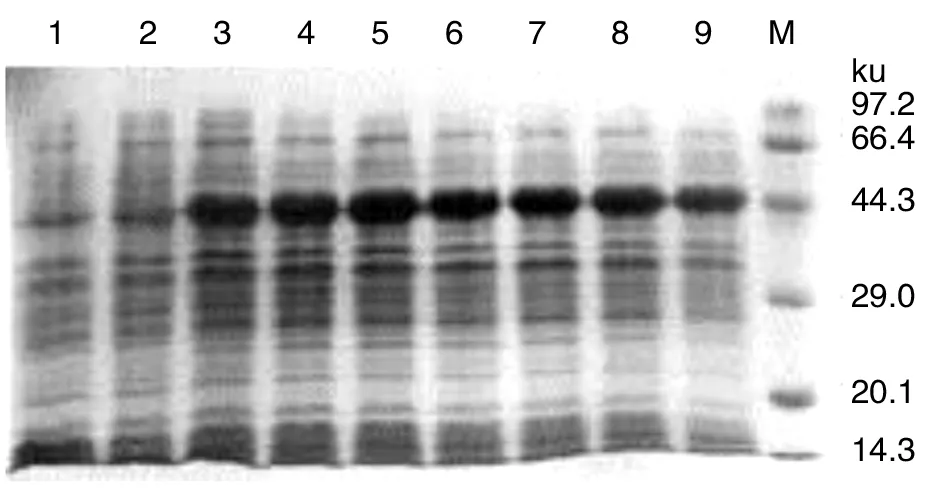
1:pET-28a(+) 在E.coli BL21诱导7h; 2-9:分别是pET-51-70在E.coli BL21诱导0-7 h; M:蛋白质MarkerLane1, pET-28a(+) expression result in E.coli BL21 with IPTG induced 7h, as control; Lanes 2-9, expression results in E.coli BL21 with IPTG induced 0-7h, respectively; Lane M, low molecular weight protein marker图4 mpb51-mpb70在E.coli BL21中的表达Fig.4 The mpb51-mpb70 recombinant expression plasmid expressed by prokaryotic expression vector in E.coli BL21
2.5目的蛋白的纯化 MPB51-MPB70、MPB70、MPB51经Ni-NTA纯化后的SDS-PAGE结果如图5所示。图中可见纯度较高的目的蛋白带。
2.6融合蛋白的鉴定 纯化的MPB51-MPB70融合蛋白Western blotting 结果如图6所示。从图中可见45 ku处存在一条蛋白印迹带,证明该融合蛋白MPB51-MPB70能够与牛结核血清反应,且具有较好的反应性。
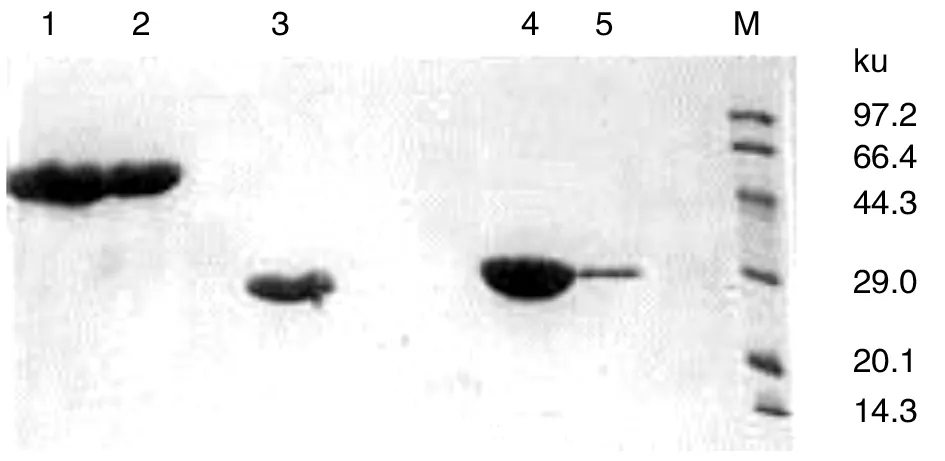
1-2: MPB51-MPB70;3:MPB70;4-5:MPB51;M:蛋白质markerLanes1-2, MPB51-MPB70; Lane 3, MPB70; Lanes 4-5, MPB51; Lane M, low molecular weight protein marker图5 原核表达蛋白的纯化Fig.5 Purification of prokaryotic recombinant expression proteins
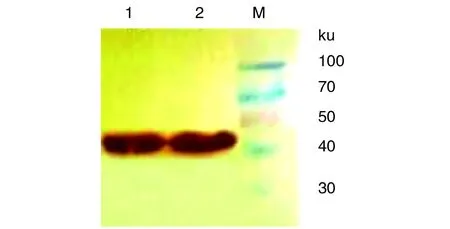
1-2: MPB51-MPB70; M: 蛋白质MarkerLanes 1-2, MPB51-MPB70 fusion protein; Lane M, low molecular weight protein marker图6 MPB51-MPB70的Western blotting鉴定Fig.6 Western blotting of the MPB51-MPB70 fusion protein
2.7MPB51-MPB70 的抗原性 通过棋盘法确定MPB51-MPB70的最适包被浓度为50 ng/mL,血清最适稀释倍数为400倍。以MPB51-MPB70(50 ng/mL)、MPB51(30 ng/mL)、MPB70(20 ng/mL)对牛结核阴、阳性血清的间接ELISA检测结果如图7所示。从图中可见,同一阳性血清的反应性(OD492nm)MPB51-MPB70比MPB51、MPB70均高,而3种重组蛋白对阴性血清检测结果均较低(OD492nm<0.12),表明MPB51-MPB70具有良好的抗原性。
3 讨 论
长期以来,在动物结核病的防制方面,尽管采取检疫、隔离与淘汰等一系列措施,也未能从根本上控制该病。其原因在于没有切实可行的疫苗,因此,开发新型、高效、安全的疫苗一直是动物结核病研究的重点所在。另外,结核病的诊断抗原敏感性或特异性尚待提高,因此,研发敏感、特异的诊断抗原也一直倍受注目。
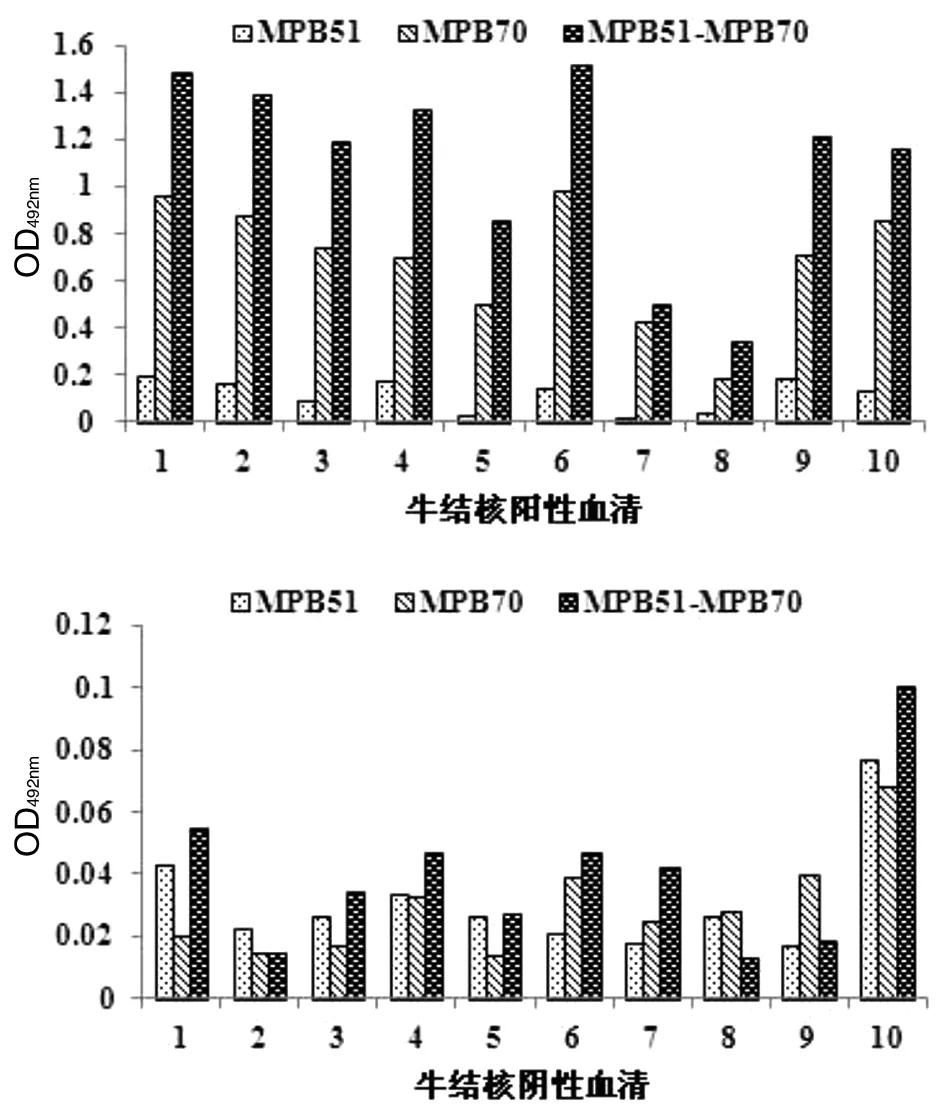
图7 重组蛋白的间接ELISA检测Fig.7 Indirect ELISA detection of bovine serum by recombinant protein
在结核病的血清学诊断中,Bethunaickan等[17]用MPT51检测了痰液涂片和培养阳性的结核病患者血清中特异性IgG和IgA,结果显示IgG的敏感性和特异性分别为57%、98%,IgG+IgA的敏感性和特异性分别为71%、95%。Cho等[18]通过蛋白质组学分析发现,牛分枝杆菌MPB70、MPB83、MtbE等20 kD高抗原性蛋白是适于结核病牛进行ELISA检测的抗原。Cho等[3]报道,与PPD比较,牛分枝杆菌MPB70对感染结核的牛血清有着更高的敏感性和特异性,分别是84%和100%。Waters等[6]采用IDEXXM.bovisELISA(MPB83/MPB70融合抗原)检测了自然感染结核的牛血清,其敏感性和特异性分别是63%和98%,并可作为感染牛皮试阴性的补充试验。Souza等[9]比较了CFP-10、ESAT-6、MPB83、Mb0143、PE5、PE13、TB10.4、TB15.3及ESAT-6/MPB70/MPB83融合蛋白检测皮试阳性牛的血清,结果显示ESAT-6/MPB70/MPB83融合蛋白的敏感性是83.2%,高于每1种单一蛋白。
Wang 等[19]研究发现,MPT51能诱导免疫小鼠产生高水平IFN-γ,可作为结核病疫苗和感染分析的目标蛋白。Hashimoto等[20]构建了MPT51重组慢病毒,诱导小鼠肺部产生特异性CD8+T细胞免疫应答。Uchijima等[21]报道了编码MPT51和巨噬细胞炎性蛋白-1α(MIP-1α)融合蛋白的DNA疫苗能够诱导小鼠产生高水平的特异性CD8+T细胞应答反应。de Sousa等[12]评估了结核杆菌重组ag85c-mpt51-hspx(CMX)融合蛋白的抗原性,结果显示,免疫小鼠体内产生了高水平的抗CMX特异性IgG1和IgG2a,脾细胞中特异性TCD4 IFN-γ+和TNF-α+的数量增加,活动性肺结核患者血清中抗CMX IgG和IgM明显高于健康的人 (P<0.0001)。Junqueira-Kipnis等[22]构建了结核杆菌Ag85c、MPT51、HspX (CMX)重组耻垢分枝杆菌,小鼠免疫试验显示,CMX重组疫苗可诱导特异性IgG1或IgG2a反应,肺和脾中CMX特异性CD4+T和CD8+T细胞显著增加(P<0.05),并产生IFN-γ、IL-17、TNF-α和IL-2。
综上研究结果表明,MPB51和MPB70在结核病诊断和疫苗研究中具有较好的应用前景。因此,为提高单一蛋白的抗原性,采用 SOE PCR 将牛分枝杆菌保护性抗原基因mpb51和mpb70以45个核苷酸Link连接,使两个蛋白之间存在15个氨基酸(Gly4Ser)3的柔性肽段,以保证各自蛋白的空间结构及免疫活性。本研究成功地获得了mpb51-mpb70 融合基因,序列分析证实,碱基序列一致,编码区域正确。在此基础上,构建了mpb51-mpb70 融合基因的原核表达重组质粒,并在大肠杆菌中表达了融合蛋白。纯化的MPB51-MPB70经间接ELISA检测显示出比单一蛋白更高的抗原反应性。为进一步研究牛结核病的诊断试剂、新型疫苗奠定了基础。
猜你喜欢
——一道江苏高考题的奥秘解读和拓展
中学生物学(2022年7期)2022-09-07
成都医学院学报(2022年4期)2022-08-19
保健医苑(2022年5期)2022-06-10
医学概论(2022年4期)2022-04-24
大众科学(2022年3期)2022-04-09
江西农业学报(2021年4期)2021-04-20
昆明医科大学学报(2021年1期)2021-02-07
三农资讯半月报(2020年11期)2020-06-21
癌变·畸变·突变(2015年3期)2015-02-27
上海预防医学(2009年12期)2009-01-27
