
许氏平鲉微卫星多重PCR体系构建及亲本对子代贡献率
2018-09-11 09:18:02韩慧宗刘阳王腾腾张明亮王斐孙娜姜海滨
中国渔业质量与标准 2018年4期
韩慧宗,刘阳,2,王腾腾,张明亮,王斐,孙娜,2,姜海滨*
(1. 山东省海洋资源与环境研究院, 山东省海洋生态修复重点实验室, 山东 烟台 264006;2. 上海海洋大学水产与生命学院, 上海 201306)
许氏平鲉(Sebastesschlegelii),隶属于鲉形目(Scorpaeniformes)、鮶科(Scorpaenidae)、平鲉属(Sebastes),俗称黑鮶,是中国黄、渤海区常见的近海冷温性底层岩礁经济鱼类。由于其肉质鲜美细腻,营养丰富,赢得了北方地区“黑石斑”的美誉,深受国内外市场欢迎。同时具有洄游范围小、抗病力强、生长速度快、可自然越冬等特点,已成为中国北方沿海网箱养殖、增殖放流、休闲垂钓和海洋牧场的重要鱼种。许氏平鲉属于卵胎生鱼类,性成熟的雌雄亲鱼常在每年11月左右交尾,交尾后精子以“精囊”的形式储存在雌鱼卵巢内,分布于输卵管、卵巢膜以下等位置[1],至翌年的4月中旬开始受精,胚胎在雌鱼体内发育,出膜后继续发育,至5月左右产出自由活动的仔鱼[2]。近二十年来,随着自然采捕捞量的增加,许氏平鲉野生资源显著减少,北方沿海开展了人工增殖放流工作[3-4]。由于市场消费和优良苗种需求量巨大,本课题组从2006年开始陆续开展了许氏平鲉良种选育工作。
许氏平鲉交配行为一般发生在晚上,不易观察,初冠囡等[5]通过对雌亲家系后代研究发现许氏平鲉存在“一雌多雄”的受精方式,而 “一雄多雌” 的受精方式目前仍没有定论。同时,单条许氏平鲉成熟雌鱼产仔量在10万尾以上,有限数量亲本就可以繁殖大量的后代群体,而参与交配的繁殖亲本对后代贡献的大小直接影响人工选育和放流群体遗传多样性[6]。Sugama等[7]对真鲷(Pagrusmajor)亲本对子代贡献率分析中也发现,只有不到30%的亲本对后代基因库有贡献。陈睿毅等[8]用8雄5雌牙鲆亲鱼配组,结果亲本贡献率最高达到47.34%,最低仅为0.53%。亲本对子代贡献率不均衡通过累代繁殖,可能造成群体的遗传多样性降低。
在水产动物研究领域,微卫星多重PCR技术因具有省时高效、低成本、高通量等优点,能够降低分型过程重复带来的人工误差[9],已广泛应用于水产动物亲子鉴定中,如大菱鲆(Scophthalmusmaximus)[10]、凡纳滨对虾(Litopenaeusvannamei)[11]、长牡蛎(Crassostreagigas)[12]、紫菜(Porphyrahaitanensis)[13]等。基于单个微卫星位点对许氏平鲉群体遗传分析、遗传图谱构建等研究已得到应用[14-15],但目前尚未有关于许氏平鲉多重PCR体系的构建及应用的报道。本研究利用本课题组开发的13对高多态性微卫星标记进行优化组合,建立了许氏平鲉3个三重PCR和2个两重PCR反应体系,对雌雄交尾成功的4个母本和所产家系及10个候选父本进行亲权鉴定,在检测许氏平鲉亲子鉴定准确率的同时,分析其交配模式,计算不同父本对子代的贡献率,旨在为许氏平鲉良种育种和苗种繁育提供理论参考。
1 材料与方法
1.1 实验材料
于2016年10月,在烟台泰华海洋科技有限公司选取健康并达到性成熟且性腺发育良好的雌、雄亲鱼各10尾,放养于工厂化室内养殖池内,通过控温控光技术,让其自由交尾。待到2017年4月底,雌、雄亲鱼未出现死亡现象,有4尾雌性亲鱼腹部隆起明显,分别放于4个养殖池内待产,于5月1日至5月3日三天内分别产出自由游动的仔鱼。样品采集10个雄性亲鱼背鳍,编号为Ma1、Ma2、Ma3……Ma10,于7月份样品采集4尾雌性亲鱼背鳍,编号为Fe1、Fe2、Fe3和Fe4,每尾雌鱼所产后代为1个雌亲家系(即半同胞家系),每个雌亲家系取子代94尾,均放置于75%无水乙醇中,-20 ℃保存备用。亲鱼培育水温为自然海水水温,仔鱼培育水温为14~20 ℃。
1.2 实验方法
1.2.1 基因组DNA的提取
采用传统的饱和酚-氯仿-异戊醇法[14]进行实验样品的基因组DNA提取。提取完成后用0.8%的琼脂糖凝胶电泳检测其完整性,通过分光光度计测定DNA浓度和纯度,将DNA质量浓度稀释至50 ng/μL,-20 ℃保存备用。
1.2.2 微卫星候选标记的筛选
从本课题组已开发的多态性较高且扩增效果较好的微卫星引物中挑选25对[4,14]。微卫星引物由上海生工生物工程有限公司合成。以扩增效率高、多态性高、杂带较少、目的条带清晰且不相重合为基本原则,挑选最优的13对微卫星引物用于多重PCR体系的构建,PCR体系引物的位点组合、重复单元、碱基序列、产物大小、退火温度如表1所示。
1.2.3 PCR反应及产物检测
PCR反应体系为25 μL,包含17.8 μL ddH2O,2.5 μL 10×PCR Buffer (Mg2+),0.5 μL 10 mmol/L dNTPs,正反引物各1 μL,1 U Taq酶,2 μL模板DNA。反应程序为:94 ℃预变性5 min;94 ℃变性40 s、退火40 s、72 ℃延伸1 min,35个循环;72 ℃延伸10 min,4 ℃下保存。PCR产物用8%非变形聚丙烯酰胺凝胶电泳检测,用10 bp DNA ladder标记等位基因位置,使用银染染色法染色,用Bio-5000 Pus扫描仪扫描电泳图谱,扩增产物目的片段大小由Gel-Pro analyzer 32凝胶分析软件定量。
1.2.4 微卫星多重PCR优化组合
根据各个微卫星位点的退火温度和PCR扩增片段大小差异性,优先利用两个微卫星位点的两两组合,同时选择扩增片段不重叠、退火温度相近(温差不超过5 ℃)、引物序列不出现错配和引物二聚体的微卫星位点进行两重PCR构建。在此基础上,加入第3个引物,筛选构建三重PCR体系。通过对引物退火温度、引物浓度比例、反应体系等条件去优化,确定最佳的反应条件。
1.3 数据统计分析
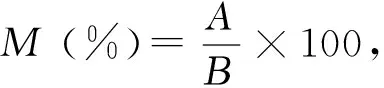
2 实验结果
2.1 许氏平鲉微卫星多重PCR体系的建立
本研究利用13对微卫星构建了3个三重PCR和2个二重PCR体系。图1示体系Multiplex Set4和Multiplex Set2在部分子代中的聚丙烯酰胺凝胶电泳图谱,目的条带清晰且不重合、染色均匀、准确性较高。结果表明,实验构建的微卫星5组多重PCR可用于交配后代的亲权鉴定分析和亲本贡献率研究。
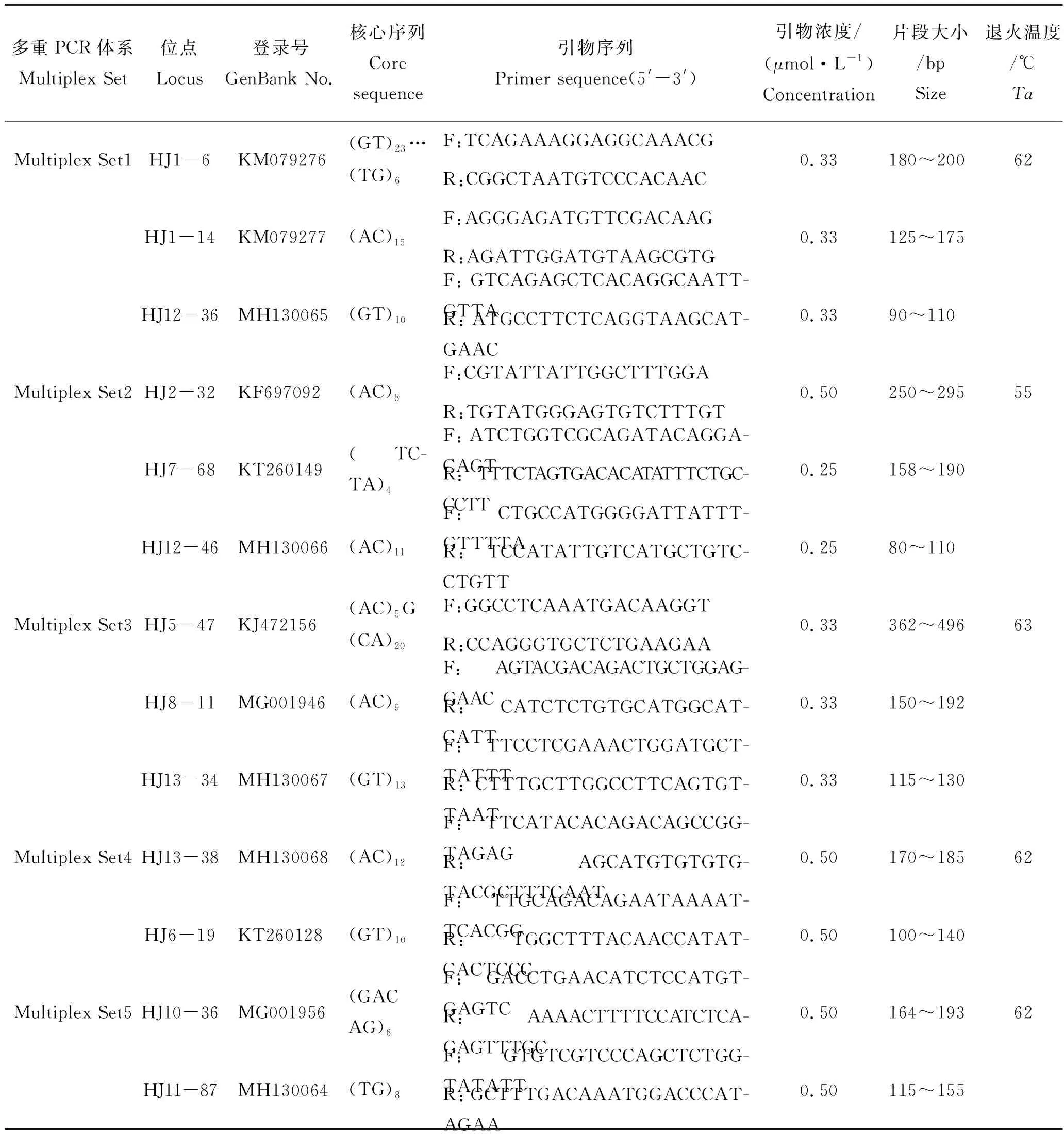
表1 许氏平鲉5组微卫星多重PCR特征Tab.1 The characteristics of 5 microsatellite multiplex PCRs in Sebastes schlegelii
2.2 许氏平鲉微卫星位点的多态性分析
表2展示了5组多重PCR中13对微卫星标记在4个家系中的遗传参数、排除率和累积排除率。等位基因数(Na)为6~13,观测杂合度(Ho)为0.452~1.000,期望杂合度(He)为0.445~0.810;多态信息含量(PIC)为0.404~0.777,参照Botstein等[20]制定的标准显示3个位点为中度多态性;其余均为高度多态性,无效等位基因频率F(Null)为-0.300~0.014。在4个家系子代中平均等位基因数均为9,平均观测杂合度依次分别为0.966、0.993、0.955和0.963,平均期望杂合度依次分别为0.733、0.751、0.698和0.678,平均多态信息含量依次分别为0.684、0.706、0.650和0.620。4个家系子代之间遗传差异性不大,均表现出较高的遗传多样性。

图1 Multiplex Set4 (a)和Multiplex Set2 (b)在部分子代中的电泳图谱Fig.1 The electrophorograms of Set4 and Set2 multiplex PCR systems in some offspring
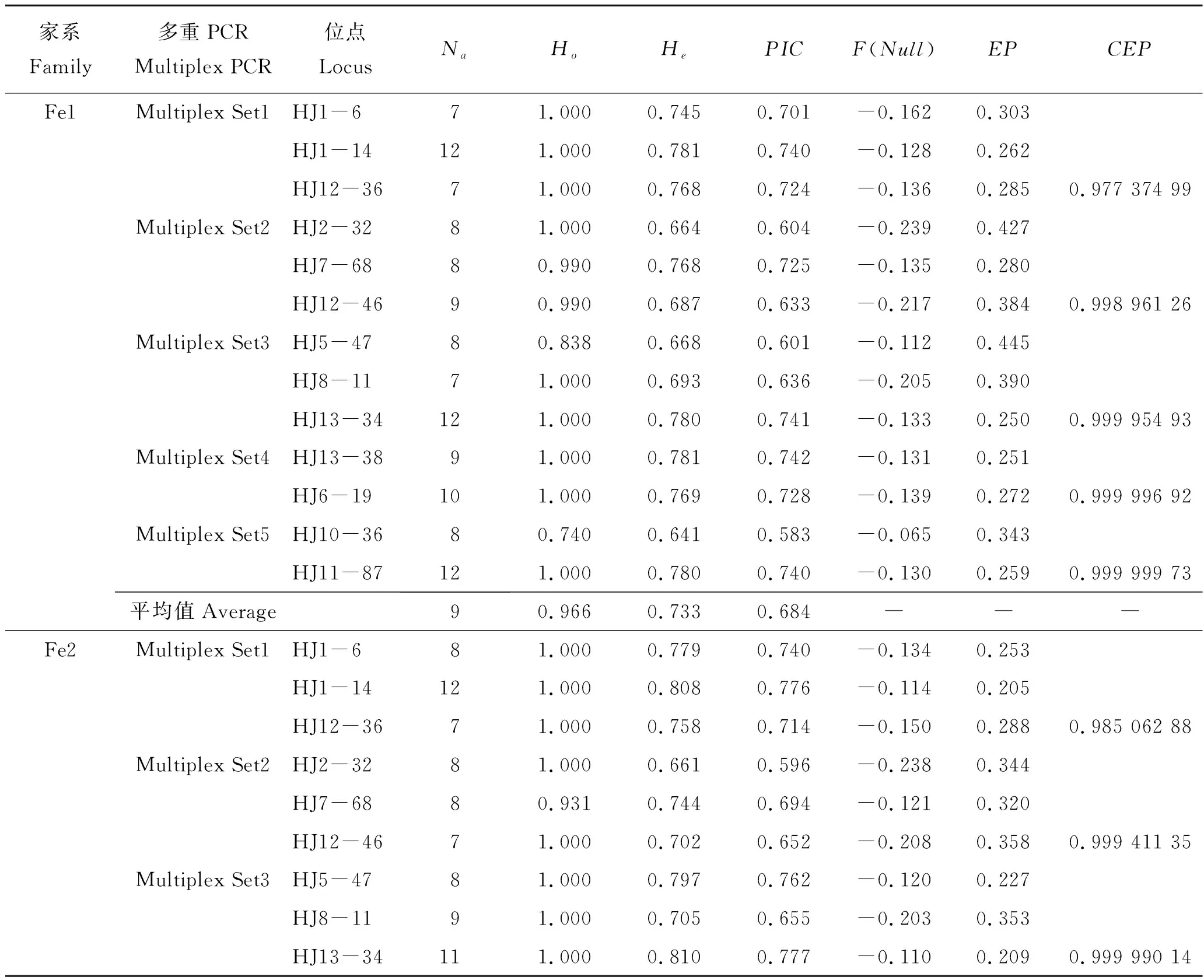
表2 5组多重PCR在4个家系中的遗传参数、排除率和累计排除率Tab.2 Genetic parameter, exclusion probability and cumulative exclusion probability of 5 multiple PCR systems in Sebastes schlegelii
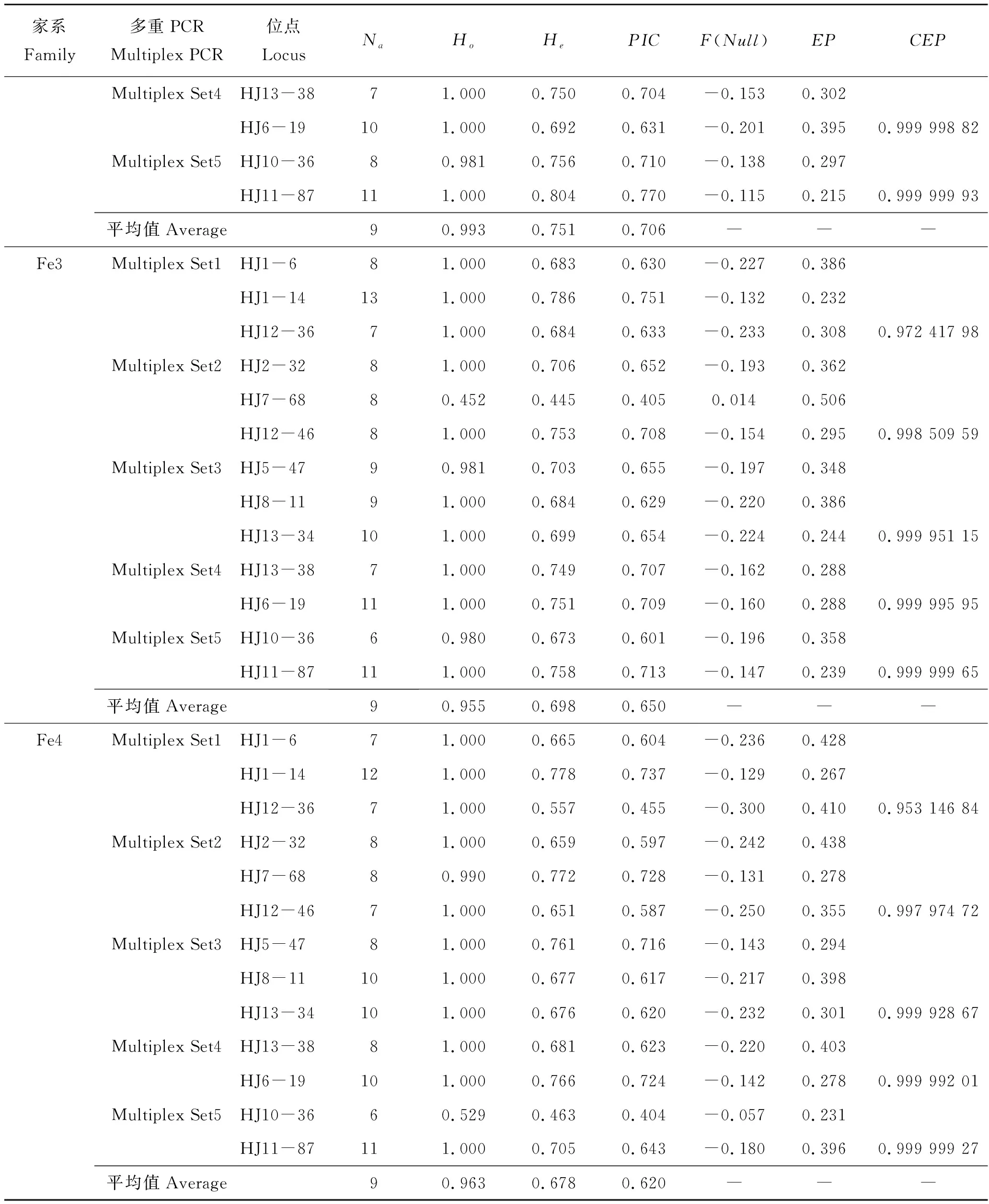
续表2,Tab.2 Continued
2.3 亲子鉴定的排除率和鉴定准确率分析
由表2和图2可以看出,鉴定4个家系时,双亲基因型已知的非亲排除率(EP)分别介于0.250~0.445、0.205~0.395、0.232~0.506、0.267~0.438。当使用3个三重PCR时累积排除率(CEP)依次分别为0.999 954 93、0.999 990 14、0.999 951 15和0.999 928 67,CEP>0.999 9,不能准确找到父本的子代数依次分别为3、2、4和3个,亲子鉴定准确率依次分别为96.81%、97.87%、95.74%和96.81%;当使用3个三重PCR和1个两重PCR时CEP依次分别为0.999 996 92、0.999 998 82、0.999 995 95和0.999 992 01,CEP>0.999 99,不能准确找到父本的子代数依次分别为1、1、2和2个,亲子鉴定准确率依次分别为98.94%、98.94%、97.87%和97.87%;当使用3个三重PCR和2个两重PCR时,CEP分别为0.999 999 73、0.999 999 93、0.999 999 65和0.999 999 27,CEP>0.999 999,4个家系均能找到相应父本,亲子鉴定准确率均达到100%。
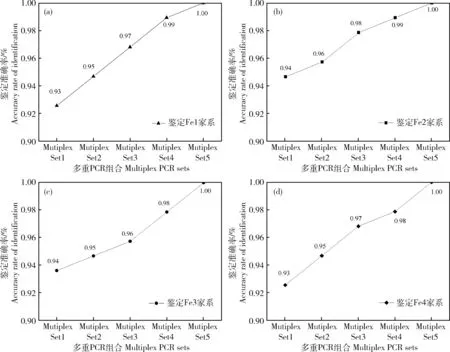
图2 5组多重PCR在4个家系中的亲权鉴定准确率Fig.2 The accuracy rates of parentage assignment for 5 multiplex PCRs in four families
2.4 亲子鉴定交配模式及父本贡献率分析
在鉴定成功率100%的情况下,筛选出4个家系子代最有可能的父本,如表3和表4所示。Fe1家系中,鉴定出父本数为6个,分别为Ma2、Ma3、Ma4、Ma5、Ma7和Ma8;Fe2家系中,鉴定出父本数为5个,分别为Ma2、Ma3、Ma5、Ma7和Ma10;Fe3家系中,鉴定出父本数为4个,分别为Ma2、Ma4、Ma7和Ma10;Fe4家系中,鉴定出父本综合数为3个,分别为Ma2、Ma6和Ma7。鉴定出许氏平鲉交配模式存在“一雌多雄”的现象。同时,Ma2和Ma7均参与到4个母本交配受精中,Ma3、Ma4、Ma5和Ma10均参与到2个母本交配受精中,所以也存在“一雄多雌”的交配模式。
在4个家系中,父本综合贡献率最高的父本分别为Ma7(32.45%)、Ma2(19.15%)和Ma6(16.22%),其贡献率总和达到67.82%,而Ma3、Ma10、Ma8、Ma4和Ma5五个父本贡献率总和为32.18%。其中Ma1和Ma9未发现子代个体,说明未参与交配或交配受精不成功,其Ma1和Ma9综合贡献率为0。
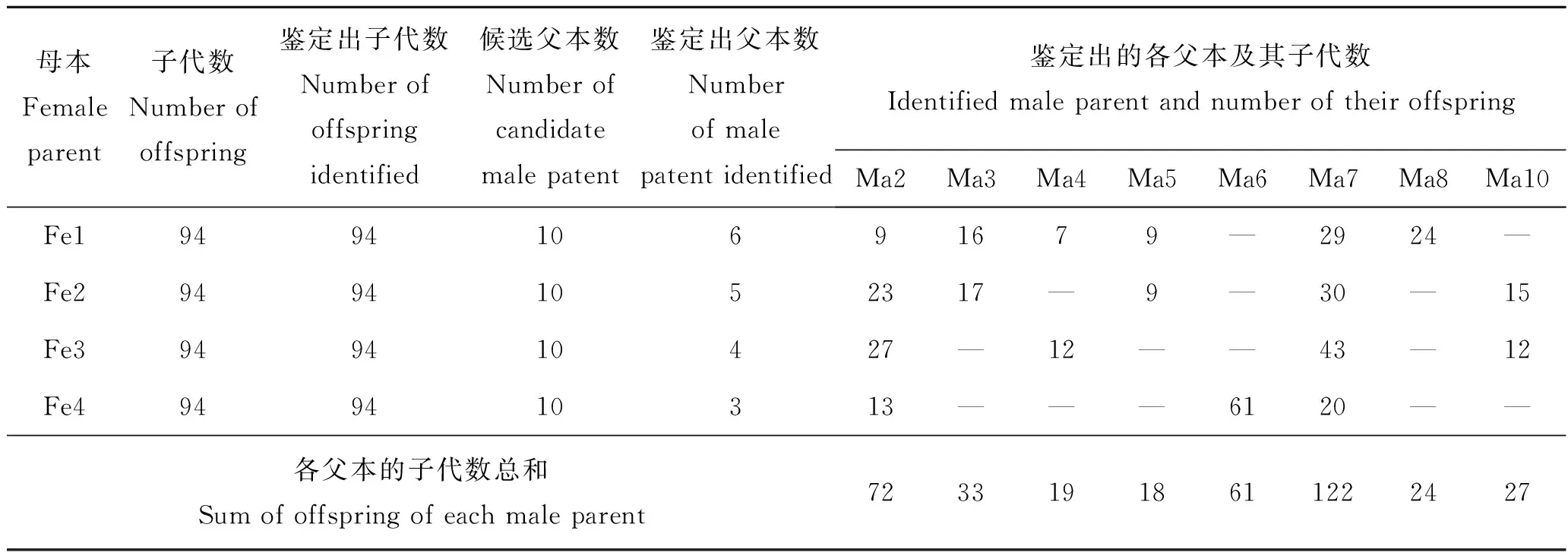
表3 许氏平鲉4个家系的亲子鉴定结果Tab.3 The numbers of parents corresponding to offsprings in the four groups
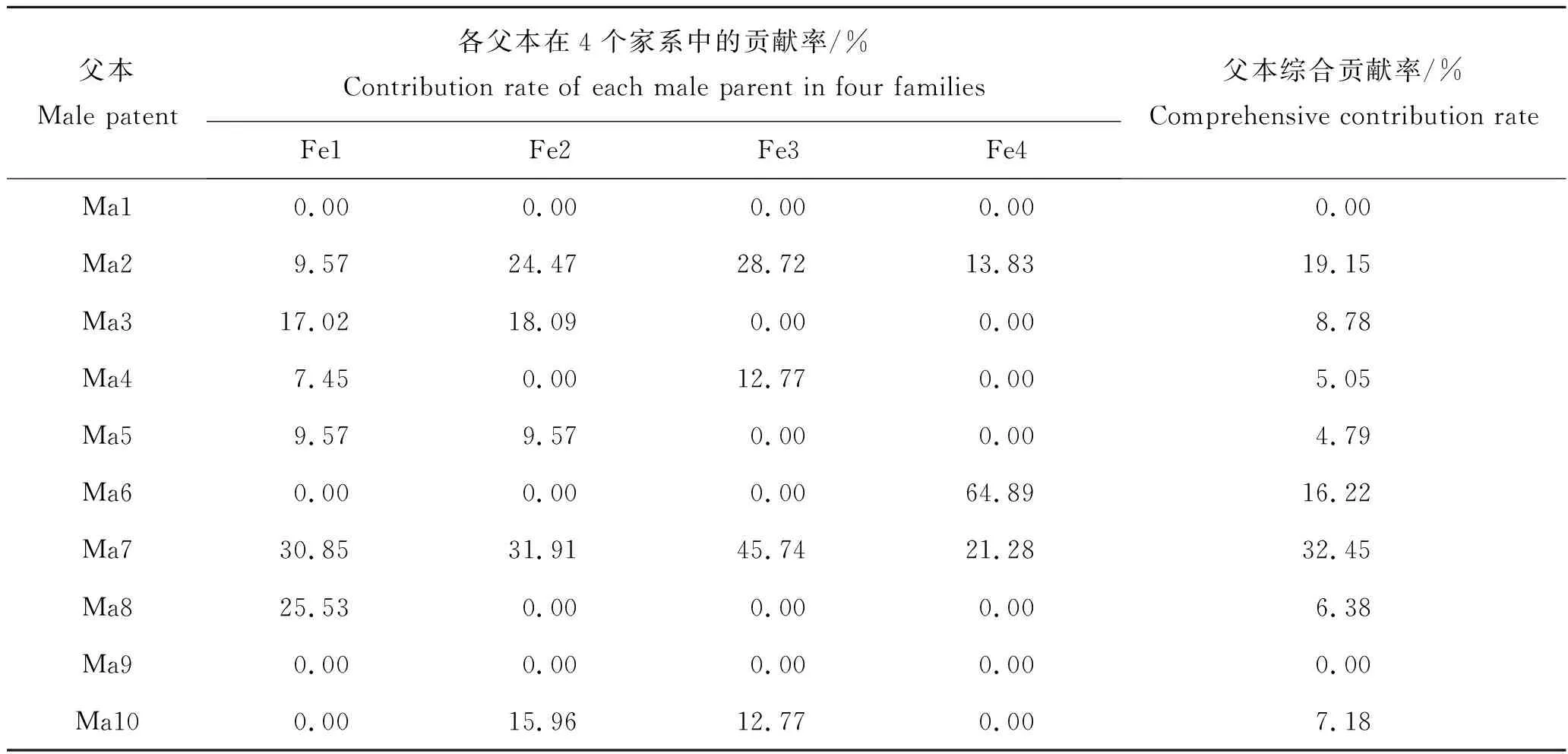
表4 许氏平鲉4个家系对应父本的贡献率Tab.4 The parental contribution rates of offsprings in the four groups
3 讨论
3.1 许氏平鲉微卫星多重PCR体系的构建
理想的多重PCR反应需要不同引物能在同一反应体系中进行特异性扩增,且要求扩增引物的退火温度相近、扩增片段不发生重叠、非特异性扩增条带不存在或较少,与单个位点PCR相比,多重PCR技术难度较大,需要对目的条带和反应条件进行反复实验、反复调整,最终得到最优的反应条件。但构建成功以后,能节省实验材料,提高效率,简化操作步骤[17]。本研究在构建许氏平鲉微卫星PCR体系时发现,引物组合、引物浓度比例和退火温度是影响多重PCR构建成功的主要因素之一,这与李东宇等[11]和张毅等[18]的研究结果相一致。首先,不同引物间组合选择在PCR扩增时要避免形成二聚体或发卡结构的形成;其次,退火温度选择尽量相近或相差不超过5 ℃[19],本研究的每重PCR均采用相同的退火温度,在此温度下引物均可有效扩增;再次,引物浓度比例直接影响多重PCR的扩增效率,过大会导致杂带或二聚体的产生,过小会影响PCR产物的形成[20]。基于以上方面,本研究在建立二重PCR基础上,再引入另外1个位点,经反复实验,直至得到最优多重PCR组合,构建了3个三重PCR体系。本研究构建的3个三重PCR体系和2个二重PCR体系,条带清晰,无重合条带,适用于许氏平鲉的亲子鉴定。
3.2 许氏平鲉家系遗传多样性和亲权分析

亲权鉴定准确率与高多态性微卫星标记数量、待测亲本与子代数量有关,通过使用高多态性的微卫星标记或增加标记数量可以提高准确性[22],同时微卫星标记准确性还与无效等位基因存在有关[23]。Dakin等[24]指出只要无效等位基因频率小于0.2,对结果产生的影响不会太大,只会稍微降低引物的累积排除率。本研究中所用的微卫星位点的无效等位基因频率均小于0.05。本研究是基于最大似然法的亲本分析软件Cervus,通过累积排除率(CEP)来评价该亲权鉴定体系成立与否,累积排除率越高,亲子鉴定率就会越高。参照人类亲子鉴定标准,CEP达到99.95%即可做亲子鉴定[25];王敏[26]研究表明CEP≥99.73%时,即可认定存在亲子关系。本研究中通过3个三重PCR和1个两重PCR鉴定时CEP均达到0.999 99以上,但不能有效鉴定父本数仍有1、1、2和2个。当增加1个多重PCR时4个家系的累积排除率均达到0.999 999以上,均能找到父本鉴定准确率达到100%。这一结果与陈亮等[27]等利用5组多重PCR技术对长鳍吻鮈(Rhinogobioventralis)亲子鉴定的结果相似,表明构建的许氏平鲉微卫星多重PCR反应体系具有很好的可靠性和适用性。
3.3 许氏平鲉的交配模式及父本贡献率分析
水产动物的交配行为因受自然条件的影响、交配时间不确定性或者交配隐秘不易发现,同时观察到的交配行为并不代表受精成功。随着分子标记技术广泛应用于亲本与子代间的亲缘关系的鉴定,其交配模式研究结果更加准确可靠。本研究利用构建的微卫星多重PCR对交尾受精成功的4组许氏平鲉亲本和子代样品进行亲权鉴定,结果显示,4个家系子代的父本数为6、5、4和3个,证明了许氏平鲉存在“一雌多雄”的交配模式,与Gao等[28]对许氏平鲉野生和养殖群体的多重父权分析结果相一致。此交配模式在卵胎生鱼类,如孔雀鱼(Poeciliareticulata)[29]、食蚊鱼(Gambusiaaffinis)[30]、褐菖鲉(Sebastiscusmarmoratus)[31]中,均存在。本研究还发现,有2个父本均参与到4个雌性交配受精过程中,有4个父本分别参与了2雌性交配受精过程中,所以也存在“一雄多雌”的交配模式。一雌多雄交配模式能有效提高子代遗传多样性和繁殖效率,应对环境异常变化[32]。
本研究表明父本对子代的贡献率差异较大,4个家系子代中Ma2、Ma6和Ma7父本贡献率较高,其总和达67.82%,远超其他父本的贡献率,说明参与交配并成功受精的精子多数来自这3个父本;而Ma1和Ma8贡献率却为0,说明Ma1和Ma8未参与交配或交配后未参与受精。贡献率大小的差异是由交配后精子竞争机制导致,精子竞争是指两个或多个精子为争夺与同一卵的受精权而竞争,是一种交配后的性选择机制。由于许氏平鲉雌雄形成熟不同步(雌性比雄性晚),且交配受精后存在精子储存现象,精子储存使得雌雄交配与精卵受精分离,不仅延长了雌性的繁殖周期,也有利于精子竞争及雌性隐性选择,为交配后性选择提供有利条件[33]。刘文芬等[34]研究短蛸(Amphioctopusfangsiao)交配模式时发现也存在交配后性选择现象。另外,在许氏平鲉遗传选育的过程中,亲本对子代贡献率不均衡也可能会导致一些等位基因的丢失,累代交配繁殖后造成后代遗传多样性降低。在许氏平鲉人工交尾进行繁育和选育时要关注有效的繁殖亲本,还要关注贡献率的差异,及时发现无效或贡献率低下的亲本,及时剔除或补充新个体。
4 结论
本研究利用13对许氏平鲉微卫星标记构建了3个三重和2个两重PCR体系,有效鉴定许氏平鲉4个家系的亲权关系,累积排除率均达到0.999 999以上,鉴定成功率100%,鉴定到父本数为3~6个。研究发现许氏平鲉存在“一雌多雄”和“一雄多雌”的交配模式,父本贡献率差异较大,Ma2、Ma6和Ma7父本贡献率总和达67.82%,Ma1和Ma8贡献率为0。本研究为许氏平鲉良种选育和人工繁育提供科学理论依据,也为卵胎生鱼类交配后性选择机制提供基础资料。
猜你喜欢
公民与法治(2023年2期)2023-03-31 03:18:44
特产研究(2022年6期)2023-01-17 05:05:06
水产养殖(2021年5期)2021-05-21 03:29:50
保健与生活(2018年17期)2018-01-27 15:35:10
法制博览(2018年34期)2018-01-22 18:21:33
四川动物(2017年4期)2017-07-31 23:54:19
今日文摘(2017年13期)2017-07-19 08:13:14
科学中国人(2015年26期)2015-03-12 03:17:05
集美大学学报(自然科学版)(2015年4期)2015-02-28 01:13:37
河北遥感(2014年3期)2014-07-10 13:16:48
