
布鲁氏菌Ⅳ型分泌系统蛋白BMEI0742基因的原核表达及功能分析
2018-07-18 02:39豆晓霞荆明龙李明奇王小凤陈创夫
江苏农业科学 2018年12期
豆晓霞,刘 洋,李 敏,荆明龙,李明奇,王小凤,陈创夫,张 辉
(石河子大学动物科技学院/动物疾病防控兵团重点实验室,新疆石河子 832000)
由布鲁氏菌(Brucella)引起的布鲁氏菌病(Brucellosis)是一种重要的人畜共患病,可造成动物流产、死胎,生殖器官不对称肿大,人类似流感状的周期性波浪热,严重威胁畜牧业的发展和公共卫生健康,给世界经济带来严重的损失[1]。布鲁氏菌是属于α-2变形菌家族的致病性革兰氏阴性菌[2],胞内寄生和复制是布鲁氏菌的主要毒力特征[3]。布鲁氏菌不仅可以通过消化道黏膜和呼吸道黏膜进入机体内,还可以通过皮肤、眼结膜等进入机体内,并被宿主体内的吞噬细胞吞噬[4],虽然布鲁氏菌可以在多数细胞内寄生,但是其仍具有宿主特异性,该菌主要寄生在巨噬细胞和胚胎滋养层细胞内[5]。布鲁氏菌属经典的6个种属分别为羊种布鲁氏菌(B.melitensis)、牛种布鲁氏菌(B.abortus)、猪种布鲁氏菌(B.suis)、犬种布鲁氏菌(B.canis)、绵羊种布鲁氏菌(B.ovis)、沙林鼠种布鲁氏菌(B.neotomae)[6-8],分别主要感染山羊和绵羊、牛、猪、狗以及啮齿类动物。最近又有4种布鲁氏菌被鉴定出来,包括从海豹分离的鳍脚类布鲁氏菌(B.pinnipediae),从海豚分离的鲸种布鲁氏菌(B.ceti)[9],从红狐、田鼠、土壤中分离的田鼠种布鲁氏菌(B.microti)[10-12],从乳腺移植病人分离的湖浪种布鲁氏菌(B.inopinata)[13]。尽管犬种布鲁氏菌和绵羊种布鲁氏菌感染动物,但人感染犬种布鲁氏菌很罕见,人感染绵羊种布鲁氏菌也未报道[14],而羊种布鲁氏菌和流产布鲁氏菌是人感染的主要病原菌。目前,人感染布鲁氏菌主要通过已经感染的动物(如绵羊,山羊,牛等一些家畜)以及动物肉制品、奶牛津液(如尿液和乳汁)、污染奶制备的奶制品等。人通过污染的环境(如气溶胶)以及伤口等途径也可造成布鲁氏菌病的传染。值得一提的是,人在感染布鲁氏菌平均3~4周后,初期会出现波浪热,尤其是食欲不振、浑身无力、盗汗、体质量减轻,偶尔还会出现头痛,后期会伴发关节疼痛等。而初期的症状和感冒类似,往往会被误认为感冒,以至于延误病情的治疗。因此,防控家畜布鲁氏菌病的发生也有利于减少人感染布鲁氏菌病的概率。
延伸因子(elongation factors,EF)是在mRNA翻译时促进多肽链延伸的蛋白质因子,在原核生物和真核生物中延伸的因子并不相同。原核延伸因子是原核细胞进行翻译时所需要的3种延伸因子,分别命名为EF-Tu、EF-Ts以及EF-G(其中EF-Tu和EF-Ts可以复合为EF-T)[15],原核延伸因子的化学本质都是蛋白质。布氏杆菌是一种胞内寄生菌,可在专业或非专业吞噬细胞内增殖。布氏杆菌在宿主细胞中的存活能力和繁殖能力对其毒力都是至关重要的,而加强病原菌与宿主相互作用分子机制的研究,是预防和控制布鲁氏菌感染的前提。最近研究表明,细菌EF-Tu蛋白是一种多功能蛋白,不仅仅是作为蛋白翻译因子起作用,而且更重要的是EF-Tu参与许多重要的细胞和疾病过程[16]。布氏杆菌EF-Tu在感染宿主细胞中作用的研究目前依然是空白。关于布鲁氏菌翻译延伸因子Tu的研究,仅见布鲁氏菌BMEI0742基因编码的翻译延伸因子Tu。本研究选取羊布鲁氏菌Ⅳ型分泌系统蛋白BMEI0742基因为研究对象,并构建其原核表达载体,检测其反应原性,分析其生物学功能,为探讨其在布氏杆菌侵入及胞内存活过程中的调控作用及其分子机制等奠定基础。
1 材料与方法
1.1 材料
1.1.1 菌株和质粒 羊种布鲁氏菌16M株、大肠杆菌(Escherichiacoli) DH5α菌株、BL21(DE3)菌株、pET-28a载体菌株由石河子大学动物科技学院/动物疾病防控兵团重点实验室保存;pMD19-T载体购自Promega公司。
1.1.2 试剂 羊种布鲁氏菌阳性血清由石河子大学动物科技学院/动物疾病防控兵团重点实验室保存;诱导剂IPTG购自Merck公司;限制性内切酶(BamH Ⅰ/XhoⅠ)、DNA Marker、T4连接酶购自大连宝生物工程有限公司;2×EsTaqMasterMix、细菌质粒小提取试剂盒以及琼脂糖凝胶回收试剂盒购自北京康为世纪生物科技有限责任公司;HRP标记的兔抗羊IgG、蛋白Marker购自北京康为世纪生物科技有限责任公司;2%蛋白质印迹(WB)无蛋白封闭液购于生工生物工程(上海)股份有限公司;其他试剂均为实验室常用分析纯度的产品。
1.1.3 仪器 洁净工作台(SW-CJ-2FD);PCR仪(Eppendorf,nexus GSX1);低温冷却液循环泵(DLSB,5L/S);超速离心机(Thermo,PIC021);恒温培养箱(DNP-9162);恒温振荡器(CRYSTAL,IS-RSV1);高速冷冻离心机(ThermoFisher);超声波细胞粉碎机(JY,88-IIN);水浴锅(XMTD-8222);聚丙烯酰胺凝胶电泳仪(DYY-8B);半干式电转仪(BIO-RAD);蛋白纯化仪(AKTA)。
1.2 方法
1.2.1 目的基因扩增 根据GenBank中登录的布鲁氏菌(NC-003317)BMEI0742基因序列设计引物,由北京六合华大基因有限公司合成,序列见表1。

表1 PCR引物序列
以羊种布鲁氏菌16M株的灭活菌液(85 ℃金属浴灭活 1 h)为模板,进行菌液PCR扩增,PCR反应体系(20 μL)见表2,反应条件为:95 ℃ 5 min;94 ℃ 40 s,64 ℃ 30 s,72 ℃ 1 min 10 s,30个循环;最后72 ℃ 1 min。反应结束后,PCR产物 4 ℃ 保存或直接进行1%琼脂糖凝胶凝胶电泳。回收目的基因片段,并将回收后目的片段与pMD19-T载体在16 ℃水浴条件下进行连接,连接体系(10 μL)见表3,构建重组质粒pMD19-T-BMEI0742。重组质粒转化至大肠杆菌DH5α感受态细胞,并在氨苄抗性的LB固体培养基上进行阳性筛选,进行菌液PCR和双酶切鉴定阳性克隆,并将提取的质粒送往生工生物工程(上海)股份有限公司进行测序。

表2 PCR反应体系

表3 连接体系
1.2.2 构建BMEI0742基因原核表达载体 将送公司测序正确的质粒pMD19-T-BMEI0742和pET-28a载体用限制性内切酶BamH Ⅰ和XhoⅠ 进行双酶切,经琼脂糖凝胶回收试剂盒回收目的片段和酶切后的线性pET-28a载体。将目的片段和线性pET-28a载体在T4连接酶的作用下16 ℃水浴连接,连接产物转化至大肠杆菌BL21(DE3)感受态细胞,经卡那抗性的LB固体培养基筛选阳性克隆,进行菌液PCR验证和酶切鉴定。
1.2.3 重组蛋白表达及表达条件的优化 将筛选出的阳性克隆菌在LB液体培养基中,37 ℃,200 r/min摇床中培养过夜,将培养过夜的菌液按1 ∶100的比例接种至新鲜的LB液体培养基中,于37 ℃、200 r/min条件下培养约1 h,使其D600 nm值为 0.4~0.6,在其他条件不变的情况下,按照不同的诱导剂(IPTG)浓度(终浓度为0.2、0.6、1.0 mmol/L)诱导不同时间(0、4、6、8 h)后收集菌体。取1 mL菌液离心后,弃掉上清,并在离心管中按照1 ∶4的比例加入经过稀释的1×SDS蛋白上样缓冲液,在振荡器上振荡使菌体和蛋白上样液充分混匀,在金属加热器上100 ℃加热10 min,取20 μL样品进行15%蛋白电泳,观察电泳结果。筛选目的基因的最佳表达条件,并以最佳的诱导表达条件进行大量诱导表达,采用AKTA蛋白纯化系统纯化后进行SDS-PAGE电泳鉴定。
1.2.4 重组蛋白的Western-Blot检测 筛选出最佳表达条件后,以最佳诱导条件诱导重组蛋白表达,并对该蛋白进行纯化,经SDS-PAGE后,将蛋白转移到NC膜上,以200 mA的电流转膜50 min。将膜放入预先用TBST缓冲液洗涤后的杂交瓶中,37 ℃封闭90 min,用TBST缓冲液洗涤NC膜3次,每次7 min。加入1 ∶500比例稀释的、经石河子大学动物疾病防控兵团重点实验室鉴定的羊种布鲁氏菌阳性血清,37 ℃孵育2 h,用TBST缓冲液洗涤NC膜3次,每次7 min,然后加入 1 ∶5 000 比例稀释的辣根过氧化氢酶标记的兔抗绵羊IgG,37 ℃孵育1 h,TBST缓冲液洗涤NC膜3次,每次7 min。避光条件下,DAB显色剂显色后拍照。
1.3 重组蛋白的生物信息学分析
1.3.1 跨膜结构分析 通过TMHMM Server v.2.0在线软件分析(http://www.cbs.dtu.dk/services/TMHMM/)羊种布鲁氏菌的BMEI0742蛋白氨基酸序列的跨膜结构。
1.3.2 信号肽分析 利用SignalP 4.1 Server在线软件预测(http://www.cbs.dtu.dk/services/SignalP/)羊种布鲁氏菌的BMEI0742蛋白的信号肽。
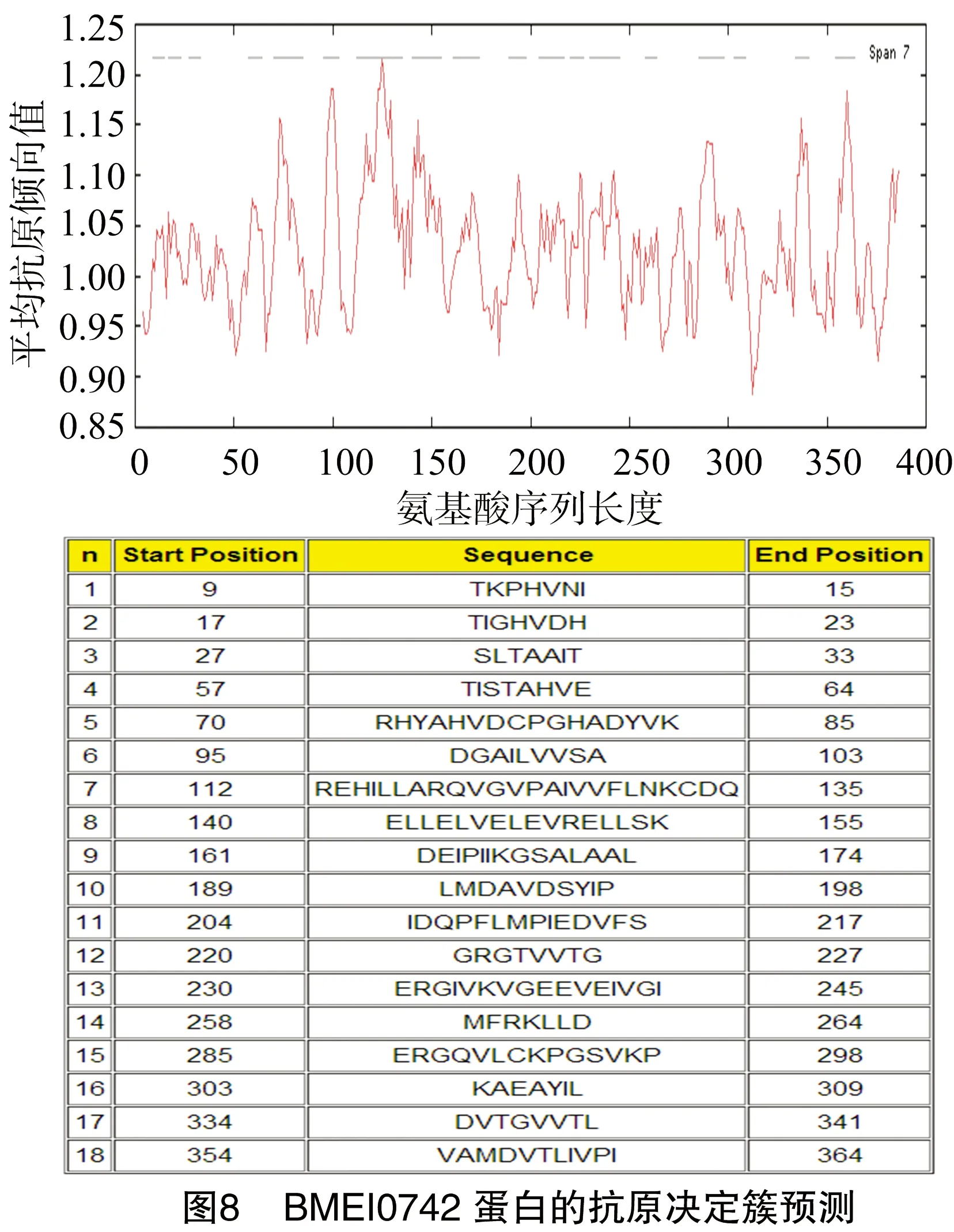
1.3.3 抗原决定簇的预测 利用Predicting Antigenic Peptides在线软件预测(http://imed.med.uce.es/Tools/antigenic.pl)羊种布鲁氏菌的BMEI0742蛋白的抗原决定簇。
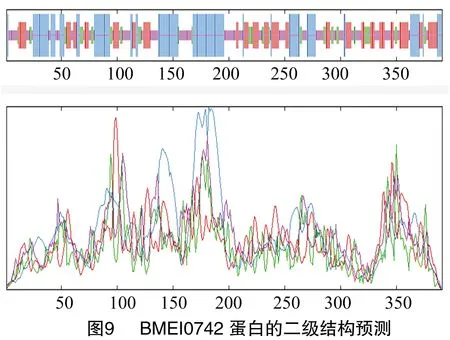
1.3.4 蛋白的二级结构预测 通过SOPMA在线软件分析(https://npsa-prabi.ibcp.fr/cgi-bin/npsa-automat.pl?page=/NPSA/npsa-sopma.html)羊种布鲁氏菌的BMEI0742蛋白的二级结构。
1.3.5 蛋白的三级结构预测 通过Phyre2在线服务器预测(http://www.sbg.bio.ic.ac.uk/phyre2/html/page.cgi?id=index)羊种布鲁氏菌的BMEI0742蛋白的三级结构。
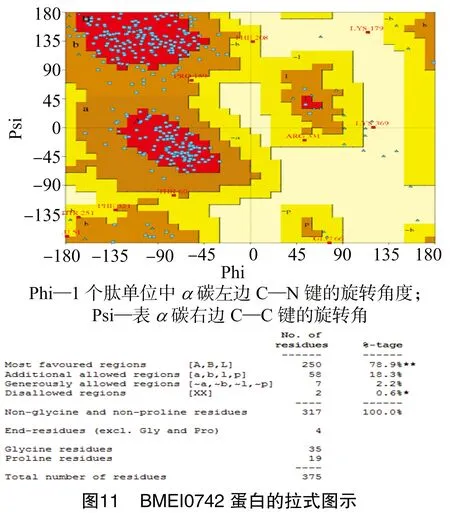
1.3.6 蛋白的3D模型评估 利用PDBsum Generate在线评估(http://www.ebi.ac.uk/thornton-srv/databases/pdbsum/Generate.html)羊种布鲁氏菌的BMEI0742蛋白的三级结构模型。
2 结果与分析
2.1 目的基因的扩增及酶切鉴定
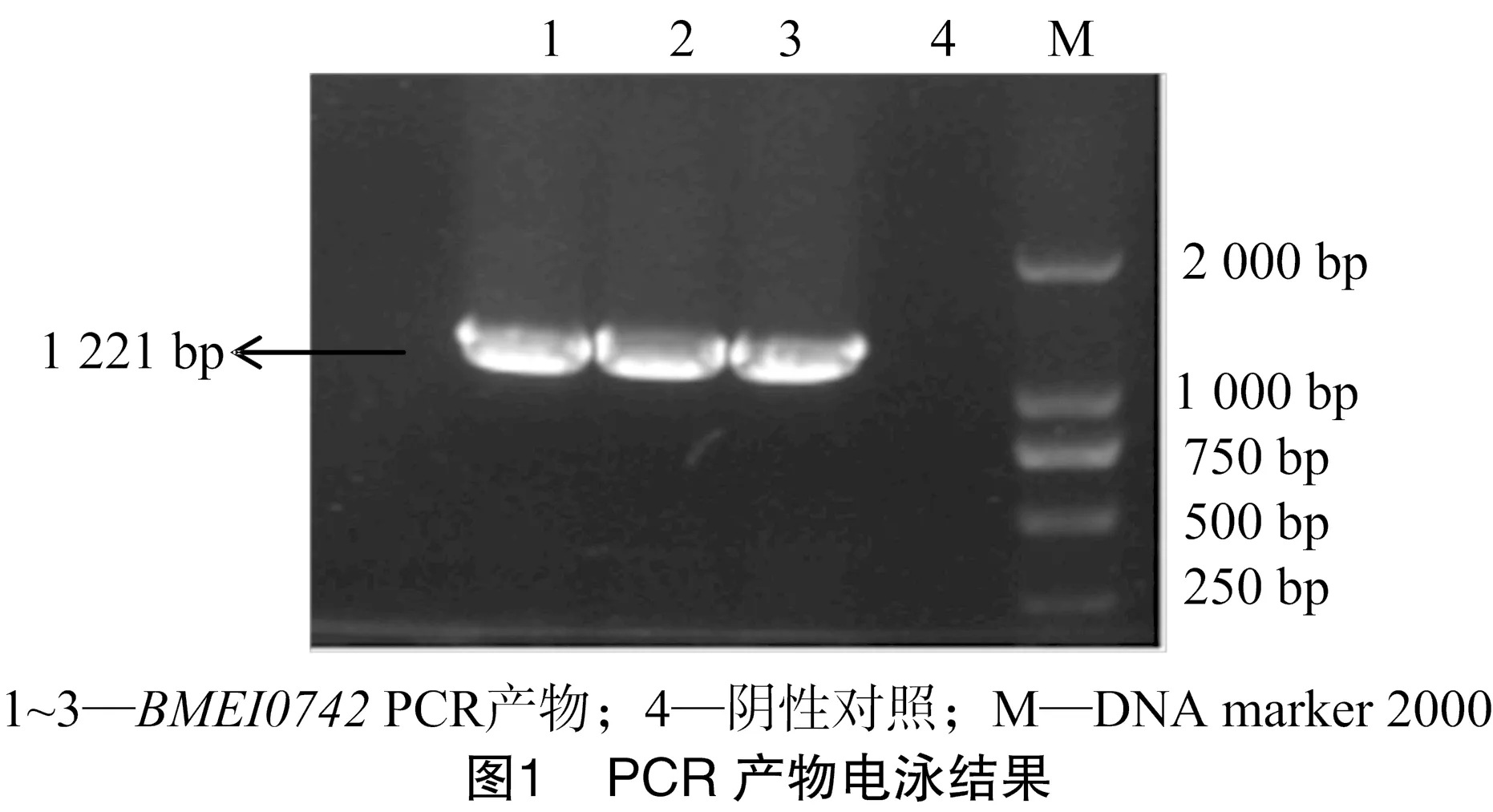
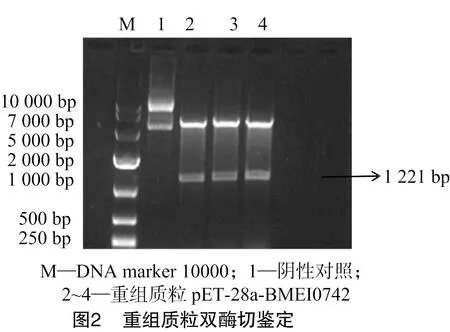
NCBI上羊种布鲁氏菌的BMEI0742基因的片段大小为 1 221 bp,PCR扩增产物经1%琼脂糖凝胶电泳分析,与实际大小一致,结果如图1所示。用相应的限制性内切酶(BamH Ⅰ/XhoⅠ)进行双酶切后,用1%琼脂糖凝胶电泳进行检测。结果显示,得到大小与预期结果相符的基因条带,表明羊种布鲁氏菌Ⅳ型分泌系统蛋白BMEI0742基因已经成功插入 pET-28a 表达载体中(图2)。
2.2 重组蛋白表达条件的优化
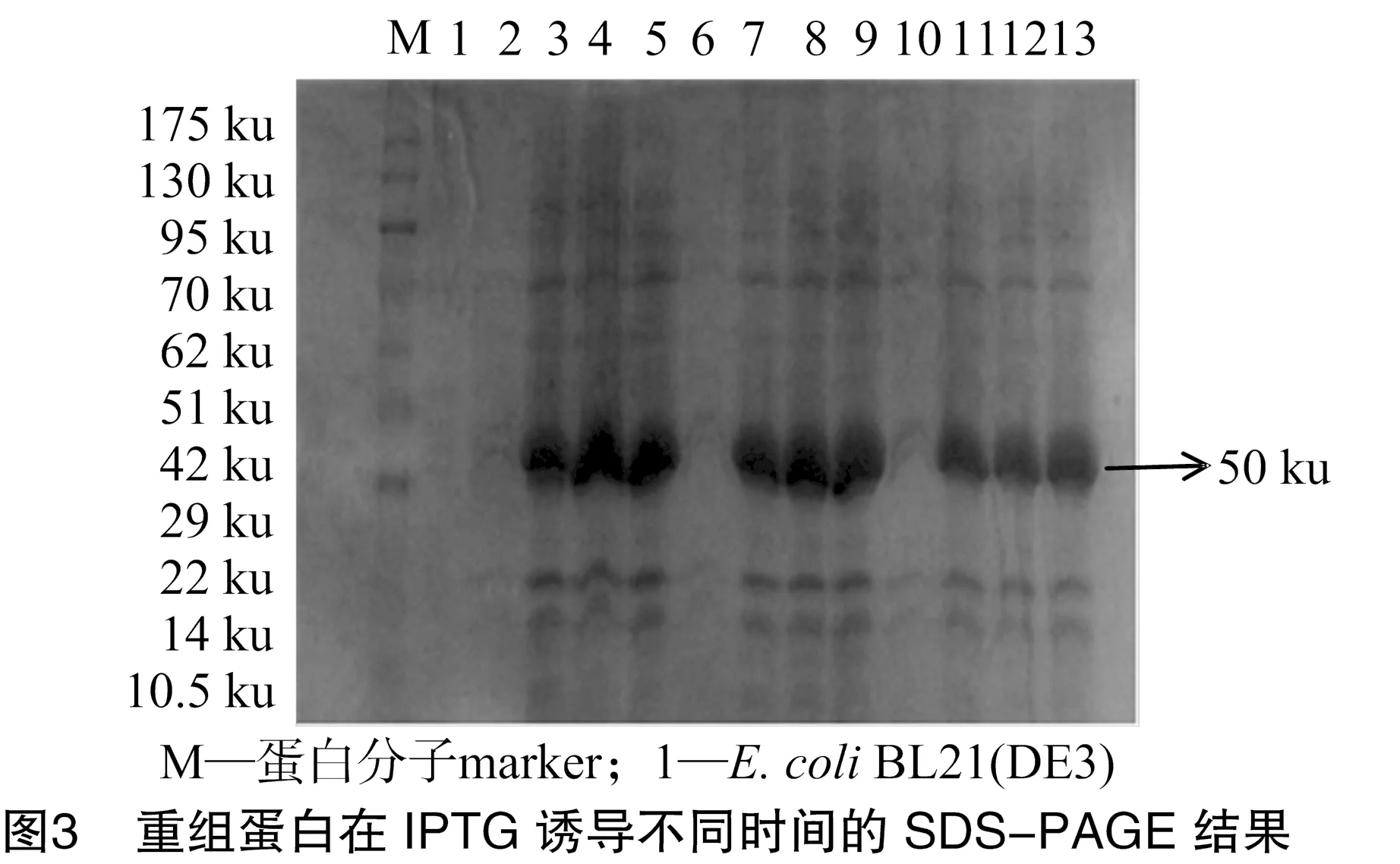
为寻找重组蛋白表达的最佳诱导时间和诱导剂浓度,分别在诱导剂浓度不变的前提下,设置时间梯度筛选最佳诱导时间;在诱导时间不变的前提下,设置诱导剂浓度梯度筛选最佳诱导剂浓度,进而筛选出最佳的诱导条件。结果显示,诱导剂IPTG的终浓度为1 mmol/L,诱导8 h时重组蛋白表达量最高,结果如图3所示。蛋白大量诱导表达后经AKTA蛋白纯化系统纯化,SDS-PAGE分析表明,纯化后的蛋白条带清晰,无杂带,表明纯化效果较好(图4)。

2.3 Western-Blot检测
对诱导8 h后的重组蛋白纯化后进行Western-Blot检测,200 mA恒流电流下转膜50 min,经观察,可见明显的阳性条带,说明诱导得到的重组蛋白能被HRP标记的兔抗绵羊IgG抗体识别,且大小与pET-28分子质量加上目标分子蛋白质量的重组蛋白总分子质量相符,结果如图5所示。

2.4 目的蛋白的生物信息学分析
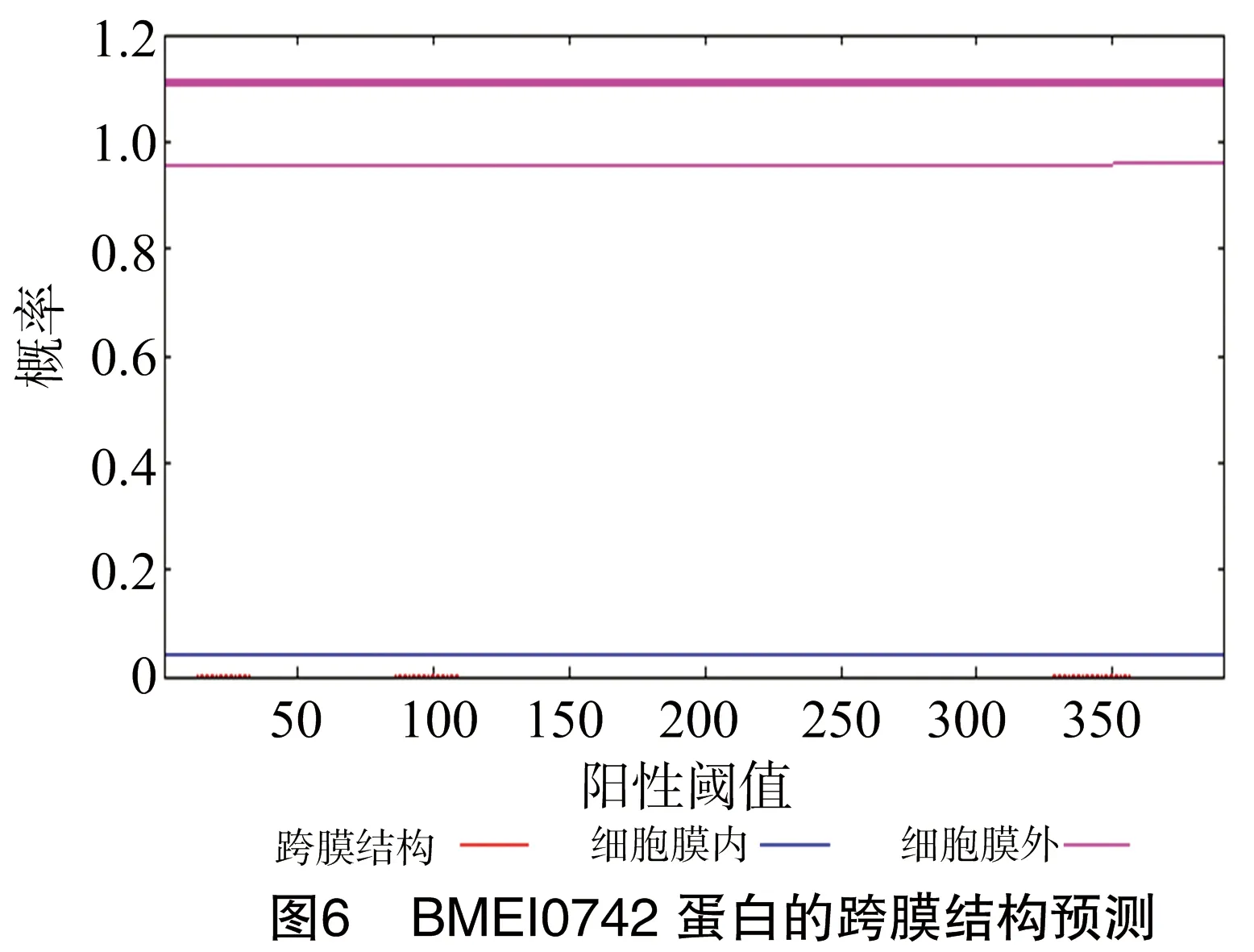
通过TMHMM Server v.2.0在线软件分析得出,BMEI0742蛋白没有跨膜结构区(图6)。通过SignalP 4.1 Server在线预测得出,BMEI0742蛋白没有信号肽(图7)。经Predicting Antigenic Peptides在线软件得出,BMEI0742蛋白有18个抗原决定簇(图8)。
经SOPMA在线软件分析得出,BMEI0742蛋白中有127个氨基酸参与形成α-螺旋,占所有氨基酸的32.48%;99个氨基酸参与延伸链的形成,占所有氨基酸的25.32%;45个氨基酸参与β-折叠的形成,占所有氨基酸的11.51%;另外,还有120个氨基酸参与无规卷曲结构的形成,占所有氨基酸的30.69%(图9)。通过Phyre2在线服务器分析预测,BMEI0742蛋白的三级结构如图10所示。经PDBsun Generate在线评估,在BMEI0742蛋白的3D同源模型的Ramachandran图中可以看出,375个氨基酸残基中有250个处于核心允许区,占78.9%;58处于额外允许区,占 18.3%;7个处于最大允许区,占2.2%;同时,也存在0.6%的氨基酸位于不允许区域中,这说明该模型具有一定的可信度,结果如图11所示。


3 讨论与结论
布病在全球170多个国家和地区流行,主要分布于地中海、非洲、拉丁美洲、中东和亚洲的部分地区[17]。我国各省(市、区)均有人、畜不同程度的布病流行。随着经济的发展,布病已经开始从牧区转向工业化或者是人口密集的经济发达地区蔓延。同时,商务和休闲旅游等产业的发展、人口流动的增加促使布病非疫区新发病例呈上升态势[18]。
生物信息学是利用计算机作为辅助工具,对生物信息数据进行整理归纳,探索生物奥秘的科学,它主要体现在基因组学和蛋白组学两大领域。布鲁氏菌基因组的测序与分析为揭示布鲁氏菌生存机制、分类与进化、毒力演化、疫苗株的弱化机制等提供有力支持,为阐述布鲁氏菌毒力相关基因的演化历史提供依据。自2002年羊种布鲁氏菌16M参考基因组测序完成以来,已经有300多株布鲁氏菌基因组完成了测序,其中有16株基因组已经完成了精细物理图谱的绘制[19]。随着蛋白质组学的发展,羊种布鲁氏菌和牛种布鲁氏菌蛋白质组学的研究都有不少报道,包括全菌蛋白质组学[20]、膜蛋白质组学[21-22]、免疫蛋白质组学[23-24]、分泌蛋白质组[3]等都有研究。本研究对BMEI0742蛋白进行了生物信息学分析,有助于了解蛋白质的功能,为布鲁氏菌的致病机制研究作基础。
细菌EF-Tu是翻译过程中重要的蛋白因子,也是含量最多的蛋白之一,在原核生物中广泛存在[25],且高度保守,被广泛用来进行种系发生分析[26]。最近的研究表明,EF-Tu是一种重要的多功能蛋白,不仅仅是作为蛋白翻译因子起作用,而且更重要的是EF-Tu参与许多重要的细胞和疾病过程,包括信号传导、翻译控制、凋亡、细胞骨架组成等。EF-Tu 没有信号肤却能联合其他一些缺乏分泌信号的蛋白以类似外吐小体(exosome)的运输方式分泌并干扰宿主细胞信号传递[27]。EF-Tu蛋白自身也具有较强的免疫原性,在细菌中高度保守,且该蛋白在其他多个种属细菌中已有关于作为鉴别诊断和候选亚单位疫苗的潜力的报道[28]。在绵羊布氏杆菌疫苗株M5-90致弱分子机制的研究过程中发现,EF-Tu具有很强的免疫原性,而重组的EF-Tu蛋白免疫小鼠后可以部分抵抗强毒株羊种布鲁氏菌M28的攻击[29]。显然,EF-Tu在多种细菌中均具有诱导宿主免疫应答的能力。这说明EF-Tu蛋白在细菌感染宿主的过程中,作为一种重要的免疫原被宿主细胞所识别和捕获,可能在其致病过程中发挥重要作用。
经过BMEI0742蛋白的生物信息学分析显示,BMEI0742蛋白没有跨膜结构,不能进行胞内外信号转导等生命活动。BMEI0742蛋白没有信号肽,说明BMEI0742蛋白属于非分泌型蛋白。BMEI0742蛋白拥有一定数量的抗原表位,表明BMEI0742蛋白作为抗原能引起良好的免疫应答反应。BMEI0742蛋白的二级结构是以α-螺旋为主,都是通过骨架上的羟基和酰胺集团之间形成的氢键维持二级结构的稳定。本研究成功构建了BMEI0742蛋白的三维模型,该模型具有一定的可信度,有利于蛋白的结构和功能的可视化分析,具体的生物学功能需要进一步的试验验证。
总之,布鲁氏菌的病原与宿主的相互作用是一个非常复杂的过程。本研究通过构建布鲁氏菌BMEI0742基因的原核表达载体并对其反应原性进行鉴定以及生物信息学分析,为进一步研究EF-Tu在布鲁氏菌感染宿主细胞过程中所发挥的生物学功能提供依据。
猜你喜欢
数学物理学报(2021年4期)2021-08-30
现代畜牧科技(2021年4期)2021-07-21
新世纪智能(数学备考)(2020年10期)2021-01-04
乡村科技(2020年1期)2020-04-20
中国动物检疫(2018年7期)2018-07-04
灾害医学与救援(电子版)(2018年1期)2018-06-05
中成药(2017年12期)2018-01-19
中国交通信息化(2017年8期)2017-06-06
中国畜禽种业(2017年10期)2017-01-15
中国民族医药杂志(2016年9期)2016-05-09
