
网箱和流水池塘养殖俄罗斯鲟的生长特性
2018-07-06 03:29许式见邵辉胡谋石灏夏永涛张颖孙大江
水产学杂志 2018年3期
许式见 ,邵辉 ,胡谋 ,石灏 ,夏永涛 ,张颖 ,孙大江
(1.杭州千岛湖鲟龙科技股份有限公司,浙江 杭州 311701;2.中国水产科学研究院黑龙江水产研究所,黑龙江 哈尔滨 1500703)
目前,我国已是世界上最大的鲟养殖与鱼子酱生产国。我国鲟年养殖量达9万t以上,占世界鲟产量的80%以上[1]。鲟鱼子酱的年出口量约70t,占世界鲟养殖鱼子酱产量的30%左右,经济效益显著[2]。俄罗斯鲟Acipenser gueldenstaedtii隶属于鲟形目Acipenseriformes鲟科Acipenseridae鲟属Acipenser,主要栖息在里海和亚速海——黑海水系[3]。于1993年引入我国,因其具有生长快、性成熟早、抗病力强及鱼子酱价值高等优点养殖发展迅速,约占我国鲟产量的5.42%[4,5]。迄今为止,有关俄罗斯鲟的研究主要集中在俄罗斯鲟鱼肉加工、精子低温保存、遗传育种及养殖技术等研究,而关于其生长特性的研究尚未见报道[6-8]。
本文通过分析1~10龄俄罗斯鲟的生长特性及生长模型的拟合情况,建立不同年龄的生长拟合模型,以了解人工养殖条件下俄罗斯鲟的生长特征,旨在揭示人工养殖条件下俄罗斯鲟的生长特征和规律,并为科学地进行鲟养殖及生产提供参考。
1 材料与方法
1.1 材料
2017年5 月,于杭州千岛湖鲟龙科技股份有限公司养殖基,随机选取1~10龄个体,各50尾进行测定,测量体长和体质量。其中,1~7龄为网箱养殖(6m×6m×7.5m),8~10龄为圆形流水池塘养殖(Φ10m),年养殖水温变化范围为8~25℃。
1.2 生长指数的计算
采用Microsoft Excel 2010和SPSS Statistics 22软件对所得数据进行生长指数计算,计算公式如下:体长与体质量的关系W=aLb;绝对增长量:L2-L1和 W2-W1;相对增长率 η= (L2-L1)/L1×100 和(W2-W1)/W1×100;生长常数 P=(lnL2-lnL1)×(t2+t1)/2 和 (lnW2-lnW1)×(t2+t1)/2;生长指标R=(lnL2-lnL1)×L1和(lnW2-lnW1)×W1;特定生长率SGR=(lnL2-lnL1)/(t2-t1)×100和(lnW2-lnW1)/(t2-t1)×100;变异系数 CV=100×SD/X;肥满度CF=W/L3×105。式中,L为体长(cm),W为体质量(kg);L1、L2和 W1、W2分别为时间 t1、t2时的体长(cm)和体质量(kg),SD为标准差,X为平均体质量(kg),a和 b为常数。
1.3 生长模型
俄罗斯鲟体长和体质量生长曲线拟合采用VBGF、幂函数、指数、对数和线性5种模型,并以AIC(Akaike’s Information Criterion)为选择最佳模型的依据。
2 结果与分析
2.1 不同年龄组俄罗斯鲟的体长生长
如表1所示,1~10龄俄罗斯鲟的体长随着年龄的增长而增加,9龄体长平均值最长,为(120.1±3.7)cm。体长绝对增长量、相对增长率、特定生长率、生长常数和生长指数总体变化趋势均表现为升高-降低-升高-降低,4龄时达到最大。养殖条件下俄罗斯鲟体长的变异系数范围为2.93%~8.46%,1龄鱼的体长变异系数最大,2~10龄的体长变异系数波动幅度均较小。单因素方差分析显示,年龄对体长的影响极显著(P<0.01),Duncan’s新复极差法比较表明1~8龄各鱼龄间体长差异均极显著(P<0.01),8龄和10龄、9龄和10龄体长差异均不显著(P>0.05),8 龄和 9龄体长差异显著(P<0.05)。因此,俄罗斯鲟体长的快速生长期为1~4龄,稳定生长期为5~8龄,9~10龄生长缓慢。
2.2 不同年龄组俄罗斯鲟的体质量生长变化
俄罗斯鲟不同年龄的体质量见表2。1~10龄俄罗斯鲟的体质量随年龄的增长而增加,9龄体质量的平均值最大值,为(20.28±2.38)kg。其年体质量绝对增长量分别在4龄与8龄时达到两个最大值,分别为4.27kg和4.65kg。与其年体质量绝对增长率的变化规律不同,俄罗斯鲟的体质量相对增长率和特定生长率在2龄时最大,分别为139.55%和87.36%;生长常数在4龄达最大值(2.66);生长指数在8龄达最大值(4.06)。此外,人工养殖条件下,俄罗斯鲟的体质量变异系数范围为9.05%~25.91%,1龄鱼体质量的变异系数最大,2~10龄的体质量变异系数波动幅度较小,变动范围为9.05%~16.57%。单因素方差分析显示,年龄对体质量的影响极显著(P<0.01),Duncan’s新复极差法比较表明 1~8 龄各鱼龄间体质量差异均极显著(P<0.01),8~10龄各年龄间体质量差异均不显著(P>0.05)。因此,俄罗斯鲟体质量的快速生长期为1~4龄,稳定生长期为5~8龄,9~10龄后生长减慢。
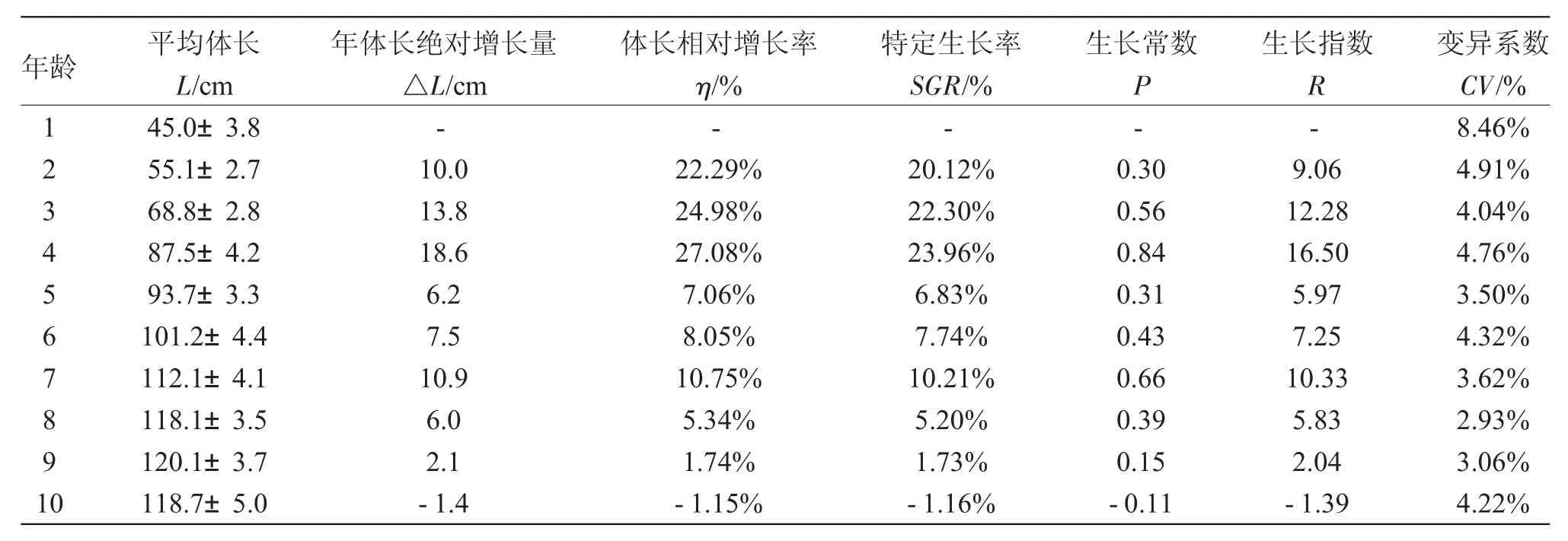
表1 不同年龄的俄罗斯鲟体长生长变化Tab.1 The growth in body length in Russian sturgeon A.gueldenstaedtii at various ages
2.3 俄罗斯鲟年龄与体长、体质量及肥满度的关系
俄罗斯鲟年龄与体长、体质量的生长曲线见图1和图2。如图1所示,9龄时俄罗斯鲟的体长与体质量生长曲线达最大值后开始降低(图1)。俄罗斯鲟的体长增长曲线在4龄和7龄时出现了2个高峰期,而体质量增长高峰期则为4龄和8龄(图2),表明1~4龄为俄罗斯鲟的生长快速期,5~8龄为稳定生长期,而9~10龄后则进入生长衰老期。
人工养殖条件下,俄罗斯鲟的体长与体质量呈幂函数相关(W=aLb),W=6×10-6L3.1292(R2=0.9846,n=500)(图3)。其中幂函数指数 b>3,表明养殖条件下俄罗斯鲟的生长系正异速生长。俄罗斯鲟的肥满度在1龄时增长迅速,之后随着年龄的增加趋于平稳变化(图4)。
2.4 生长方程
5种方程式对俄罗斯鲟体长与体质量生长的拟合参数、决定系数R及RSS统计量见表3。5种方程式中,VBGF和对数2种生长方程对体长生长的拟合效果接近,拟合度最好的为VBGF生长方程,AIC值最小;指数方程式拟合度最低。VBGF体长生长方程为Lt=130.776(1-e-0.269(t-0.122))(R2=0.9485,n=500)。体质量生长拟合度最好的为VBGF生长方程,AIC值最小,体质量生长方程为Wt=35.709(1-e-0.112(t-1.905))(R2=0.8781,n=500)(表3)。计算表明,拐点年龄为4.36龄,拐点处W=8.59 kg,L=90.52cm。
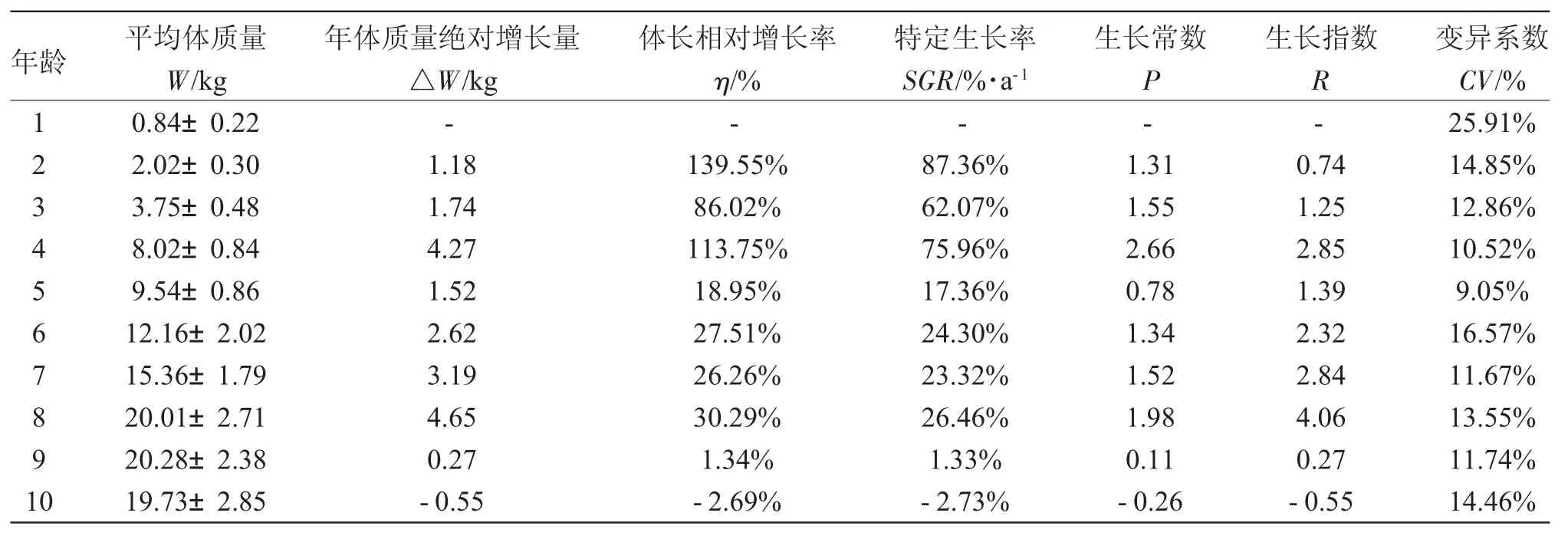
表2 不同年龄的俄罗斯鲟体质量生长变化Tab.2 The growth in body weight in Russian sturgeon A.gueldenstaedtii at various ages
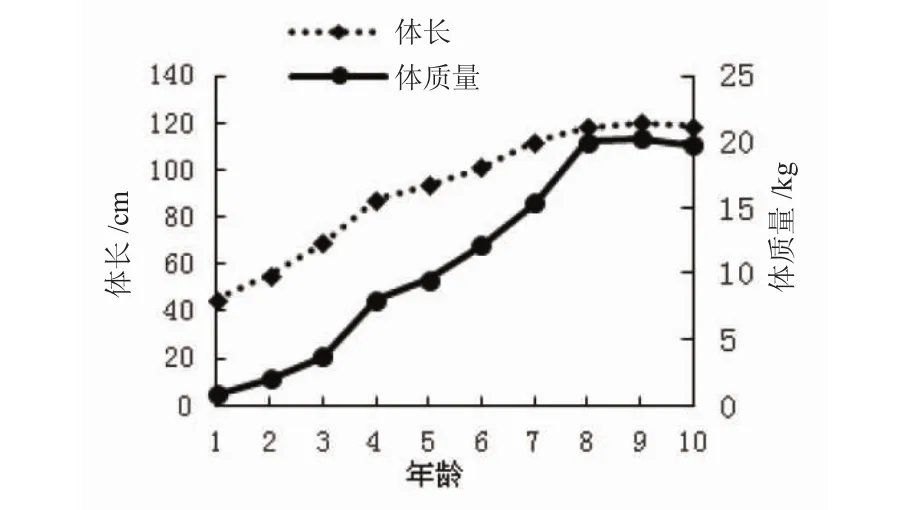
图1 俄罗斯鲟年龄与体长、体质量的关系Fig.1 The relation of the age,body length and weight growth of A.gueldenstaedtii
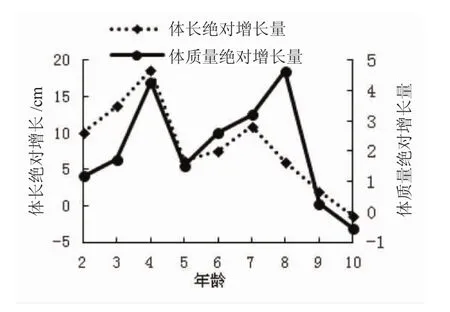
图2 俄罗斯鲟年龄与体长和体质量增长关系Fig.2 The relation of the age,length growth body weight of A.gueldenstaedtii
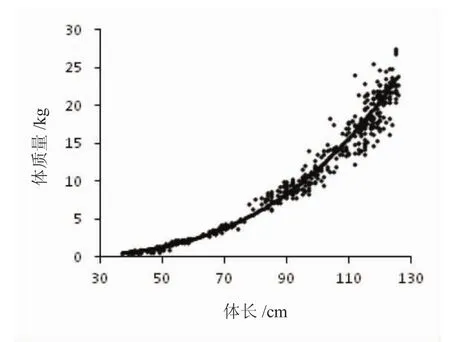
图3 俄罗斯鲟体长与体质量的关系Fig.3 The relation between body length and weight of A.gueldenstaedtii
图4 不同年龄俄罗斯鲟的肥满度Fig.4 The relation between age and CF of A.gueldenstaedtii
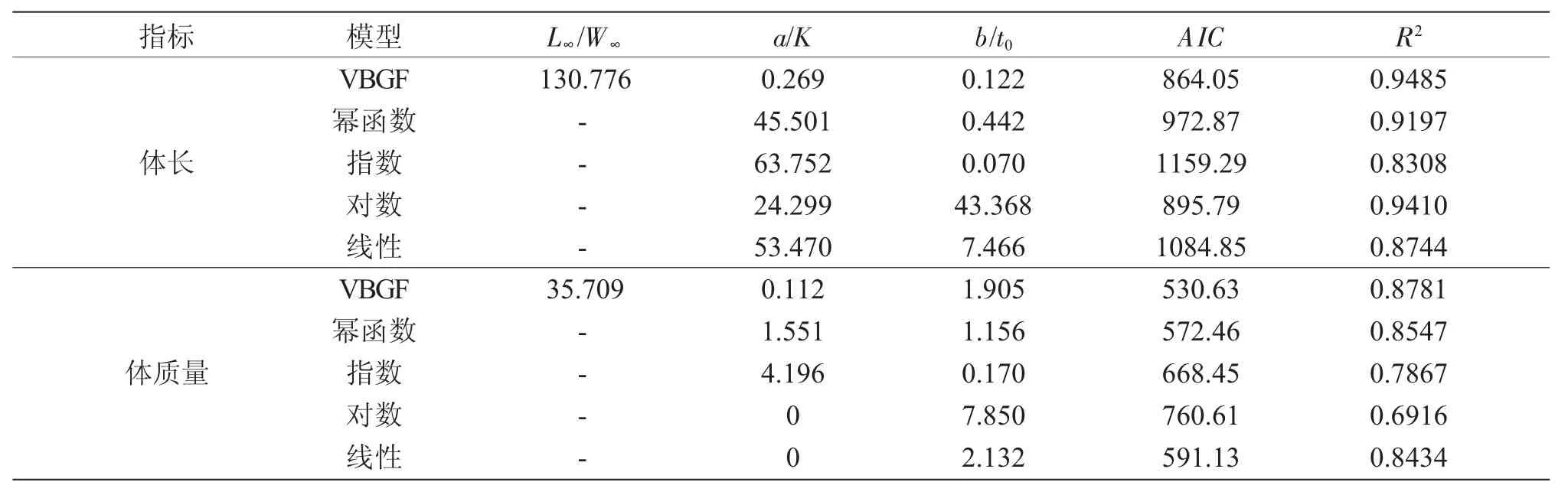
表3 俄罗斯鲟体长和体质量生长方程的拟合数据Tab.3 The parameters and fitness of functions describing the growth in Russian sturgeon A.gueldenstaedtii
3 讨论
鲟为大个体珍稀淡水鱼类,其味道鲜美、营养丰富,尤其鲟鱼子酱被誉为“黑色黄金”,具有极高的经济价值与营养价值[9]。鱼类的生长规律对鱼类开发利用具有重要的指导意义[10]。鲟寿命长,可达二十至上百年,而我国鲟养殖仅有近二十年的养殖历史,缺乏鲟生长性能的相关数据。与人工养殖杂交鲟的生长性能不同,本研究中1~10龄俄罗斯鲟的体质量增长速度呈先上升后下降的特点,体长增长速度则逐渐减小[11]。鱼类的体长与体质量关系是评价鱼类种质质量和养殖效果的重要标准之一[7]。与中华鲟Acipenser sinensis和白鲟A.transmontanus亲鱼的b值均大于3相同,1~10龄俄罗斯鲟的b值也大于3[12,13]。
研究表明,不同种鲟的肥满度与其生长阶段、养殖状况等相关[14,15]。养殖条件下,1~10龄俄罗斯鲟的肥满度变化在0.44~0.58之间,高于0~3龄达氏鳇杂交种(0.37~0.39)、西伯利亚鲟 A.baerii(0.35~0.58)、波斯鲟 A.persicus(0.61~0.65)和俄罗斯鲟A.gueldenstaedtii(0.37~0.38)等幼鲟的肥满度(0.35~0.65)变幅区间[14,15]。而与西伯利亚鲟亲鱼的肥满度(0.44~0.63)变幅区间相似[15]。
目前,VBGF生长方程已广泛地应用于鲟生长性能的评价[11,17-22]。对不同年龄、不同养殖环境鲟生长性能的研究表明,VBGF生长方程对鲟的生长特征描述结果存在种属、遗传、生理及生境差异[16-22]。对野生湖鲟与养殖杂交鲟的体长与年龄研究表明,湖鲟与杂交鲟的体长与年龄较为符合VBGF生长模型,这与本研究结果基本一致[11,23]。此外,多种生长模型对杂交鲟与短吻鲟A.brevirostrum的体长和体质量生长的描述结果表明,Ln(体长0.94,体质量0.97)、多项式 (0.95,0.97)、VBGF (0.93,0.97)和Gompertz(0.87,0.87)中,体长和体质量生长的 Gom-pertz方程式的R2值均小于其他3种方程式[11,18]。本研究中,VBGF(0.95,0.88)和幂函数(0.92,0.85)对1~10龄俄罗斯鲟体长与体质量的拟合R2值高于logistic(-8.4433,0.88)、指数 (0.83,0.78)、对数(0.94,0.69)与线性(0.87,0.84)方程式。
[1]水生野生动物保护分会.中国鲟鱼养殖与利用调查报告[J].水野协(2016)51 号.
[2] Elisa Boscari,Anna Barmintseva,Shuhuan Zhang,et al.Genetic identification of the caviar-producing Amur and Kaluga sturgeons revealed a high level of concealed hybridization[J].Food Control,2017,82:243-250.
[3] Celikkale M S,Memis D,ErcanE,et al.Growth performance of juvenile Russian sturgeon (Acipenser gueldenstaedti Brandt&Ratzenburg,1833)at twostocking densities in net cages[J].Zeitschrift fur Angewandte Ichthyologie,2005,21(1):14-18.
[4]林克文,姜存楷.俄罗斯鲟(Acipenser gueldenstaedti Brandt)引种养殖技术[J].现代渔业信息,2002,17(6):22-24.
[5]高露姣,夏永涛,黄艳青,等.俄罗斯鲟鱼卵与西伯利亚鲟鱼卵的营养成分比较[J].海洋渔业,2012(1):57-63.
[6]樊燕,侯虎,孙晨阳,等.加工方式对俄罗斯鲟鱼肉水分状态的影响[J].中国食品学报,2017(6):97-102.
[7]黄晓荣,张涛,冯广朋,等.超低温冷冻对俄罗斯鲟精子形态结构的影响[J].海洋渔业,2017(6):640-648.
[8]罗坤,夏永涛,王斌,等.俄罗斯鲟早期生长性状遗传参数的估计[J].中国水产科学,2015,22(3):426-432.
[9]孙大江,曲秋芝,张颖,等.中国的鲟鱼养殖[J].水产学杂志,2011,24(4):67-70.
[10]刘士瑞.浅色黄姑鱼发育、生长及呼吸代谢的研究[D].上海:上海水产大学,2007.
[11]张颖,刘晓勇,曲秋芝,等.养殖条件下施氏鲟(♂)×达氏鳇(♀)杂交后代的生长特性[J].水产学杂志,2013,26(2):1-8.
[12]邓中粦,余志堂,许蕴玕.中华鲟年龄鉴别和繁殖群体结构的研究[J].水生生物学报,1985,9(2):99-110.
[13]马骏,邓中粦,邓昕,等.白鲟年龄鉴定及其生长的初步研究[J].水生生物学报,1996,20(2):150-159.
[14] Cristina S B,Pãsãrin D&Simeanu.The study of some morphological characteristics of the sturgeon species of Polyodon spathula in different development stages[J].U-niversitatea de tiin e Agricole i Medici n ãVeterinarãIa i,2010:243-247.
[15]Sokolov L I,Koshelev O V&Khalatyan G I.Ecomorphological characteristics of Siberian sturgeon,Acipenser baeri,fromriver[J].Vopr Ikhtiol,1986,5:741-749
[16] Gisbert E&Williot P.Larval behaviour and effect of the timing of initial feeding on growth and survival of Siberian sturgeon(Acipenser baeri)larvaeundersmallscalehatcheryproduction[J].Aquaculture,1997,156(1):63-76.
[17]Shuman D A,Klumb R A,Wilson R H,et al.Pallid sturgeon size structure,condition,and growth in the Missouri River Basin[J].J Appl Ichthyol,2011,27(2):269-281.
[18] Ryan J W.Age,growth and recruitment of Hudson river shortnose sturgeon(Acipenser brevirostrum)[M].Maryland:UniversityofMaryland,2005.
[19]Fisheries B C.Fraser river white sturgeon monitoring program[R].Comprehensive report(1995 to 1999),Final Report Prepared for BC Fisheries.RL&L.Report No.815F:92.
[20]Scott R E,Dennis L S,Greg J P,et al.Comparison of age and growth of shovelnose sturgeon in the Missouri and Yellowstone Rivers[J].North American Journal ofFisheries Management,2003,23(1):230-240.
[21] Kergg M Smith&Edward A Baker.Characteristics of spawning lake sturgeon in the upper Black River,Michigan [J].North American Journal of Fisheries Management,2005,25(1):301-307.
[22]何斌,陈先均,杜军,等.人工养殖条件下达氏鲟生长特性的研究[J].西南农业学报,2011,24(1):335-339.
[23]David L G N,William F H B&Rossiter A.Conservation implications of behaviour and growth of the lake sturgeon,Acipenser fulvescens,in northern Ontario[J].Environmental BiologyofFishes,1999,55(1-2):135-144.
猜你喜欢
小猕猴智力画刊(2022年9期)2022-11-04
天天爱科学(2022年9期)2022-09-15
中学生数理化·七年级数学人教版(2022年5期)2022-06-05
昆明医科大学学报(2022年1期)2022-02-28
中学生数理化·七年级数学人教版(2021年5期)2021-11-22
新世纪智能(数学备考)(2020年12期)2020-03-29
小学生作文选刊·低年级版(2017年2期)2017-03-06
电子技术与软件工程(2016年20期)2016-12-21
小学生导刊(低年级)(2016年8期)2016-09-24
河北渔业(2015年10期)2015-10-15
