
半滑舌鳎繁育群体的遗传组成及家系配组策略
2018-07-06 03:26李超鲁翠云曹顶臣郑先虎程磊李庆华薛志勇孙效文
水产学杂志 2018年3期
李超 ,鲁翠云 ,曹顶臣 ,郑先虎 ,程磊 ,李庆华 ,薛志勇 ,孙效文
(1.中国水产科学研究院黑龙江水产研究所,淡水鱼类育种国家地方联合工程实验室,黑龙江 哈尔滨 150070;2.海阳市黄海水产有限公司,山东 海阳 265122)
半滑舌鳎Cynoglossus semilaevis分布于黄、渤海地区的底层大型经济鱼类,具有个体大、生长快、肉味鲜美等特点,深受消费者喜爱,逐渐成为我国海水养殖的重要种类。半滑舌鳎雌、雄个体生长速度相差悬殊,同龄个体雌性较雄性生长快30%~100%[1],鉴定性别特异分子标记[2-4],利用雌核发育[5,6]、多倍体[7-9]等技术开展全雌性半滑舌鳎苗种培育成为研究热点。但是,半滑舌鳎雌性异型性染色体(ZW)[10]的形式使其单性育种过程较雄性异型性染色体(XY)鱼类更为复杂,无法获得成活的“超雌”鱼(WW),导致半滑舌鳎全雌鱼的培育进展缓慢[11,12]。
家系选育和群体选育是常用的鱼类育种方法。陈松林等[13,14]通过家系间生长及抗病等性能的测试,获得了生长速度快、抗病力强的家系。近几年,鱼类分子育种发展迅速,结合传统育种方法开发出的基于亲本遗传背景的分子育种技术方案,在避免近亲繁殖、优化群体遗传结构、维持群体遗传多样性等方面效果显著,已应用于镜鲤Cyprinus carpio var.specularis、大黄鱼Larimichthys crocea等鱼类的选育中[15-18]。黎中宝等[19]研究表明,半滑舌鳎的2个养殖群体的平均多态位点比例(37.85%)较2个野生群体(42.57%)降低了12.47%。面对日益萎缩的野生资源,在半滑舌鳎选育中注意保护群体基因库的丰度对种质的持续利用十分必要。近十年来,半滑舌鳎已完成共显性标记[20]、高密度图谱及经济性状相关QTL[21,22]等遗传资源的储备,具备了开展分子育种的基础。本研究利用20个微卫星标记分析半滑舌鳎亲鱼群体的遗传组成,检测雌雄亲本间的遗传距离分布,确定半滑舌鳎家系配组雌、雄遗传距离的最佳“阈值”范围,根据统计结果制定半滑舌鳎的配组策略。
1 材料与方法
1.1 材料
半滑舌鳎亲鱼样品采自海阳市黄海水产有限公司。选择体表健康无损伤、体色纯正无白化、性腺发育较好的3龄后备亲鱼共291尾,其中雌鱼98尾,平均体质量(2 060±350)g,全长(64.18±4.94)cm,体高(19.84±1.83)cm;雄鱼 193尾,平均体质量(500±140)g,全长(42.08±4.77)cm,体高(11.78±1.48)cm。剪取部分鳍条组织用于提取基因组DNA[23],同时采用只读式电子芯片进行个体标识。
经过分析获得上述后备亲鱼的遗传结构,根据个体间的遗传距离划分繁殖谱系,以混合家系的生产方式获得选育组,以公司常规生产的苗种作为对照,进行室内工厂化养殖。经10个月养殖,分别从群体选育组和对照组子代中随机选取300尾个体,测量体质量、体长等生长性状。
1.2 标记的选择
依据半滑舌鳎高密度遗传连锁图谱[21]均匀选择并合成191个微卫星标记,随机选用6个样本筛选标记的扩增条件及多态性,最终用于群体分析的20个微卫星标记分布于12个连锁群,其引物序列见表1。
1.3 PCR扩增及检测
用基因组DNA提取试剂盒(TIANGEN,Code No.DP304)从鳍条组织中提取基因组DNA,紫外分光光度计定量后稀释成50ng/μL。建立15μL PCR反应体系,含有终浓度10 mmol/L Tris-Cl(pH 8.0)、50mmol/L KCl、1.5mmol/L MgCl2、200μmol/L dNTP、0.5μmol/L引物,以及1U Taq DNA聚合酶和100ng DNA模板。反应程序为:94℃预变性3min;94℃变性 30s,58℃复性 30s,72℃延伸 30s,27 个循环;最后72℃延伸5min。扩增产物用8%非变性聚丙烯酰胺凝胶电泳检测,银染显带。以DL1000(Takara Bio Inc.,Code No.3590A)作为分子量标准,用软件Gel-ProAnalyzer(Version 4.5)分析电泳条带的片段长度。
1.4 数据处理
使用软件“鱼类种质资源遗传分析装置(ZL200710144749.3)”将基因型数据转化为“0,1”格式,用软件POPGENE(Version 1.32)计算各标记等位基因频率(allele frequency,P)、等位基因数(observed number of alleles,Na)、有效等位基因数(effective number of alleles,Ne)、观测杂合度(observed heterozygosity,Ho)和期望杂合度(expected heterozygosi-ty,He)。
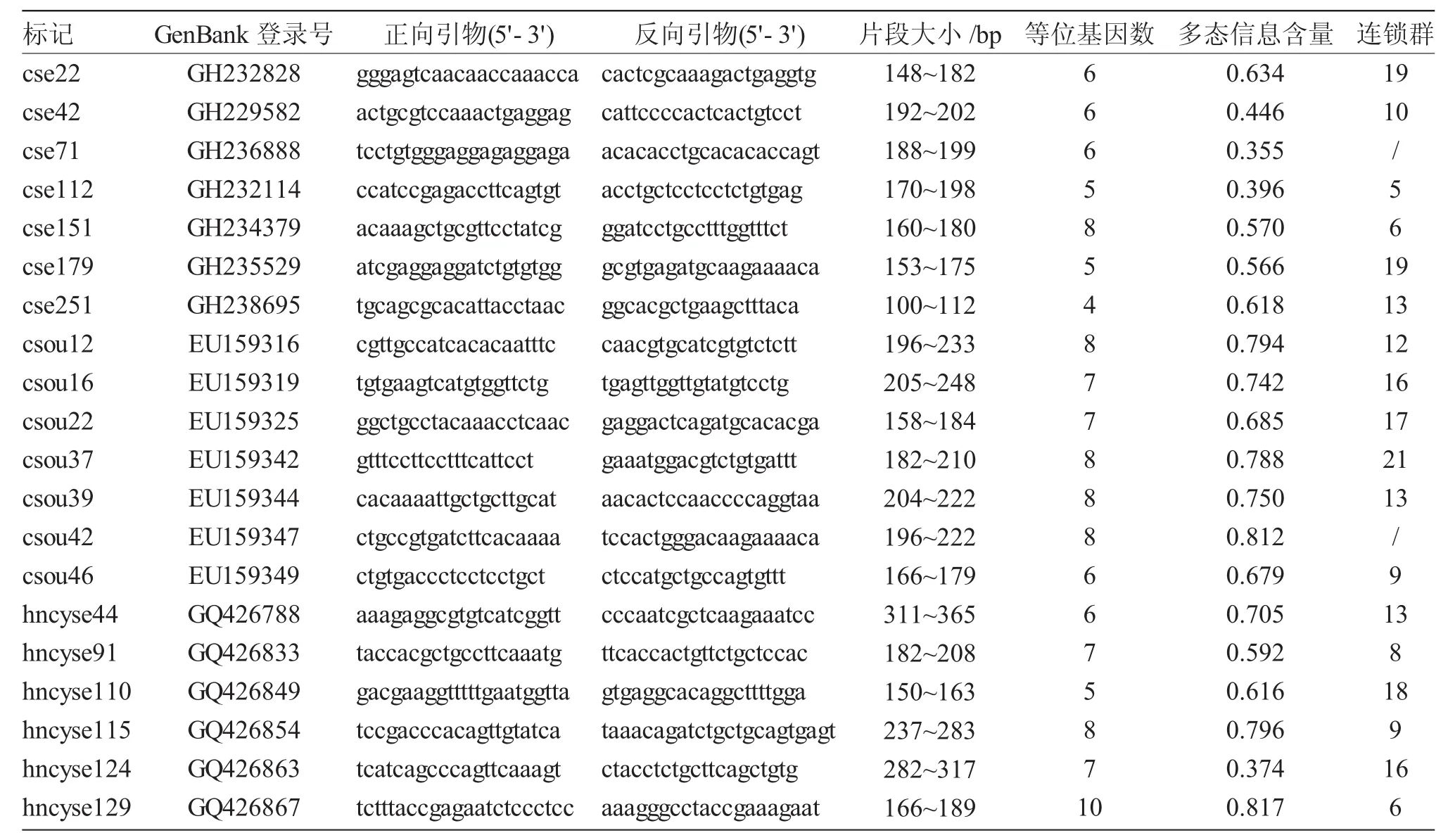
表1 20个半滑舌鳎多态微卫星标记的引物信息Tab.1 Information of twenty microsatellite markers for half-smooth tongue sole C.semilaevis
用Bosteine等[24]的公式计算多态信息含量(polymorphisminformation content,PIC):
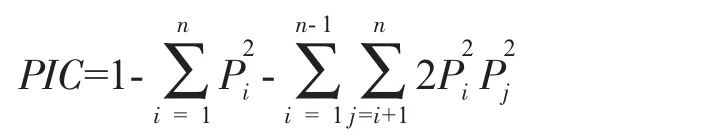
式中,Pi、Pj分别为群体中第i个和第j个等位基因频率,n为某一基因座上等位基因数。
使用软件 PHYLIP(Version 3.695)的 GENDIST程序,计算雌、雄个体间的遗传距离,使用软件“基于遗传背景的分子育种软件1.0”(登记号2011SR09-1052)绘制雌、雄个体聚类图并划分繁殖系谱。
2 结果与分析
2.1 微卫星标记筛选及扩增情况
随机选取6个半滑舌鳎样本确定191个微卫星标记的最适退火温度及多态性。结果共有180个标记获得清晰、稳定的目的条带,112个标记表现出不同程度的多态性,多态微卫星标记的比例为62.20%。从中选择20个微卫星标记(表2)检测全部样本的基因型,片段大小在100~365bp之间,每个标记扩增出4~10个等位基因,共检测到135个等位基因,多态信息含量在0.355~0.817之间,其中16个为高度多态标记(PIC≥0.5)[24],占 80%(图1)。
2.2 雌、雄群体遗传结构
鉴于半滑舌鳎雌、雄亲鱼的生长性状差异巨大,对雌、雄繁育群体的遗传结构分别统计(表2)。结果:20个微卫星标记在雌性群体的有效等位基因数(Ne)为 1.792~5.686,平均为 3.578;雄性群体为1.471~6.116,平均为3.508,雌性群体略大于雄性群体,但是,成对样本T检验结果显示,两者的差异不显著(P=0.713>0.05)。雌性群体的观测杂合度(Ho)为0.418~0.835,平均为 0.606;雄性群体为 0.28~0.798,平均为0.548,雌性群体的观测杂合度极显著大于雄性群体(P=0.003<0.01)。雌性群体的期望杂合度(He)和多态信息含量(PIC)较雄性群体高,但也没有达到显著水平(期望杂合度P=0.077;多态信息含量P=0.081)。统计结果表明:该半滑舌鳎雌性群体的多态性略大于雄性群体,亲鱼繁殖群体保持了高度的遗传多样性水平(PIC>0.5)[24],具备进一步繁殖、筛选优良品系的遗传潜力。
在 20 个标记中,cse22(174bp)、cse42(198bp)、cse71(195bp)、cse112(184bp)、cse179(171bp)和hncyse124(300bp)6个标记的单个等位基因在雌、雄群体的等位基因频率均超过了0.5(详细数据未列出),提示人们在选育过程中要注意对稀有等位基因的保护,减少与性状无关的少量等位基因的富集。
2.3 雌雄个体间遗传距离及繁殖策略
依据个体的基因型数据计算表明,半滑舌鳎雌、雄亲本个体间的遗传距离在0.254~1.959之间,平均值为0.890,不同遗传距离范围的频率呈正态分布,其中个体间遗传距离在0.7~1.0之间的占51.82%(图2),根据子代优良性状与亲本间遗传距离呈“n”形曲线的规律[25],设定亲本间遗传距离阈值在0.7~1.0范围之内最佳。
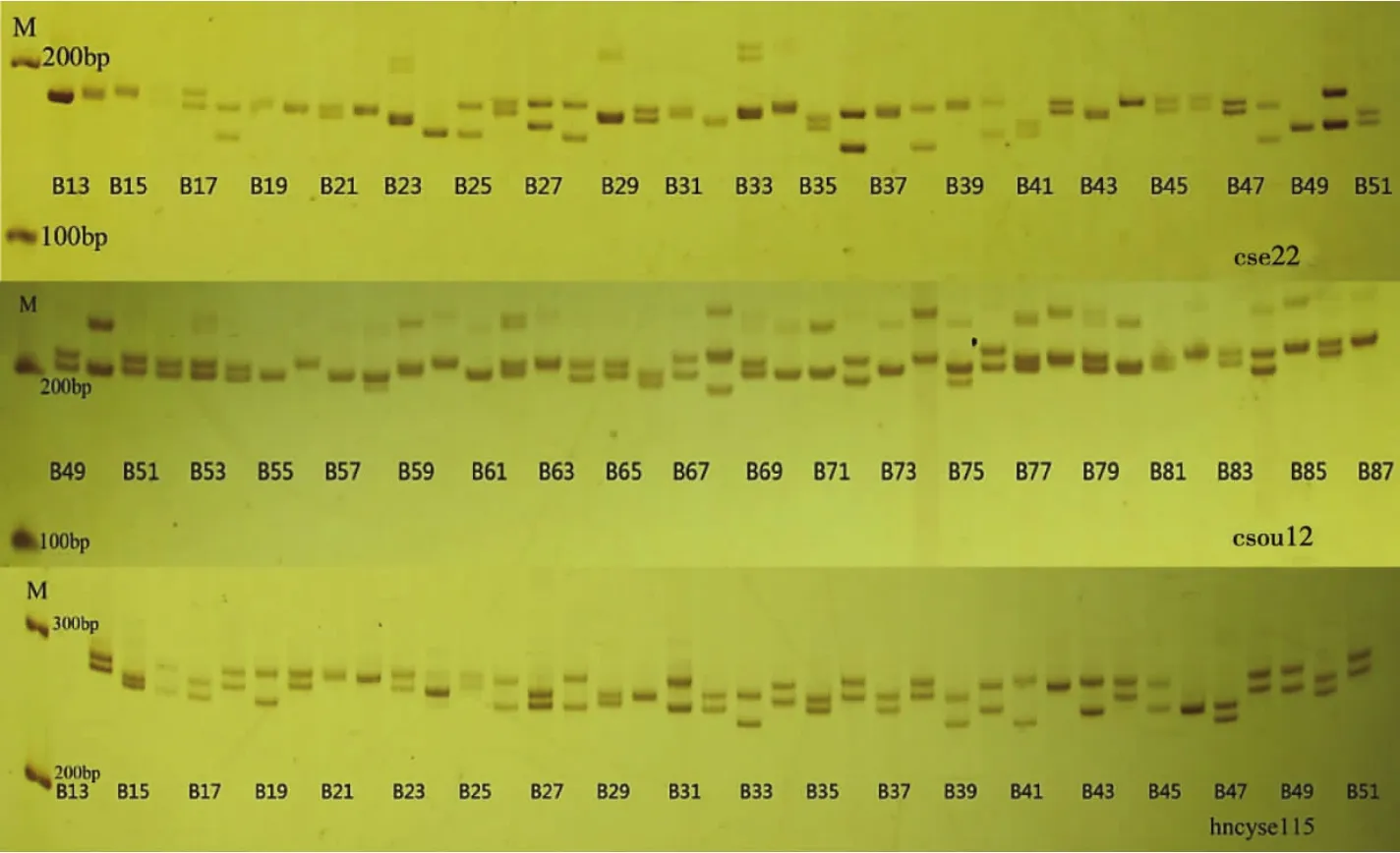
图1 部分半滑舌鳎亲鱼样本在微卫星标记cse22、csou12和hncyse115的电泳图谱Fig.1 Electrophoresis pattern of PCR product of partial samples at cse22,csou12,and hncyse115 in half-smooth tongue sole C.semilaevis
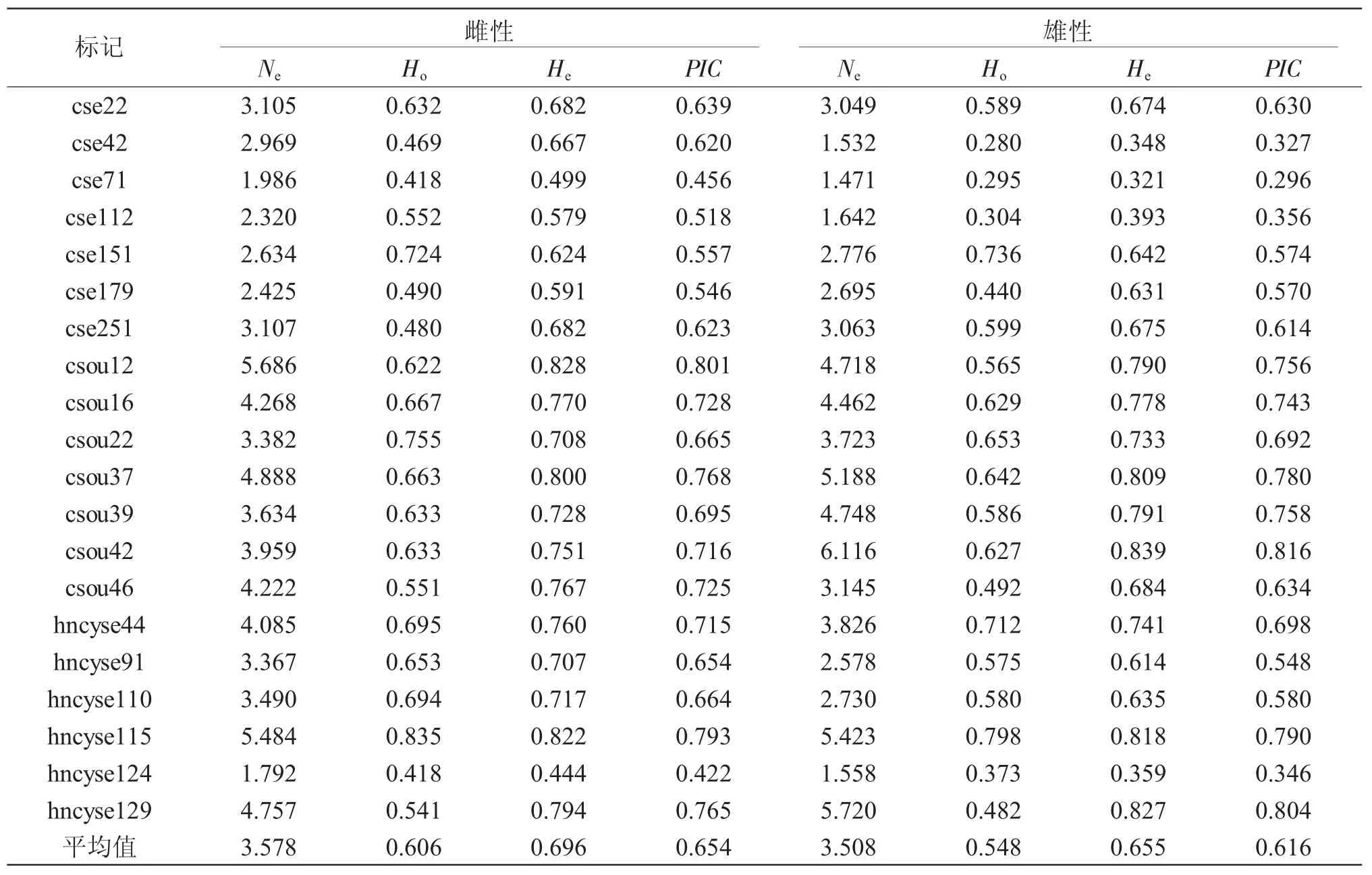
表2 半滑舌鳎群体的遗传多样性参数Tab.2 Genetic diversity index of microsatellite loci in half-smooth tongue sole C.semilaevis population
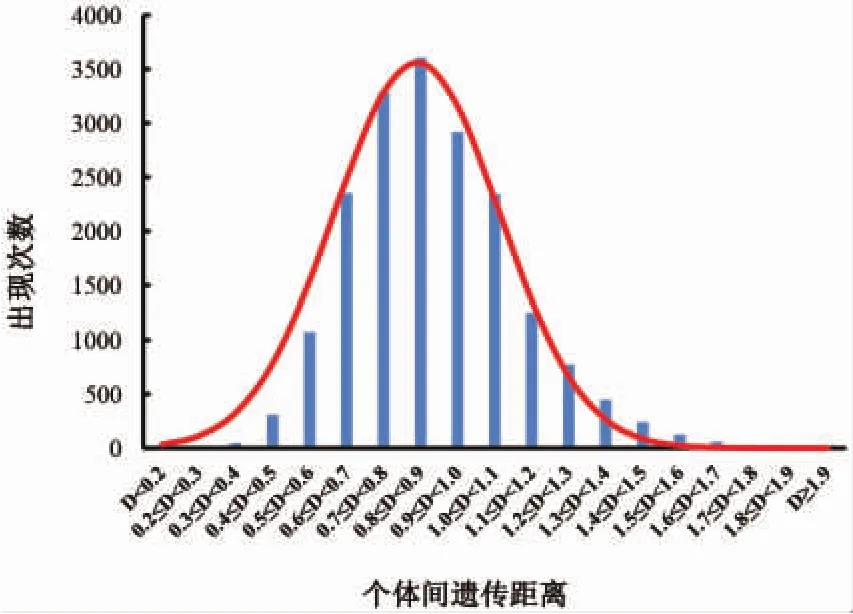
图2 半滑舌鳎雌、雄个体间遗传距离分布Fig.2 Profile of genetic distance between female and male in half-smooth tongue sole C.semilaevis population
用“基于亲本遗传背景的分子育种软件1.0”采用家系配组和群体配组相结合的方式划分繁殖系谱,最终确定半滑舌鳎的繁殖策略为:选择位于不同繁殖系谱、遗传距离在0.7~1.0之间,且表型性状在均值以上的一对或多对亲本生产单家系或混合家系。
根据此策略获得的混合家系选育组,对体质量、体长等生长性状的统计结果显示,在相同养殖条件下,经本策略获得的混合家系选育组半滑舌鳎的平均体质量、体长与对照组差异显著,其中平均体质量高于对照组24.56%,平均体长高于对照组6.98%(表3)。
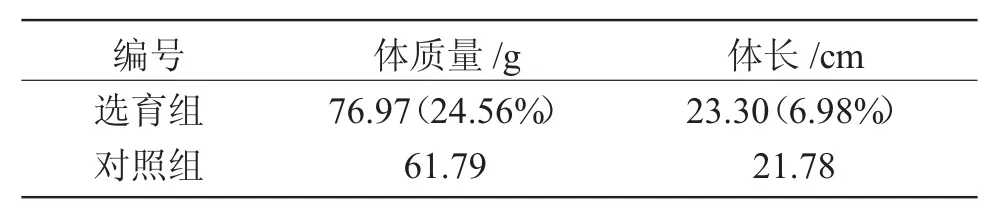
表3 半滑舌鳎亲本经选育的子代和对照组子代的生长性状Tab.3 Growth of F1in selective population and control population in half-smooth tongue sole C.semilaevis
3 讨论
自21世纪初半滑舌鳎的人工繁殖及工厂化育苗技术逐渐成功以来,由于价值高、利润大,苗种繁育和成鱼养殖产业发展迅猛。维持繁殖群体内的遗传多样性水平是群体持续利用的前提和保障。本研究采集的半滑舌鳎繁殖群体样本较大,雌性的有效等位基因数、观测杂合度、期望杂合度和多态信息含量4项遗传参数分别为 3.578、0.606、0.696和0.654;雄性分别为 3.508、0.548、0.655 和 0.616,表明半滑舌鳎育种群体处于高度的多态水平,具备进一步开展品种选育和生产的潜力。但是,遗传参数较 2011 年徐营等[26]的研究结果(3.935、0.7135、0.7303和0.6804)已有所降低。多项研究也表明,养殖群体的遗传多样性较野生群体有显著降低[19,27]。因此,在野生资源日益萎缩,无法及时补充现有养殖群体的遗传多度的情况下,在育苗过程中保护现有遗传资源非常必要。
在获得经济利益的同时,多代近交带来的性状衰退也非常严重。以半滑舌鳎白化病为例,在母本均为养殖群体的情况下,父本为养殖群体的家系的平均白化率(19.38%)远高于父本为野生群体的(3.21%)[28]。基于亲本遗传背景的分子育种技术能够最大限度地避免近亲交配[15],其中最核心的技术要点就是要首先确定最佳雌、雄亲本间配组的遗传距离“阈值”。本研究利用20个微卫星标记分析了半滑舌鳎雌、雄个体间的遗传距离,发现其分布呈正态性,与家系生长性状和亲本间遗传距离的相关性趋势相似。鲁翠云等[16]的研究发现,随着镜鲤亲本间遗传距离的增加,家系的生长性状呈先升高后下降的趋势,配组的最佳遗传距离范围在0.5~0.7之间。毕金贞等[29]对牙鲆Paralichthys olivaceus的研究中得到了相似的结果,随着遗传距离的增大,遗传距离与性状的关系呈现显著正相关(D=0.2578~0.5958)、无显著相关性(D=0.6099~0.6604)到显著负相关(D=0.6640~0.9773)的变化。因此,结合生产实际和本研究结果,确定指导半滑舌鳎家系配组的亲本间遗传距离范围为0.7~1.0之间。
半滑舌鳎作为近十几年新兴的海水养殖鱼类,亲鱼多为经过人工驯化的野生种或选育时间短的累代养殖种,在选育过程中制定正确的选育方式,避免近亲繁殖,保护群体遗传多样性对种质资源的持续利用及养殖业的健康发展十分必要。本研究借鉴其他鱼类的分子育种经验,结合研究结果和生产实际,制定出半滑舌鳎分子育种的策略为:选择位于不同繁殖系谱,遗传距离在0.7~1.0之间,且表型性状在均值以上的一对或多对亲本生产单家系、混合家系或群体繁殖,将群体内或家系内的近交系数降至最低,以最大限度地避免由近亲繁殖造成的优良性状衰退,并在此基础上生产、筛选性状优良的基础群体。
[1]李静,陈松林,温海深.鱼类性别相关基因及性别特异标记的研究进展[J].海洋水产研究,2006,27(4):90-95.
[2]Chen S L,Li J,Deng S P,et al.Isolation of female-specific AFLP markers and molecular identification ofgenetic sexin half-smooth tongue sole (Cynoglossus semilaevis)[J].Marine Biotechnology,2007,9(2):273-280.
[3]马洪雨,陈松林,李静,等.半滑舌鳎雌性特异AFLP标记CseF783的克隆及其在遗传性别鉴定中的应用[J].遗传,2009,31(1):88-94.
[4]Liao X L,Xu G B and Chen S L.Molecular method for sex identification of half-smooth tongue sole(Cynoglossus semilaevis)using a novel sex-linked microsatellite marker[J].International Journal of Molecular Sciences,2014,15(7):12952-12958.
[5]Chen S L,Tian Y S,Yang J F,et al.Artificial gynogenesis andsexdeterminationinhalf-smoothtonguesole(Cynoglossus semilaevis)[J].Marine Biotechnology,2009,11(2):243-251.
[6] Ji X S,Tian Y S,Yang J F,et al.Artificial gynogenesis in Cynoglossus semilaevis with homologous sperm and its verificationusingmicrosattlitemarkers[J].AquacultureResearch,2010,41(6):913-920.
[7]陈松林,李文龙,季相山,等.半滑舌鳎三倍体鱼苗的人工诱导与鉴定[J].水产学报,2011,35(6):925-931.
[8]李文龙,陈松林,季相山,等.半滑舌鳎四倍体鱼苗的诱导与鉴定[J].中国水产科学,2012,19(2):196-201.
[9]刘志鹏,王旭波,翟介明,等.半滑舌鳎三倍体诱导研究[J].中国海洋大学学报,2012,42(6):77-80.
[10]周丽青,杨爱国,柳学周,等.半滑舌鳎染色体核型分析[J].水产学报,2005,29(3):417-419.
[11]Chen SL,ZhangGJ,ShaoC W,et al.Whole-genome sequence ofa flatfish provides insights into ZW sex chromosome evolution and adaptation to a benthic lifestyle[J].Nature Genetics,2014,46(3):253-260.
[12]张全启,王旭波,王志刚,等.牙鲆和半滑舌鳎性别控制育种研究进展[C].“全球变化下的海洋与湖沼生态安全”学术交流会论文集,2014.
[13]陈松林,杜民,杨景峰,等.半滑舌鳎家系建立及其生长和抗病性能测定[J].水产学报,2010,34(12):1789-1794.
[14]陈松林,李仰真,张静,等.半滑舌鳎快速生长及高雌性家系的筛选[J].水产学报,2013,37(4):481-488.
[15]孙效文,鲁翠云,贾智英,等.水产动物分子育种研究进展[J].中国水产科学,2009,16(6):981-990.
[16]鲁翠云,曹顶臣,孙效文,等.微卫星分子标记辅助镜鲤家系构建[J].中国水产科学,2008,15(6):893-901.
[17]鲁翠云,金万昆,李超,等.微卫星分子标记指导镜鲤群体选育[J].水产学杂志,2011,24(3):29-34.
[18]常玉梅.鲤鱼、中华绒螯蟹和大黄鱼微卫星标记的开发与应用研究[D].广州:中山大学,2008.
[19]黎中宝,雷光高,常建波,等.半滑舌鳎(Cynoglossus semilaevis)野生与养殖群体的遗传多样性的比较研究[J].海洋与湖沼,2011,42(3):414-418.
[20]Wang X,Zhang Q,Sun X,et al.Isolation and characterization of 64 novel microsatellite markers from a fosmid library of female half-smooth tongue sole(Cynoglossus semilaevis)[J].Molecular Ecology Resource,2008,8(6):1303-1306.
[21]Song W,Li Y,Zhao Y,et al.Construction of a high-density microsatellite genetic linkage map and mapping of sexual and growth-related traits in half-smooth tongue sole (Cynoglossus semilaevis)[J].PloS One,2012,7(12):e52097.
[22]张俊杰,张文婷,高峰涛,等.半滑舌鳎3D-BAC池构建及性别连锁标记的物理定位[J].农业生物技术学报,2015,23(2):141-150.
[23]李超,鲁翠云,郑先虎,等.一种保存鱼类鳍条的便捷方法[J].水产学杂志,2014,27(1):22-24.
[24]Botstein D,White R L,Skolnick M,et al.Construction ofa genetic linkage map in man using restriction fragment length polymorphisms [J].American Journal of Human Genetics,1980,32(3):314-331.
[25]孙效文.鱼类分子育种学[M].北京:海洋出版社,2010.
[26]徐营,邵长伟,邓寒,等.半滑舌鳎养殖群体和减数分裂雌核发育群体的微卫星标记遗传多样性分析[J].渔业科学进展,2011,32(4):14-19.
[27]马骞,林琳,柳淑芳,等.半滑舌鳎生长相关基因的微卫星及其在种群遗传结构分析中的应用[J].渔业科学进展,2012,33(4):18-25.
[28]李仰真,陈松林,邢贺飞,等.半滑舌鳎白化现象的初步研究[J].水产学报,2014,38(1):76-83.
[29]毕金贞,陈松林.牙鲆亲本间遗传距离与其后代生长速度的相关性分析[J].中国农学通报,2010,26(15):395-401.
猜你喜欢
中华实用诊断与治疗杂志(2022年1期)2022-08-31
中华实用诊断与治疗杂志(2022年1期)2022-08-31
中国临床医学影像杂志(2022年5期)2022-07-26
烟台大学学报(自然科学与工程版)(2022年3期)2022-06-30
西南农业学报(2022年5期)2022-06-06
水产科学(2022年2期)2022-03-20
南京师范大学学报(工程技术版)(2021年2期)2021-10-20
中国糖料(2021年3期)2021-07-13
中南林业科技大学学报(2019年4期)2019-04-08
热带农业科技(2019年1期)2019-01-14
