
一株H3N8亚型马流感病毒的鉴定及HA基因序列分析
2018-05-21 04:22王小蕾段会娟杨志远刘立新赵际成刘月焕
动物医学进展 2018年4期
王小蕾,段会娟,杨志远,刘立新,赵际成,潘 洁,刘月焕,林 健
(北京市农林科学院畜牧兽医研究所,北京 100097)
马流感是由正黏病毒科A型流感病毒属病毒引起的马、驴、骡等动物的一种急性呼吸道传染病。世界动物卫生组织(OIE)规定马流感为法定报告动物疫病,我国将其列为三类动物疫病。马流感在世界多个国家均有流行,该病的暴发给赛马业带来巨大的经济损失[1-2]。马流感病毒(Subtype equine influenza virus,EIV)分为H7N7(马A1型)和H3N8(马A2型)2个亚型。H7N7 EIV于1956年在捷克布拉格马群中分离到,但自1977年以后再未从马体内分离到;H3N8亚型EIV于1963年在美国迈阿密从阿根廷进口的马群中首次分离到,此后在多个国家暴发和流行[3]。H3N8亚型EIV根据分离地分成欧洲谱系和美洲谱系,目前这2种类型的EIV在养马的国家都存在[4]。最近的遗传进化分析表明,美洲谱系H3N8亚型EIV进一步分成3个分支,分别为南美分支、肯塔基分支和佛罗里达分支[5],其中佛罗里达分支又分为Ⅰ群和Ⅱ群[3]。
新中国成立以来,我国大陆经历了4次较大的马流感疫情。第1次流行发生在1974年,导致此次马流感疫情的毒株是H7N7亚型的EIV,代表毒株为A/马/京防/74-1。第2次疫情发生在1989年-1990年,导致疫情的是禽源的H3N8亚型EIV,代表毒株是A/euqine/Jilin/1/1989。第3次的马流感疫情发生在1992年-1994年,也是由H3N8亚型EIV引起,代表毒株为A/equine/Qinghai/1/1994[6]。后来的回顾性调查研究表明,导致我国20世纪90年代流行的EIV为欧洲谱系[7]。2007年-2008年我国再次暴发马流感大流行,由新疆蔓延至全国各地,沉重打击了我国的养马业和即将兴起的赛马业,分离到的流行株A/equine/Xinjiang/3/2007 (H3N8)[8]、A/Equine/Huabei/1/2007(H3N8)[9]通过遗传进化分析显示为美洲谱系。最近一次报道的马流感疫情于2013年8月发生在江苏徐州,分离的毒株属于美洲谱系[7]。
2016年4月北京市农林科学院畜牧兽医研究所动物疫病研究室从北京密云发病马群中分离到1株马流感病毒,命名为A/equine/Beijing/2016,经鉴定为H3N8亚型。对该病毒分离株及近10年来国内部分H3N8亚型马流感分离株进行了HA基因进化及抗原位点的初步分析,为我国马流感防控选择有效的疫苗候选株提供参考。
1 材料与方法
1.1 材料
1.1.1 鸡胚 10日龄SPF鸡胚,购于北京梅里亚维通实验动物技术有限公司。
1.1.2 试剂和引物 TPB培养基、PBS(0.01 mol/L,pH 7.2)、双抗、阿氏液和0.5%鸡红细胞悬液按常规制备。RNA提取试剂盒、RNA PCR Kit (AMV) Ver.3.0,宝生物工程(大连)有限公司产品。参考GenBank数据库中H3N8亚型EIV的基因序列,用Primer5.0引物设计软件设计HA基因和NA基因的特异性引物。引物序列见表1。
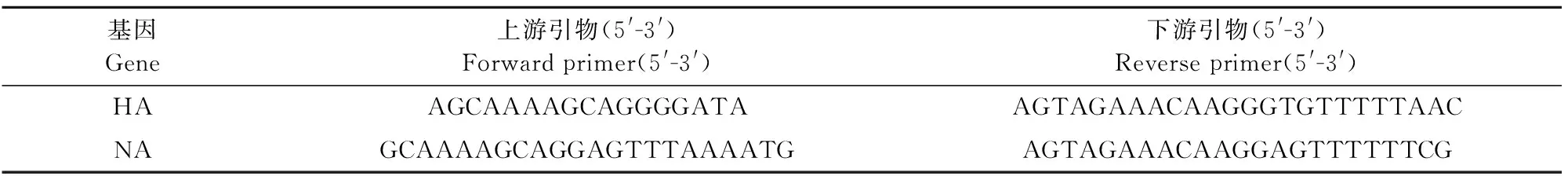
表1 HA和NA基因的特异性引物
1.1.3 EIV阴性、阳性参考血清及EIV H3亚型HI抗原 EIV H7N7 亚型(A/ equine/Prague/1/56)阳性血清购于英国 VLA ( Veterinary Laboratory Agency );EIV H3亚型HI抗原(HB株)及阴性、阳性血清,购于中海生物科技有限公司。
1.2 方法
1.2.1 样本采集 选择临床症状典型的病马,用无菌棉拭子蘸取鼻腔分泌物,置于装有TPB溶液的离心管;采集病马的血液,分离血清。拭子及血清样品置-20℃保存。
1.2.2 血清学检测 分别采取3匹发病马发病初期和发病后2周血清,用分离毒株、马流感H3亚型HI抗原通过血凝抑制试验检测发病前后血清抗体。
1.2.3 病毒的分离和传代 装有分泌物拭子的离心管振荡后5 000 r/min离心10 min,取上清用0.22 μm滤膜过滤。以0.2 mL/胚的剂量经尿囊腔接种SPF鸡胚,置36℃孵化至96 h,收获24 h~96 h存活鸡胚的尿囊液。将收获的病毒尿囊液稀释后,经SPF鸡胚传至第3代,分别测定每代鸡胚尿囊液对0.5%鸡红细胞悬液的凝集(HA)效价。
1.2.4 血凝试验(HA) 将收集的F1~F3代鸡胚尿囊液作为被检抗原,与0.5%鸡红细胞进行凝集试验。
1.2.5 血凝抑制试验(HI) 分别将F3代病毒液及EIV H3亚型HI抗原(HB株)用PBS稀释成含6个血凝单位后作为被检抗原,分别与H3N8、H7N7亚型EIV阳性参考血清及采集的发病马血清进行HI试验。
1.2.6 HA基因和NA基因测序 病毒RNA提取和RT-PCR分别按试剂盒说明书进行,HA和NA基因的扩增产物送宝生物工程(大连)有限公司进行载体构建并测序。
1.2.6.1 序列分析 通过NCBI BLAST将病毒分离株与GenBank数据库中的EIV进行核苷酸序列同源性比较。将测得的HA基因核苷酸序列用DNA Star软件推导出氨基酸序列,对该分离株和已报道的近年来部分国内EIV分离株及目前OIE推荐疫苗株的HA氨基酸序列进行比较分析。
1.2.6.2 绘制系统发育树 应用MEG6.0软件,分别绘制HA基因的进化树,分析病毒分离株的遗传演化关系。
2 结果
2.1 临床调查
发病马场初期共有6匹赛马发病,病初体温稍微升高,一般在39℃左右,个别超过40℃,稽留3 d~5 d,结膜潮红,流泪;病初干咳严重,后期转为湿咳,有痰;病初流浆液性鼻液,后逐渐变成灰白色脓性分泌物,下颌淋巴结正常,触诊喉头无疼痛反应。病马厌食,精神委顿,呼吸频数。大多数马病程持续1周左右,然后逐渐康复。之后周边地区马陆续出现类似症状,没有死亡病例。
2.2 病毒分离
接种鸡胚96 h内全部存活,存活鸡胚尿囊液对0.5%鸡红细胞的凝集效价为1∶16~1∶64之间。F1、F2代血凝效价较低,F3代血凝效价达到1∶64。
2.3 血凝抑制试验
分离毒株与马流感H7亚型阳性和阴性血清、发病初期的马血清HI试验结果均为阴性,与发病后2周的血清及马流感H3亚型阳性血清为阳性反应。此结果与马流感H3亚型HI抗原(HB株)检测结果一致,说明该病毒分离株为H3亚型。将该分离株命名为A/equine/Beijing/2016。具体检测结果见表2。
2.4 HA、NA基因测序结果
用HA基因和NA基因的特异性引物进行PCR,各得到一条特异性条带(图1)。
经测序,A/equine/Beijing/2016的HA基因达1 768 bp,NCBI BLAST结果显示,A/equine/Beijing/2016与A/equine/Xuzhou/01/2013 (H3N8) (GenBank:KF806985.1)同源性最高,达99%。NA基因为1 459 bp,与A/equine/Heilongjiang/SS1/2013 (H3N8)(GenBank:KC986392.2)同源性最高,达99%。A/equine/Beijing/2016为H3N8亚型。
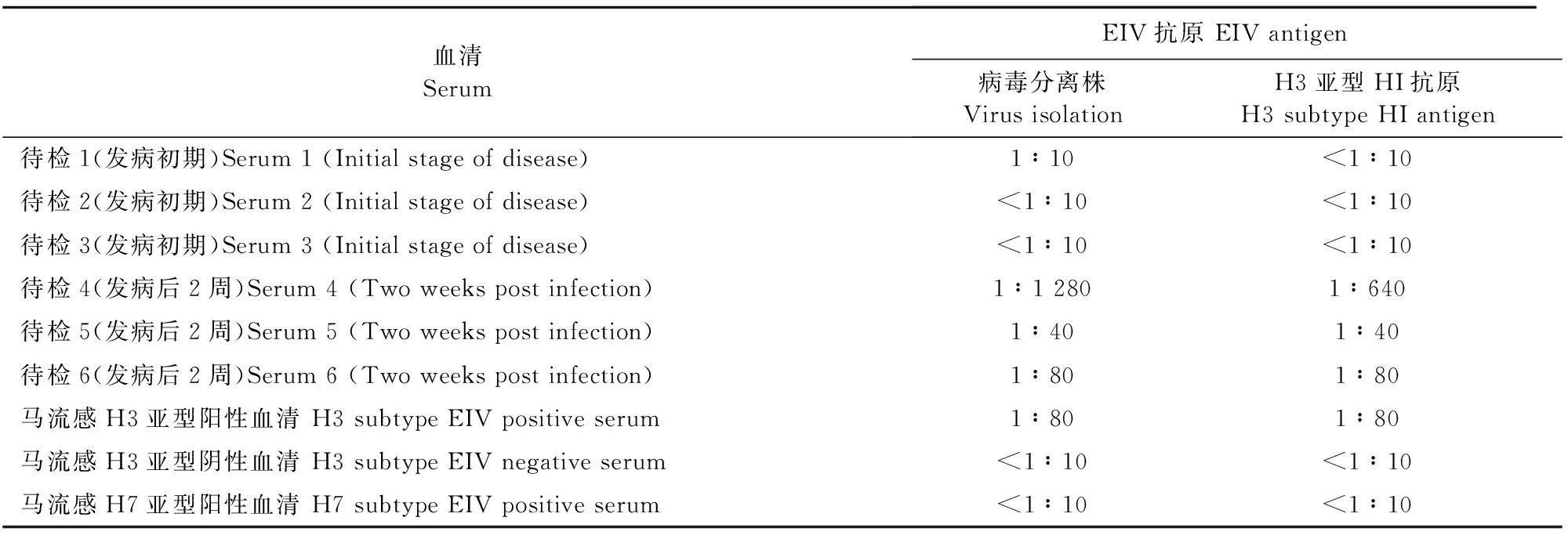
表2 马血清EIV HI效价

1.HA基因;2.NA基因;M.DNA标准DL 2 0001.HA gene; 2.NA gene; M.DNA Marker DL 2 000
2.5 HA基因的遗传演化分析
如图2所示,HA基因的遗传进化分析表明,A/equine/Beijing/2016 为H3N8亚型EIV美洲谱系的佛罗里达Ⅱ群,与国内2007年以来的EIV分离株一致。
2.6 HA氨基酸序列分析
流感病毒的HA基因易发生变异,导致抗原性发生改变。将A/equine/Beijing/2016及国内其他研究者报道的一部分美洲谱系佛罗里达Ⅱ群H3N8亚型EIV分离株推导的HA 氨基酸序列与OIE推荐的佛罗里达Ⅱ群疫苗候选株A/equine/Richmond/1/2007的HA 氨基酸序列进行比较,发生改变的氨基酸位点如表3所示。根据H3亚型人流感病毒HA蛋白的抗原位点及序列特征[10],人们推测了H3N8亚型马流感病毒表面的许多重要的氨基酸位点,命名为A、B、C、D和E。由表3可见,相比于OIE推荐的疫苗候选株A/equine/Richmond/1/2007,分离株A/equine/Beijing/2016在抗原位点A(144 Ala→Thr)、抗原位点B(159Asn→Asp,198Glu→Gly)、抗原位点C(276Ile→Val)均发生了改变。
3 讨论
近年来,国内外不断有马流感发生的报道,所分离的毒株多为H3N8亚型美洲谱系的佛罗里达分支[11-15]。此次北京某马场出现马流感疑似疫情后,通过临床调查和实验室病毒分离、血清学检测、分离毒株HA及NA基因序列测定和分析,证实此次分离毒株为H3N8亚型马流感病毒,同样属于美洲谱系的佛罗里达Ⅱ群。2007年-2016年,北京市农林科学院畜牧兽医研究所动物疫病研究室在内蒙、甘肃、河北、新疆等地区进行了马流感疫情调查,并采集马血清进行抗体检测。在调查中发现,近年来马流感在上述地区多次发生,其发生频率远高于2007年之前,应该引起行业的重视。
根据文献报道[16],5个公认的抗原位点中如果有≥2个抗原位点同时出现≥4个氨基酸的变化,则意味着一个重要的抗原漂移。对此次EIV分离株的HA氨基酸序列的分析发现,相比OIE推荐的佛罗里达Ⅱ群的疫苗候选株A/equine/Richmond/1/2007,该分离株出现了一个重要的抗原漂移,但其致病性、免疫原性等特性的研究有待进行。2007年-2013年间的EIV分离株相互比对,也出现了一些位点的氨基酸序列变化,这提示HA的氨基酸序列可能处于不断变异之中。目前国内马流感尚无商品化疫苗可用于免疫预防,但对马流感疫苗的研发已有多项报道[17]。流感病毒HA基因的抗原漂移可能会降低疫苗的保护效力,致使在疫苗免疫的马匹中发生马流感,因此在疫苗研发时根据流行情况选择新分离病毒株作为疫苗候选株显得尤为重要。而这也为当前马流感的防疫工作带来挑战,有必要加强对国内马流感病毒的流行病学监测。

■为本此分离得到的分离株;▲为我国大陆2007年以来的部分EIV分离株;◆为我国大陆20世纪90年代部分EIV分离株■ The isolate of this test; ▲Part of EIV isolates in mainland China since 2007; ◆Part of EIV isolates in mainland China in 1990’s

马流感病毒株Equineinfluenzavirus氨基酸位置Aminoacidposition71147135A142144155B156158159165168198B228234240276C290291329Richmond/1/2007NASRGATKGNNMEGWGINERHuabei/1/2007··················D·Xinjiang/3/2007··TI·······T······D·Xinjiang/5/2007··TI··············D·Guangxi/1/2008····E··A···········D·Hubei/6/2008··TI··············D·Gansu/7/2008··················D·InnerMongolia/8/2008··TI··············D·Liaoning/9/2008··T·······D·······DKHeilongjiang/10/2008············G·····D·Heilongjiang/1/2010·······NE···G·····D·Heilongjiang/1/2011·····T······G·····D·Heilongjiang/SS1/2013·····T······GR····D·Xuzhou/01/2013·····T······G·C·V·D·Beijing/2016·S···T···D··G···VTD·
参考文献:
[1] Guthrie A J,Stevens K B,Bosman P P.The circumstances surrounding the outbreak and spread of equine influenza in South Africa [J].Rev Sci Tech,1999,18(1):179-185.
[2] Powell D G,Watkins K L,Li P H,et al.Outbreak of equine influenza among horses in Hong Kong during 1992 [J].Vet Rec,1995,136(21):531-536.
[3] Bryant N A,Rash A S,Russell C A,et al.Antigenic and genetic variations in European and North American equine influenza virus strains (H3N8) isolated from 2006 to 2007 [J].Vet Microbiol,2009,138(1-2):41-52.
[4] Daly J M,Lai A C,Binns M M,et al.Antigenic and genetic evolution of equine H3N8 influenza a viruses [J].J Gen Virol,1996,77 ( Pt 4):661-671.
[5] Lai A C,Chambers T M,Holland Re Jr,et al.Diverged evolution of recent equine-2 influenza (H3N8) viruses in the Western Hemisphere [J].Arch Virol,2001,146(6):1063-1074.
[6] 杨建德,相文华.我国马流感的研究现状 [J].黑龙江畜牧兽医,2002(3):42-44.
[7] 葛菲菲,刘 健,鞠厚斌,等.1株H3N8亚型马流感病毒的分离鉴定及全基因进化分析 [J].中国兽医学报,2015,35(9):1435-1440.
[8] 戴伶俐.马流感病毒A/Equine/Xinjiang/3/07(H3N8)HA基因的序列分析及两种PCR检测方法的建立[D].黑龙江哈尔滨:中国农业科学院,2009.
[9] 蒋桃珍,刘月焕,林 健,等.2007年华北地区H3N8亚型马流感病毒的分离与鉴定 [J].中国兽医学报,2010,30(5):607-611.
[10] Wiley D C,Wilson I A,Skehel J J.Structural identification of the antibody-binding sites of Hong Kong influenza haemagglutinin and their involvement in antigenic variation [J].Nature,1981,289(5796):373-378.
[11] Alves Beuttemmuller E,Woodward A,Rash A,et al.Characterisation of the epidemic strain of H3N8 equine influenza virus responsible for outbreaks in South America in 2012 [J].Virol J,2016,13:45. doi: 10.1186/s12985-016-0503-9.
[12] Rash A,Morton R,Woodward A,et al.Evolution and divergence of H3N8 equine influenza viruses circulating in the United Kingdom from 2013 to 2015 [J].Pathogens,2017,6(1):1-16.
[13] Fougerolle S,Legrand L,Lecouturier F,et al.Genetic evolution of equine influenza virus strains (H3N8) isolated in France from 1967 to 2015 and the implications of several potential pathogenic factors [J].Virology,2017,505:210-217.
[14] 赵 钊,郭 巍,关平原,等.黑龙江省一株马流感病毒的全基因组测序及同源性分析 [J].中国动物检疫,2015,32(12):69-72.
[15] 冯淑萍,韦建华,易春华,等.广西马流感H3N8毒株的HA基因序列测定与分析 [J].黑龙江畜牧兽医,2015(13):31-34.
[16] Wilson I A,Cox N J.Structural basis of immune recognition of influenza virus hemagglutinin [J].Annu Rev Immunol,1990,8:737-771.
[17] 刘春国,王 伟,刘 飞,等.H3N8亚型马流感病毒VLPs的构建及其生物学特性分析 [J].中国兽医科学,2014,44(3):240-244.
猜你喜欢
艺术品鉴(2022年16期)2022-07-09
科学大观园(2022年2期)2022-01-23
河池学院学报(2021年1期)2021-07-10
景德镇陶瓷(2021年1期)2021-03-24
广东医科大学学报(2020年6期)2020-02-06
东方考古(2017年0期)2017-07-11
现代检验医学杂志(2016年3期)2016-11-15
现代检验医学杂志(2016年1期)2016-11-12
分析测试学报(2015年6期)2016-01-13
动物医学进展(2015年10期)2015-12-07
