
IL-2对布鲁菌侵染小鼠巨噬细胞释放细胞因子及吞噬能力的调节作用
2018-05-21 04:22齐江姣李村院王元元周光普高剑峰
动物医学进展 2018年4期
齐江姣,罗 成,李村院,王元元,周光普,谭 君,高剑峰
(石河子大学生命科学学院细胞分子生物学实验室,新疆石河子 832003)
白介素2(IL-2)是一种调控免疫应答的重要因子[1],它具有多向性作用(主要是对淋巴细胞生长、增殖、分化的促进),对机体的免疫应答和抗病菌、病毒等有重要作用,能活化T 细胞,促进细胞因子产生,刺激自然杀伤细胞(NK细胞)增殖,提高NK细胞杀伤活性及产生细胞因子[2],淋巴因子激活的杀伤细胞(LAK细胞)的诱导产生;促进B细胞增殖和分泌抗体;具有广泛的生物活性(激活巨噬细胞,参与抗体反应、造血和肿瘤监视)。单核-巨噬细胞受到IL-2 的持续作用后,其抗原递呈能力、杀菌力、细胞毒性均明显增强,分泌某些细胞因子的能力也得到加强[3]。
研究发现,IL-2对于一些疾病的治疗或癌症细胞的作用已取得了一系列的成果[4-6]。Roviello G等[7]研究结果表明,IL-2对黑色素瘤或肾以外的实体肿瘤有重要作用。Abhishek K等[8]详细阐述了IL-2与登革热严重程度的关系,结果表明IL-2对登革热治疗有重要的作用。熊一全等[9]就IL-2增强淋巴细胞对乳腺癌细胞的杀伤作用研究,表明IL-2能促进淋巴细胞对MCF-7的杀伤作用。但对于IL-2在布鲁菌病(简称布病)炎症反应中的作用尚不清楚,由于布病是一种世界范围内流行的高度接触性人畜共患病,会导致不孕不育及流产等症状,给人类的健康和畜牧业的发展造成巨大的威胁[10],故研究IL-2对被M5株侵染的巨噬细胞(MΦ)所释放的细胞因子(TNF-α、IFN-γ和NO)及吞噬能力的调节作用具有重要的意义[11]。本文就巨噬细胞本身TNF-α、NO和IFN-γ的含量[12]以及在IL-2对M5侵染的腹腔巨噬细胞对上述细胞因子释放量进行比较,以此探讨IL-2对布病反应的调节作用,为布病的治疗提供新的理论依据。
1 材料与方法
1.1 材料
1.1.1 实验动物与菌种 昆明小鼠,18 g~22 g,由石河子大学实验动物中心所提供;布鲁菌的菌种(M5),由石河子大学生命科学学院细胞分子生物学实验室保存。
1.1.2 主要试剂 IL-2 ,Cyagen 生物公司产品;TNF-α、NO、IFN-γ酶联免疫吸附试验(ELISA) 试剂盒,美国Biosource公司产品;DMEM 培养基,美国Gibco 公司产品;胎牛血清,Hyclone 公司产品;台盼蓝染色液,北京中生瑞泰科技有限公司产品。
1.1.3 仪器设备 高压蒸汽自动灭菌锅,Hirayama公司产品;CO2培养箱、-80℃超低温冰箱,Thermo公司产品;恒温水浴槽,北京长安科学仪器厂生产;高速低温离心机,Beckman公司产品;611UF纯水仪,Bioteck公司产品。
1.2 方法
1.2.1 腹腔巨噬细胞的收集和纯化 小鼠腹腔注射20 g/L无菌淀粉溶液,每只1.5 mL,24 h后颈椎脱位法处死小鼠,将小鼠浸泡在750 mL/L酒精中5 min进行消毒后,取出小鼠,用无菌的滤纸吸干酒精,用5 mL注射器往腹腔注射5 mL无菌PBS(37℃提前温浴),轻揉腹壁,无菌操作取腹腔液,1 500 r/min离心5 min,洗涤2次后,用不含抗生素的完全DMEM Basic(1×) 培养液(含100 mL/L BSA)悬浮细胞,并置于培养瓶中培养过夜。第2天,用PBS彻底冲洗去除未贴壁细胞,台盼蓝染色检测细胞存活率,用不含抗生素的完全DMEM Basic(1×) 培养液调整细胞浓度为2×105cells/mL,备用。
1.2.2 布鲁菌刺激后细胞因子释放的时效关系 试验组分为阴性对照组(正常细胞)和侵染组(M5侵染),用麦氏比浊管法调整菌液浓度,按100∶1的比例用M5侵染MΦ,将其置于37℃、体积分数为5% CO2的细胞培养箱中培养,分别在培养0、2、6、12、24 h时,收集各时间点的细胞培养上清液10 μL,然后用ELISA试剂盒测定TNF-α、NO 和IFN-γ的浓度大小。
1.2.3 IL-2作用下巨噬细胞吞噬M5的能力 试验组分为阴性对照组(正常细胞)、空白对照组(M5 侵染)、不同浓度IL-2调节组(IL-2+M5侵染),用麦氏比浊管法调整菌液浓度,按100∶1的比例在6孔板中用M5 侵染MΦ,分别加入0、1×106、3×106、5×106UI的IL-2,将其置于37℃、体积分数为5% CO2的细胞培养箱中培养,分别培养0、2、6、12、24 h。取各试验组的MΦ,计数,调整细胞浓度为2×106cells/mL,加入1 mL的裂解液,吸取10 μL,稀释103倍,利用CFU涂布计数法,取1 mL(2 000个细胞)均匀涂布于TSA固体培养基上培养,72 h后观察集落形成情况,计算吞噬指数;
吞噬指数=(2 000个MΦ吞菌总数/2 000个MΦ)×100%
1.2.4 IL-2作用M5侵染的巨噬细胞的细胞因子释放的时效关系 试验组分为阴性对照组(正常细胞)、空白对照组(M5 侵染)、不同浓度IL-2调节组(IL-2+M5侵染),用麦氏比浊管法调整菌液浓度,按100∶1的比例在6孔板中用M5侵染MΦ,分别加入0、1×106、3×106、5×106UI的IL-2,将其置于37℃、体积分数为5%CO2的细胞培养箱中培养,分别在培养0、2、6、12、24 h时,收集细胞培养上清液10 μL,然后用ELISA试剂盒测定TNF-α、NO 和IFN-γ的浓度大小。
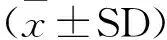
2 结果
2.1 布鲁菌M5侵染巨噬细胞后释放各细胞因子的时效关系
为了排除培养液(主要是血清中)中的干扰,故设阴性对照组,表1中皆是减去阴性对照所得数据。布鲁菌M5侵染巨噬细胞后,释放了TNF-α、IFN-γ、NO等因子,M5刺激后2 h,TNF-α升高,6 h达到峰值12.360 ng/L±1.202 ng/L,与M5刺激0 h比较,呈极显著差异(P<0.01);M5侵染24 h时,IFN-γ达到峰值24.524 ng/L±0.621 ng/L(P<0.01);在M5侵染2 h时,NO含量持续升高,且在12 h时达到峰值6.424 ng/L±0.545 ng/L(P<0.01)。

表1 腹腔巨噬细胞(MΦ)被布鲁菌M5侵染后释放细胞因子的时效关系
注:同列数据肩标相邻大写字母表示差异显著(P<0.05),肩标相间大写字母表示差异极显著(P<0.01),肩标相同大写字母或无字母标注表示差异不显著(P>0.05)。
Note:In the same column, values with adjacent capital letters mean significant difference (P< 0.05),with alternate capital letters mean extremely significant difference (P< 0.01),and with the same or no letters mean no significant difference (P> 0.05).
2.2 IL-2作用下巨噬细胞胞内活菌数
由表2可知,布鲁菌M5侵染细胞0 h时,在各不同含量组IL-2作用下,与对照组0IU相比,吞噬指数均无显著差异(P>0.05);但在2、6、12、24 h时,不同含量IL-2作用下,吞噬指数均呈显著差异(P<0.05)。
相同IL-2含量条件下,各培养不同时间的MΦ,与对照组0 h相比,吞噬指数均呈极显著差异(P<0.01);在2 h,IL-2含量为5×106IU时,吞噬指数达到最大值,高于相邻两组,极显著高于对照组(P<0.01)。表明IL-2的免疫调节功能具有双向性;对巨噬细胞的吞菌能力有明显的促进作用,并存在浓度和时间上的依赖性。
2.3 IL-2对布鲁菌侵染腹腔巨噬细胞后释放TNF-α的时效关系
由表3可知,在0、6、24 h时,在不同含量组IL-2作用下,与对照组0IU相比,均无显著差异(P>0.05);在2 h和12 h时,MΦ 分泌的TNF-α 因子随着IL-2含量的增加为增长,且当IL-2含量为5×106IU 时,均显著高于对照组0 h的含量(P<0.05),尤以2 h,IL-2 含量为5×106IU条件下,细胞释放TNF-α的效果达到最佳(P<0.01)。表明IL-2可以促进MΦ增殖或者活化,使其分泌TNF-α作用增强,并存在含量依赖性。
当IL-2含量为1×106IU和3×106IU 时,培养24 h时,血清中TNF-α 的含量显著高于对照组0 h的含量(P<0.01),且以含量为1×106IU时,血清中TNF-α含量最大;当IL-2含量为5×106IU 时,培养2 h和12 h的MΦ释放TNF-α因子的含量均极显著高于对照组(P<0.01),且培养2 h的释放量最大,效果最佳,极显著高于相邻两组(P<0.01)。表明了IL-2能使MΦ 活化,促进MΦ 分泌TNF-α,启动机体免疫应答,并存在着双向调节性和时间依赖性。

表2 IL-2对被M5侵染的巨噬细胞的吞噬能力的调节作用
注:①同行数据肩标相邻小写字母表示差异显著(P<0.05),肩标相间小写字母表示差异极显著(P<0.01),肩标相同小写字母或无字母标注表示差异不显著(P>0.05);② 同列数据肩标相邻大写字母表示差异显著(P<0.05),肩标相间大写字母表示差异极显著(P<0.01),肩标相同大写字母或无字母标注表示差异不显著(P>0.05)。
Note:① In the same row, values with adjacent small letters mean significant difference (P< 0.05),with alternate small letters mean extremely significant difference (P< 0.01),and with the same or no letters mean no significant difference (P> 0.05);② In the same column, values with adjacent capital letters mean significant difference (P< 0.05),with alternate capital letters mean extremely significant difference (P< 0.01),and with the same or no letters mean no significant difference (P> 0.05).
表3 IL-2调节作用下被M5侵染的腹腔巨噬细胞释放TNF-α的时效关系
注:①同行数据肩标相邻小写字母表示差异显著(P<0.05),肩标相间小写字母表示差异极显著(P<0.01),肩标相同小写字母或无字母标注表示差异不显著(P>0.05);②同列数据肩标相邻大写字母表示差异显著(P<0.05),肩标相间大写字母表示差异极显著(P<0.01),肩标相同大写字母或无字母标注表示差异不显著(P>0.05)。
Note:① In the same row, values with adjacent small letters mean significant difference (P< 0.05),with alternate small letters mean extremely significant difference (P< 0.01),and with the same or no letters mean no significant difference (P> 0.05);② In the same column, values with adjacent capital letters mean significant difference (P< 0.05),with alternate capital letters mean extremely significant difference (P< 0.01),and with the same or no letters mean no significant difference (P> 0.05).
2.4 IL-2对布鲁菌侵染腹腔巨噬细胞后释放IFN-γ 的时效关系
由表4可知,在2 h和24 h时,在各不同含量组IL-2作用下,与对照组0 IU相比,均无显著差异(P>0.05);IL-2含量为3×106IU和5×106IU时,MΦ分别培养2 h、6 h所释放的IFN-γ均显著高于对照组(P<0.05),尤以6 h,IL-2 含量为3×106IU条件下,细胞释放IFN-γ 的效果达到最佳(P<0.01)。结果显示IL-2对于IFN-γ 的释放有明显的促进作用,一定程度上可以增强机体的免疫应答能力。
同一含量IL-2条件下,培养2 h后,血清中MΦ 释放IFN-γ的含量均显著高于对照组0 h的含量(P<0.05);培养24 h,血清中IFN-γ的含量与对照组相比,均呈极显著差异(P<0.01);相同含量组条件下,IFN-γ的含量随着时间点的变化而变化,尤以24 h,IL-2含量为1×106IU时,IFN-γ的含量极显著高于对照组0 h的含量,达到最大值,MΦ 释放IFN-γ的能力在IL-2的作用下有所提高,并存在着时间依赖性。
表4 IL-2调节作用下被M5侵染的腹腔巨噬细胞释放IFN-γ的时效关系
注:①同行数据肩标相邻小写字母表示差异显著(P<0.05),肩标相间小写字母表示差异极显著(P<0.01),肩标相同小写字母或无字母标注表示差异不显著(P>0.05);②同列数据肩标相邻大写字母表示差异显著(P<0.05),肩标相间大写字母表示差异极显著(P<0.01),肩标相同大写字母或无字母标注表示差异不显著(P>0.05)。
Note:① In the same row, values with adjacent small letters mean significant difference (P< 0.05),with alternate small letters mean extremely significant difference (P< 0.01),and with the same or no letters mean no significant difference (P> 0.05);② In the same column, values with adjacent capital letters mean significant difference (P< 0.05),with alternate capital letters mean extremely significant difference (P< 0.01),and with the same or no letters mean no significant difference (P> 0.05).
2.5 IL-2对布鲁菌侵染腹腔巨噬细胞后释放NO的时效关系
由表5可知,MΦ被M5侵染0 h时,各不同含量组IL-2作用下,血清中NO含量与对照组0 IU相比,均无显著差异(P>0.05);含量组为1×106IU和5×106IU时,分别培养6、12、24 h后,血清中NO含量显著高于对照组0 IU含量(P<0.05);且在IL-2含量为5×106IU,培养12 h血清中NO含量达到最大,MΦ的免疫应答能力达到最佳。
IL-2含量组为1×106IU和5×106IU时,血清中NO含量随着培养时间的增长而增加,且在培养12 h时MΦ 分泌NO的含量显著高于对照组0 h的含量(P<0.05),达到峰值,高于相邻的2个时间点。结果说明在IL-2作用下,MΦ 分泌NO的的能力具有双向调节性,并存在时间依赖性。
表5 IL-2调节作用下被M5侵染的腹腔巨噬细胞释放NO的时效关系
注:①同行数据肩标相邻小写字母表示差异显著(P<0.05),肩标相间小写字母表示差异极显著(P<0.01),肩标相同小写字母或无字母标注表示差异不显著(P>0.05);②同列数据肩标相邻大写字母表示差异显著(P<0.05),肩标相间大写字母表示差异极显著(P<0.01),肩标相同大写字母或无字母标注表示差异不显著(P>0.05)。
Note:① In the same row, values with adjacent small letters mean significant difference (P< 0.05),with alternate small letters mean extremely significant difference (P< 0.01),and with the same or no letters mean no significant difference (P> 0.05);② In the same column, values with adjacent capital letters mean significant difference (P< 0.05),with alternate capital letters mean extremely significant difference (P< 0.01),and with the same or no letters mean no significant difference (P> 0.05).
3 讨论
巨噬细胞是炎症反应过程中的关键免疫细胞,不同炎症反应阶段,巨噬细胞发挥不同甚至相反的功能。本研究中采用不同含量IL-2在不同时间点作用巨噬细胞,发现随着IL-2含量的增加,巨噬细胞的吞噬能力在不断增加[13],且IL-2含量越大,吞噬M5的能力越强,表明IL-2在一定程度上对巨噬细胞的吞噬能力有促进作用。在时间点上虽然也存在一定的依赖性,但胞内活菌数呈现一定的“波浪式”规律,大胆猜想认为IL-2对巨噬细胞的增殖或凋亡能力也有一定的影响,有待于进一步研究。
TNF-α是一个具有多重功能的细胞因子,具有免疫调节作用[14],在炎症反应、机体防御中起着关键的作用。TNF在肿瘤作用方面有利有弊:①TNF是一个内源性的肿瘤促进因素,它可以刺激肿瘤细胞生长[15],增殖,侵袭,转移,血管生成;②TNF又有杀肿瘤细胞的作用。活化后的MΦ能分泌大量的趋化因子和炎症因子,参与机体固有和获得性免疫调节。近年来的一些研究表明,近百种的活性物质被活化的MΦ产生,如TNF-α、NO等,其中许多与免疫应答及炎症有关[16]。TNF-α的出现能诱发机体代谢和血流动力学明显变化,促进炎症介质生成。IFN是一种广谱抗病毒剂,但它并不直接作用病毒,使细胞产生抗病毒蛋白,主要是通过细胞表面受体作用,以此增强巨噬细胞的活力,起到免疫调节作用[17-18]。IFN是一组有多种功能的活性蛋白质(主要是糖蛋白),它们在同种细胞上具有广谱的抗病毒、影响细胞生长,以及分化、调节免疫功能等多种生物活性[19]。有研究发现IFN-γ可显著增强MΦ FcγR表达,以此提高MΦ的吞噬能力。另有资料表明单独用IFN-γ刺激MΦ时,在一定剂量范围内可使MΦ由静止态转变为应答态,成为致敏性的MΦ,然后在脂多糖或某些细胞因子作用下转变为活化的MΦ。NO能够降低疼痛和一些炎症反应,具有很强的抗炎作用。NO主要由NO合酶(NOS)催化产生,诱生型NO合酶存在MΦ内,在受到抗原或LPS等激活时,MΦ产生IL-1、TNF-α等因子[20],以自分泌或旁分泌方式协同LPS诱导MΦ中iNOS的表达;Erdogan等研究发现大鼠布鲁菌感染模型中,肝脏和脾脏中NO浓度显著升高,而血浆中并不发生显著变化。
通过本试验数据分析发现,在IL-2浓度组为5×106IU作用下,MΦ被M5侵染2 h时,血清中TNF-α的含量达到最大值,大大提高了巨噬细胞的免疫应答作用,表明了IL-2能够促进MΦ活化,加强其分泌因子的能力,且存在着时间和浓度上的依赖性,还能够启动炎症反应的发生。在IL-2的调节作用下,血清中IFN-γ的含量无论从时间上还是IL-2浓度上,都可以看出,与对照组相比有显著性的增加,从另一方面也证明了IL-2可以增强MΦ的免疫作用;但是这个显著性的增加并没呈现严格规律性,可能与一些其他因子的影响有关,如IL-6,有待于进一步的探讨研究;在IL-2作用下,血清中NO含量存在差异性,但这种差异性并不是很大,即在一定程度上IL-2可以促进MΦ的活化,对NO的产生的作用是有限的,对机体免疫力的提高并没有所期望的效果。
本研究结果表明,IL-2可以促进MΦ活化,刺激MΦ使其分泌TNF-α、NO、IFN-γ的能力增强,参与抗布鲁菌的免疫应答,从而为临床运用IL-2研究及治疗布病提供新的思路和理论依据。IL-2能在一定程度上增强巨噬细胞的吞噬能力,存在着含量依赖性,且在含量和时间点亦具有双向调节性,含量过高时反而有一定程度的抑制作用,可能是由于IL-2对巨噬细胞的增殖或凋亡能力也有一定的影响,有待于进一步研究。
参考文献:
[1] 翟志敏.IL-2对免疫激活和免疫耐受的双向调节作用[J].中国药理学通报,2013,29(3):319-322.
[2] 王媛媛,李兆明,李旭东,等.IL-2、IL-6及TNF-α在结外NK/T细胞淋巴瘤鼻型患者血清中的表达及临床意义[J].中国癌症杂志,2015,25(5):377-381.
[3] 亓文静,李 岩,王 玉,等.IL-2通过Jak3-Stat5通路促进巨噬细胞M1极化[J].基础医学与临床,2015,35(8):1055-1060.
[4] 王若天,孟明耀,李 鹏,等.IL-2和IL-15诱导的CIK细胞联合化疗治疗直肠和结肠癌的临床疗效[J].昆明医科大学学报,2014,35(11):97-101.
[5] 贾春丽,杨 颖,李志鹏,等.IL-2联合顺铂治疗恶性胸腔积液的有效性及安全性Meta分析[J].海南医学,2017,28(5):839-843.
[6] 杨荣敏,段相国,陈 建,等.血清IL-2、TNF-α及IL-13在类风湿关节炎诊断与治疗中的临床意义[J].现代免疫学,2017,37(1):44-49.
[7] Roviello G,Zanotti L,Correale P,et al.Is still there a role for IL-2 for solid tumors other than melanoma or renal cancer?[J].Immunotherapy,2017,9(1):25-32.
[8] Abhishek K S,Chakravarti A,Baveja C P,et al.Association of interleukin-2,-4 and -10 with dengue severity.[J].Indian J Pathol Microbiol,2017,60(1):66-69.
[9] 熊一全,黄 韬,赵向旺,等.白细胞介素-2增强淋巴细胞对乳腺癌细胞的杀伤作用[J].中华实验外科杂志,2015,32(5):990-992.
[10] Atluri V L,Xavier M N,Jong M F D,et al.Interactions of the human pathogenicBrucellaspecies with their hosts[J].Annu Rev Microbiol,2011,65(1):523-541.
[11] 李 高,蔡仁中,蔡用清,等.肺癌患者血清中IL-2、IFN-γ、TNF-α、NKG2D的表达及临床意义[J].海南医学,2015,26(12):1728-1731.
[12] 戴 春,周永春,黄云超,等.非小细胞肺癌患者血清中细胞因子(IL-2、IL-4、IL-6、IL-10)的含量与临床分期的关系[J].现代肿瘤医学,2015,23(6):779-781.
[13] 李桉琪,祁元明,周哲骏,等.不同浓度IL-2对体外诱导肽特异性细胞毒性T淋巴细胞培养体系的影响[J].中国癌症杂志,2016,26(6):756-762.
[14] Lin X H,Kashima Y,Khan M,et al.Short comminication:An immunohistochemical study of TNF-α in optic nerves from AIDS patients[J].Curr Eye Res,1997,16(10):1064-1068.
[15] Starke R M,Chalouhi N,Ali M S,et al.190 critical role of TNF-α in cerebral aneurysm formation and rupture[J].Neurosurgery,2013,60:183.
[16] 陈 英,张文玲,黄 涛,等.炎症因子TNF-α、IL-6、IL-17与类风湿关节炎并发动脉粥样硬化的关系[J].免疫学杂志,2017,33(3):268-272.
[17] 涂 浩,赵星灿,杨占娜,等.猪重组IL-2、IL-4和IFN-γ对口蹄疫合成肽疫苗的免疫增强作用研究[J].畜牧兽医学报,2015,46(8):1390-1399.
[18] Tanaka S,Furuya K,Yamamoto K,et al.Procyanidin B2 gallates inhibit IFN-γ and IL-17 production in T cells by suppressing T-bet and RORγt expression[J].Int Immunopharmacol,2017,44:87-96.
[19] Liu W,Liu X,Luo M,et al.dNK derived IFN-γ mediates VSMC migration and apoptosis via the induction of LncRNA MEG3:A role in uterovascular transformation[J].Placenta,2017,50:32-39.
[20] 邝高艳,严 可,柴 爽,等.加味独活寄生合剂治疗膝骨关节炎临床疗效及对关节液中IL-1,IL-6,TNF-α及NO的影响[J].中国实验方剂学杂志,2017,23(1):174-178.
猜你喜欢
当代水产(2022年1期)2022-04-26
中老年保健(2021年3期)2021-08-22
现代临床医学(2021年4期)2021-07-31
天津医科大学学报(2021年2期)2021-03-29
现代临床医学(2021年1期)2021-01-26
西南农业学报(2016年5期)2016-05-17
西南农业学报(2016年6期)2016-04-16
中西医结合心脑血管病杂志(2016年20期)2016-03-01
医学研究杂志(2015年12期)2015-06-10
现代检验医学杂志(2014年1期)2014-02-06
