
高温干旱复合胁迫及复水对刺槐幼苗水分运输的影响
2018-05-14 05:48刘璇吴永波邵维
生态科学 2018年2期
刘璇,吴永波,邵维
南京林业大学,江苏省南方现代林业协同创新中心,南京 210037
1 前言
IPCC第五次评估报告明确指出,21世纪末全球表面温度变化可能超过1.5℃,2100年之后将持续变暖,干湿地区的降水差异增大[1],平均降水量变化出现显著空间差异[2]。随着全球变暖和气候异常,特别是当高温和少雨相伴发生时,高温胁迫发生的频率和范围很可能增大[3]。
植物在生长发育过程中通过调整水力结构影响导水阻力和水分需求,并对水分运输和水分平衡产生影响。王丁[4]的研究表明,干旱胁迫下刺槐的水分运输受到较大影响,且复水之后恢复程度较小。在众多被环境因素抑制的细胞机能中,光合作用被公认为是特别敏感的生理过程[5]。短时高温条件下快速干旱,可使叶片气孔导度下降,进而引起净光合速率降低[6]。有研究发现植物在干旱复水后存在补偿效应,会进行自我调节恢复到正常水平,即植物在干旱复水以后会出现生长加快、光合、蒸腾速率提高等现象[7],但高温干旱复合胁迫及复水对植物水力结构及光合作用的研究较少。
刺槐(Robinia pseudoacaciaLinn.)作为豆科固氮植物,因生态幅宽、适应性强、生长快、具有良好的适应性和抗逆性[8]等特性在我国被当成抗旱的先锋树种被广泛种植,对干旱地区植被的恢复和重建意义重大[9],是干旱、半干旱地区主要的造林树种之一。有研究表明干旱程度与刺槐叶水势呈负相关[10];水分胁迫导致刺槐叶水势先降低后升高,解除水分胁迫后,刺槐叶水势会发生补偿效应来减小水分胁迫的伤害[11]。目前刺槐水分运输的研究主要集中在干旱胁迫,而高温干旱复合胁迫对刺槐水分运输影响的研究较少。
作为全球气候变化的两个重要表征,亦最可能同时出现的两个因子,高温和干旱在影响植物分布方面起着决定性作用[12],而高温和干旱往往同时发生。本文通过在不同高温和干旱胁迫下对刺槐幼苗导水率、比导率的测定,并结合气孔导度和蒸腾速率的变化,试图分析高温干旱复合胁迫以及复水对刺槐幼苗水分传输过程的影响。
2 试验设计与测定指标
2.1 试验设计
试验材料为1年生刺槐幼苗,平均株高78.6 cm;土壤为棕壤土,pH 6.3,田间持水量24.8%,水解氮114.28 mg·kg-1, 速效磷 3.45 mg·kg-1, 速效钾 285.71 mg·kg-1;试验用盆规格 32 cm×27 cm×28 cm。试验在FYS-8智能人工气候室(相对湿度60%;最大光合有效辐射 1800 μmol·m-2·s-1)内进行。不同气候室分别设置为25℃(T1,CK)、33℃(T2,轻度高温胁迫)、40℃(T3,中度高温胁迫)3个不同温度。试验共设3个水分梯度分别为田间持水量的100%(D1,CK)、70%(D2,轻度干旱胁迫)、40%(D3,中度干旱胁迫)。根据设定的土壤含水率计算每盆重量,作为标准盆重,采用称重法进行水分控制。共T1D1、T1D2、T1D3、T2D1、T2D2、T2D3、T3D1、T3D2、T3D3 9个处理,其中T1D1为对照(CK)。每个处理3个重复,一共27盆。
2.2 测定指标与方法
在试验期间,每周测定1次光合作用性能指标,于每次上午10:00用Li-6400 XT便携式光合仪测定刺槐幼苗蒸腾速率(Tr)、气孔导度(Gs),每个指标重复3次。
在实验结束前测定刺槐幼苗导水率及边材比导率,采用王明浩[13]的方法。环切刺槐幼苗茎段枝条两端表皮,露出木质部,用封口膜绕两端处数圈,利用塑胶管将离体茎段、移液管、三通阀、量筒连接,量筒中水柱产生一定水压,迫使植物离体茎段产生水流,稳定5 min后记录单位体积水流量所用时间,计算水流速度F(kg·s-1),同时将茎段近主枝端用1%番红染色2 h,再用游标卡尺测量茎段横截面直径和心材直径,并计算茎段边材横截面积。
导水率(Kh)=F·L·ΔP-1; 边材比导率(Ks)=Kh·Aw式中F为通过离体茎段的水流速度,L为茎段长度,ΔP为引起该茎段水流动的压强,Aw为茎段边材横截面积。
2.3 数据处理
用IBM SPSS Statistic 19.0进行数据处理和差异显著性分析,用SigmaPlot进行制图分析。
3 结果与分析
3.1 不同处理对刺槐幼苗导水率、比导率的影响
由图1可知,Ks在高温干旱复合胁迫下总体表现为下降趋势,不同高温胁迫下的最低值均出现在田间持水量40%水平。25℃、33℃时,Ks均在田间持水量70%时上升而在田间持水量为40%时下降明显(P<0.05);40℃时,Ks表现为持续下降并在田间持水量40%时达到最低。田间持水量40%时处理的幼苗Ks分别比田间持水量100% 时降低80.5%(25℃)、94.1%(33℃)、86.0%(40℃),仅为田间持水量 70% 的 31.4%(25℃)、3.1%(33℃)、45.9%(40 ℃)。
Kh在25℃时只在田间持水量40%时有明显的下降趋势;33℃时表现为田间持水量70%时上升,田间持水量40%时下降;40℃时随干旱胁迫的增加下降明显(P<0.05)。Kh与Ks一致,25℃、33℃、40℃时的最低值均出现在田间持水量40%水平,分别为田间持水量100%的18.9%(25℃)、5.1%(33℃)、14.1%(40℃),这可能是因为复合胁迫加剧了Ks、Kh的下降。方差分析结果(表1)也表明:干旱对Ks、Kh的影响差异显著(P<0.05),高温胁迫对Ks、Kh的影响差异不显著。
3.2 不同处理对刺槐幼苗气孔导度与蒸腾速率的影响
由图2可知,Gs在25℃、33℃、40℃均随着干旱胁迫程度增加而表现田间持水量70%时升高、田间持水量40%时下降。在田间持水量40%时的水平,Gs除在25℃下接近于田间持水量100%外,33℃、40℃均高于田间持水量 100%水平,分别上升了15.1%(33℃)、23.4%(40℃)。高温干旱复合胁迫(33℃下田间持水量70%和40℃下田间持水量70%)促进了Gs增加,随着复合胁迫中干旱胁迫程度增加(33℃下田间持水量40%和40℃下田间持水量40%),Gs下降,但仍高于对照,表明高温干旱复合胁迫对Gs有一定的促进作用。

图1 高温干旱下刺槐幼苗Ks、Kh的变化Fig.1 Ksand Khof Robinia pseudoqcacia Linn.seedling under the combined stress of elevated temperature and drought

图2 高温干旱下刺槐幼苗Gs、Tr变化Fig.2 Gs and Tr of Robinia pseudoqcacia Linn.seedling under the stress of elevated temperature and drought
Tr在25℃、33℃下均表现为随着干旱胁迫程度的增加,在田间持水量70%时升高、田间持水量40%时下降,在40℃下,表现为在干旱胁迫下缓慢上升。由图2可知,Tr在高温单一胁迫下上升明显(P<0.05),在干旱单一胁迫(25℃)下下降。复合胁迫下,Tr在田间持水量40% 处理水平,与对照(25℃下田间持水量100%)相比,分别上升了63.2%(33℃)、64.3%(40℃),表明高温干旱复合胁迫对Tr存在促进作用,与Gs结果一致。方差分析结果(表1)也表明:高温对Tr的影响存在差异显著性(P<0.05),对Gs的影响差异不显著,干旱胁迫对Tr、Gs的影响存在差异显著性(P<0.05)。
3.3 不同处理下刺槐幼苗各个指标之间的方差分析
对不同处理下刺槐幼苗各项指标进行方差分析结果表明(表1),高温处理下Tr差异显著(P<0.05),Gs、Kh、Ks差异不显著(P>0.05);干旱处理下,Tr、Gs、Ks、Kh差异显著(P<0.05)。
3.4 复水对刺槐幼苗水力结构参数、气孔导度、蒸腾速率的影响
在胁迫4周后对幼苗进行了为期2周的复水处理,结果表明(表2),复水后,刺槐幼苗茎段Kh、Ks变化趋势基本一致:25℃时Kh、Ks恢复且显著高于胁迫前;33℃、40℃时,Kh、Ks持续降低。在高温干旱复合胁迫(33℃下田间持水量40%)复水后Ks、Kh变化不明显,这可能是因为胁迫已导致Kh和Ks降至最低,植物的水分运输结构可能已经遭到破坏。Tr、Gs在复水后明显上升且随胁迫程度的增加恢复程度增加,说明复水有利于促进植物的水分传输,也在一定程度上对植物光合作用有补偿作用。

表1 不同处理下各项指标的方差分析Tab.1 Variance analysis on various indicators under different treatments
4 讨论
复合胁迫对植物的影响不同于单一胁迫[14],高温干旱复合胁迫效应并不是两个因子单一胁迫的简单相加,而是相互作用,其响应基因和代谢途径[15]也不同于单一胁迫。植物对一种因子胁迫的响应可以改变其他因子的胁迫程度[16]。高温干旱复合胁迫中干旱虽然是影响植物水分传输的直接胁迫因子,但其危害严重度很大程度上取决于温度[17]。
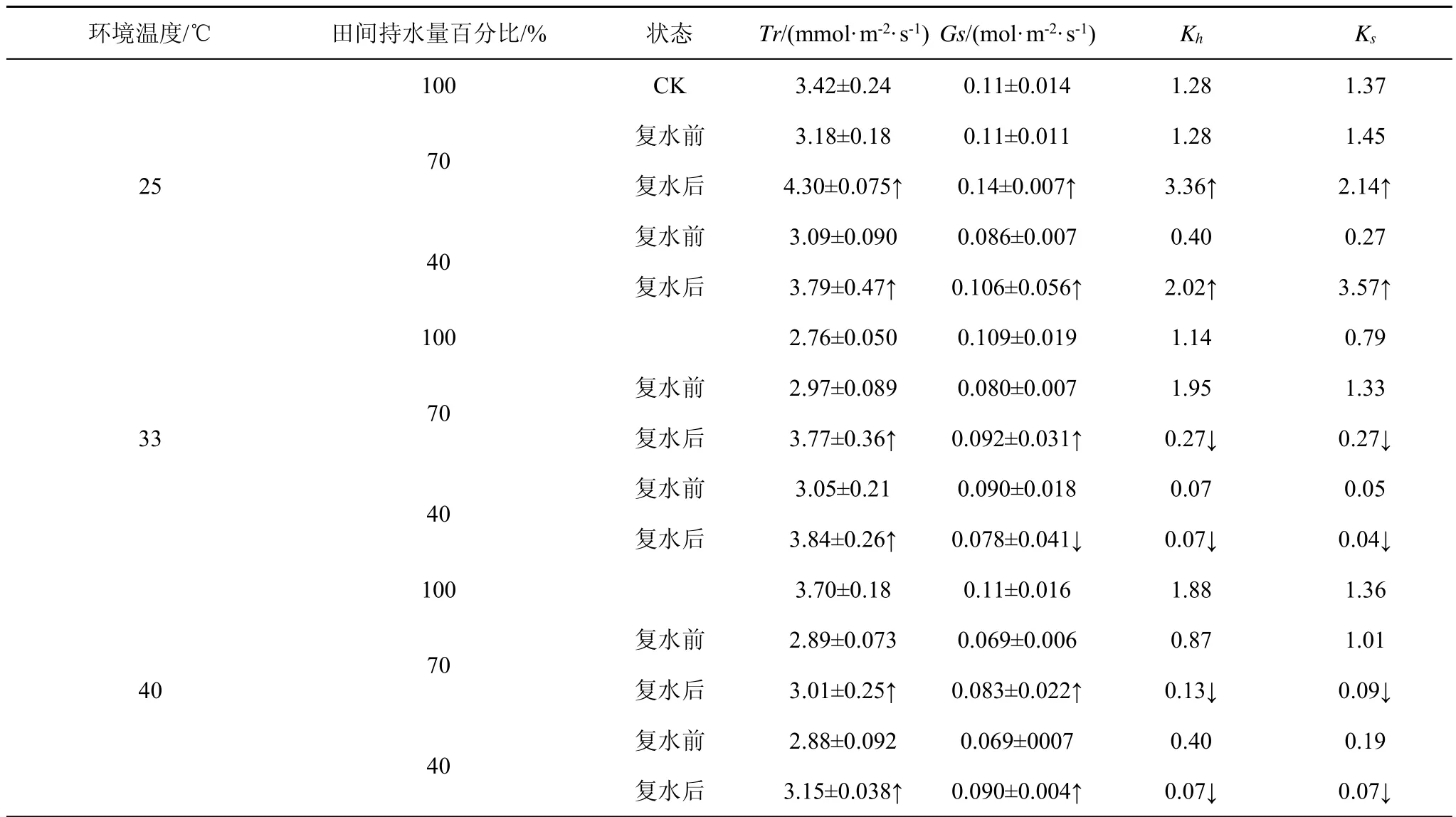
表2 复水前后刺槐幼苗水分传输参数值比较Tab.2 The values of water transportation parameters of Robinia pseudoqcacia Linn.seedling before and after rewatering
本研究中,同一干旱胁迫下温度升高,叶片蒸腾速率增加,植物发生水分亏缺,导水率呈现下降趋势,这与Aasamaa等[18]对山杨树的研究结果类似。刺槐属于低水势忍耐脱水耐旱树种,导管直径大,输水阻力小,易发生栓塞现象[19]。在一定水势范围内,植物通过栓塞机制降低导水率以限制水分丧失,提高其干旱环境下的生存能力,但当导水率降低到一定程度后,栓塞会对其正常生理活动造成危害[20]。同一高温胁迫下,刺槐茎段导水率、比导率随着干旱胁迫增加总体呈下降趋势,胁迫强度越大,下降越明显,这与翟洪波等[21]的研究结果一致。
在本研究中高温单一胁迫下气孔导度、蒸腾速率上升;干旱单一胁迫下气孔导度、蒸腾速率下降。在不同程度高温干旱胁迫下气孔导度与蒸腾速率表现不同:轻度高温干旱胁迫下气孔导度、蒸腾速率上升,说明植物可能对高温进行一定的防御反应;随高温胁迫进一步加剧,气孔导度、蒸腾速率有所下降,这可能是蒸腾降温也给植物带来了不良影响,如水分的入不敷出导致植物缺水萎蔫,而水分短缺又相伴着气孔的关闭,蒸腾速率的下降[22]。复合胁迫下气孔导度、蒸腾速率接近或略高于高温或干旱单一胁迫,说明高温可能在复合胁迫中对气孔导度、蒸腾速率起到了主导作用,高温与干旱之间有可能存在相互抵消作用,一方面,随着水分胁迫的加剧,木质部形成栓塞,导水率下降,蒸腾速率加强,叶水势下降,使叶片内水供应不足;另一方面,当植物根部感知水分胁迫时,水势降低,分泌脱落酸,脱落酸向上运输引起气孔的部分关闭[23],导致气孔导度下降。
研究表明,干旱胁迫与高温胁迫伤害植物的本质存在区别:干旱胁迫是由于植物失水造成的质膜损伤[24];高温胁迫则是由于高温引起植物蒸腾作用失水过度导致细胞脱水损伤[25];与单一干旱胁迫相比,高温胁迫有加强水分胁迫的效应[26]。本研究结果显示高温干旱复合胁迫导致导水率、比导率下降,且复合胁迫中干旱胁迫越大,比导率、导水率下降幅度越大。与单一胁迫相比,复合胁迫下刺槐幼苗水分运输所受影响较大,这可能是高温与干旱的交互作用对比导率、导水率产生一定的叠加影响,进一步破坏了渗透调节和水的关系[27]。本研究中高温单一胁迫下气孔开放,蒸腾作用加强;干旱单一胁迫下气孔关闭,蒸腾作用随之降低。在轻度高温干旱复合胁迫下,气孔导度、蒸腾速率增加说明适度高温会促进光合性能上升[28],随着植物干旱胁迫梯度增加,植物光合性能会逐渐降低,这可能是因为短时间高温条件下快速干旱,可使叶片气孔导度下降,进而引起光合作用速率降低[6]。
本研究结果表明,干旱单一胁迫下的导水率、比导率在复水后均有所恢复,但复合胁迫下导水率、比导率在复水后基本无变化,这可能是因为复合胁迫下高温加剧了干旱对植物水分运输的影响,对植物的水分运输结构造成了一定损伤,即使复水亦不能恢复这种损伤。复水后刺槐幼苗气孔导度、蒸腾速率均比复水前明显升高,但仍低于对照,与前人研究结果[29]一致,说明在土壤水分状况改善后,刺槐幼苗叶片部分损伤得到修复,从而使光合性能得到部分恢复[30]。
5 结论
植物在高温或干旱胁迫下其体内的水分运输系统会通过自身的调节来保障植物正常的生长发育。相对于单一胁迫,高温干旱复合胁迫下植株导水率、比导率下降更为显著。植株的气孔导度、蒸腾速率在胁迫下均有所上升,且受高温影响较大;复合胁迫下对其的影响介于两种单一胁迫之间。复水有利于恢复单一干旱胁迫下刺槐幼苗的比导率和导水率,而复合胁迫下恢复效果不明显。
参考文献
[1]ALEXANDER L,SIMON BINDOFF N L.Working Group I Contribution to the IPCC Fifth Assessment Report Climate Change 2013:The PhysicalScience Basis Summary for Policymakers[R].IPCC,2013.
[2]董思言,高学杰.长期气候变化——IPCC第五次评估报告解读[J].气候变化研究进展,2014,10(1):56–59.
[3]SALVUCCI M E,CRAFTS-BRANDNER S J.Inhibition of photosynthesis by heat stress:the activation state of Rubisco as a limiting factor in photosynthesis[J].Physiologia plantarum,2004,120(2):179–186.
[4]王丁,杨雪,韩鸿鹏,等.干旱胁迫及复水对刺槐苗水分运输过程的影响[J].南京林业大学学报(自然科学版),2015,29(1):67–72.
[5]J BERRYA,BJORKMAN O.Photosynthetic Response and Adaptation to Temperature in Higher Plants[J].Annual Review of Plant Physiology,1980,31(1):491–543.
[6]张往祥,曹福亮.高温期间水分对银杏光合作用和光化学效率的影响[J].林业科学研究,2002,15(6):672–679.
[7]胡田田,康绍忠.植物抗旱性中的补偿效应及其在农业节水中的应用[J].生态学报,2005,25(4):885–891.
[8]高艳明,李建设,高娜,等.刺槐组织培养繁殖技术[J].林业科技,2005,30(1):9–10.
[9]尹达,杜宁,徐飞,等.外来物种刺槐(Robinia pseudoacaciaL.)在中国的研究进展[J].山东林业科技,2014,44(6):92–99.
[10]李继文,王进鑫,张慕黎,等.干旱及复水对刺槐叶水势的影响[J].西北林学院学报,2009,24(3):33–36.
[11]徐飞,郭卫华,徐伟红,等.刺槐幼苗形态、生物量分配和光合特性对水分胁迫的响应[J].北京林业大学学报,2010,32(1):24–30.
[12]GODOY O,DE LEMOS-FILHO J P,VALLADARES F.Invasive species can handle higher leaf temperature under water stress than Mediterranean natives[J].Environmental and Experimental Botany,2011,71(2):207–214.
[13]王明浩,张晓玮,王婧如,等.一种简易准确测定木质部导水率的新方法[J].植物生理学报,2013,49(3):297–300.
[14]APRILE A,HAVLICKOVA L,PANNA R,et al.Different stress responsive strategies to drought and heat in two durum wheatcultivars with contrasting water use efficiency[J].BMC Genomics,2013,14(1):1–37.
[15]MITTLER R.Abiotic stress,the field environment and stress combination[J].Trends in Plant Science,2006,11(1):15–19.
[16]刘振山.小麦苗期干旱、高温和旱热共胁迫转录表达谱及ABD部分同源基因表达分化分析[D].北京:中国农业大学,2015.
[17]OLUFAYO A,BALDY C,RUELLEB P,et al.Diurnal course of canopy temperature and leaf water potential of sorghum (SorghumbicolorL. Moench) under a Mediterranean climate[J]. Agricultural and Forest Meteorology,1993,64(3/4):223–236.
[18]AASAMAA K,ÜLONIINEMETS,S BER A.Leaf hydraulicconductance in relation to anatomicaland functional traits during Populus tremula leaf ontogeny[J].Tree Physiology,2005,25(11):1409–1418.
[19]刘晓燕,李吉跃,翟洪波,等.从树木水力结构特征探讨植物耐旱性[J].北京林业大学学报,2003,23(3):48–54.
[20]张硕新,申卫军,张远迎,等.几个抗旱树种木质部栓塞脆弱性的研究[J].西北林学院学报,1997,12(2):2–7.
[21]翟洪波,李吉跃,姜金璞.干旱胁迫对油松侧柏苗木水力结构特征的影响[J].北京林业大学学报,2002,24(Z1):45–49.
[22]POCKMANW T,SPERRY J S.Vulnerability to xylem cavitation and the distribution of Sonoran Desert vegetation[J].American Journal of Botany,2000,87(9):1287–1299.
[23]TYREE M T,ZIMMERMANN M H.Xylem structure and the ascent of sap[M].Springer Science&Business Media,2013.
[24]孙梅霞,祖朝龙,徐经年.干旱对植物影响的研究进展[J].安徽农业科学,2004,32(2):365–367.
[25]李衍素,高俊杰,陈民生,等.高温胁迫对豇豆幼苗叶片膜伤害与保护性物质的影响[J].山东农业大学学报(自然科学版),2007,38(3):378–382.
[26]刘琴,孙辉,何道文.干旱和高温对植物胁迫效应的研究进展[J].西华师范大学学报(自然科学版),2005,26(4):364–368.
[27]MACHADO S,PAULSEN G.Combined effects of drought and high temperature on water relations of wheat and sorghum[J].Plant and Soil,2001,233(2):179–187.
[28]XU Xiao,YANG Fan,XIAO Xiangwen,et al.Sex-specific responses of Populus cathayana to drought and elevated temperatures[J].Plant,Cell&Environment,2008,31(6):850–860.
[29]王磊,张彤,丁圣彦.干旱和复水对大豆光合生理生态特性的影响[J].生态学报,2006,26(7):2073–2078.
[30]郭丽丽,刘改秀,王丽娜,等.逐渐干旱和复水对牡丹光合特性的影响[J]. 上海农业学报, 2014, 30(5): 105–108.
猜你喜欢
农业工程学报(2022年3期)2022-04-16
西南农业学报(2021年7期)2021-08-23
节水灌溉(2021年5期)2021-05-26
生态学报(2021年6期)2021-04-25
农业工程学报(2021年3期)2021-04-15
现代园艺(2020年2期)2020-12-22
节水灌溉(2020年9期)2020-09-16
中国农业科技导报(2020年12期)2020-03-15
小溪流(故事作文)(2018年10期)2018-12-28
现代园艺(2018年2期)2018-03-15
