
贵州省马尾松主要群体的遗传多样性分析
2018-04-20 09:58杨雪梅朱亚艳陈庭巧谢维斌
中南林业科技大学学报 2018年5期
杨雪梅 ,赵 杨 ,2,朱亚艳 ,陈庭巧,谢维斌
(1.贵州大学 林学院,贵州 贵阳 550025;2. 贵州省森林资源与环境研究中心,贵州 贵阳 550025;3.都匀马鞍山国有林场,贵州 都匀 558000)
马尾松Pinus massoniana是我国特有的针叶树种,以其分布广、适应性强、速生、用途广、材质优良而成为我国工业用材极其重要的树种之一。多年来,育种工作者按照种质资源收集与保存、优树选择、营建种子园、控制授粉、测定与选择等常规育种方法进行了大量的遗传改良工作。目前马尾松的遗传改良已经进入到高世代改良阶段,在通过人为选择获得显著的遗传增益的同时,人们也在担心马尾松遗传多样性是否降低。近些年来,利用分子标记技术,尤其是SSR和ISSR分子标记技术进行马尾松天然群体遗传多样性[1-6]、种子园遗传多样性[7-17]、育种群体遗传多样性[18-19]等研究,并有大量的研究报道。如冯源恒等[6]研究发现,广西马尾松的天然群体积累了丰富的遗传变异,遗传多样性主要存在于林分内,广西马尾松第一代育种群体具有较高的遗传多样性,来源于人工林的平均有效等位基因数、遗传多样性指数及观测杂合度都低于天然林[19]。这些研究结果为马尾松遗传改良提供了很好的指导作用。但是,不同研究者多是针对种子园、天然林等特定群体进行研究评价,很少有针对不同改良阶段群体遗传多样性进行对比分析。马尾松分布范围广,在漫长进化的过程中,为了适应复杂多样的自然地理环境,形成了不同的地理变异群体。早期各省马尾松遗传改良基本是按照地域特色、就近取材的原则来进行,不同地域马尾松群体间遗传背景保持一定的独立性[1],且不同省份马尾松遗传改良进度不同,因此,不同省份针对特定群体进行的遗传多样性研究成果很难直接应用于其它省份的马尾松遗传改良工作。
贵州省作为马尾松重要分布区之一,因其独特的位置和地理条件,孕育了遗传品质多样的马尾松资源。目前已建立马尾松种子园4个,优树基因收集区收集保存优良无性系388个,为贵州马尾松的遗传改良奠定了良好的物质基础[17]。目前,贵州省马尾松的育种研究正进入高世代改良阶段。为了实现可持续的多世代改良,本研究以ISSR分子标记为技术手段,对贵州省马尾松主要分布区天然林群体样本、贵州省都匀马尾松种子园(贵州省马尾松良种主要供种基地)基因收集区、种子园及子代进行遗传多样性分析对比,以期为贵州省马尾松种质资源的收集与保存、种子园遗传管理以及马尾松多世代育种策略的制定提供参考依据。
1 材料与方法
1.1 实验材料
在对贵州省马尾松天然林开展资源调查的基础上,根据不同气候,选择了11个面积不低于20 hm2的天然群体为研究对象(见表1)。根据林分面积大小随机选择7~21个单株,单株间距不少于200 m,12个群体共取样162个。
贵州都匀马寨马尾松1.5代无性系种子园始建于1992年,建园材料多来自贵州省的初选优树,每个无性系随机选择单株,共取94个样本。
贵州都匀马寨马尾松良种基地基因收集区始建于1998年,材料来自贵州省初选优树,嫁接保存,每个无性系随机选择1株,取幼嫩针叶作为试验材料,共取80份。
在种子园结实盛期子代苗中随机选择81个单株,所有试验材料分别采集嫩叶,硅胶干燥,低温保存,备用。
1.2 DNA提取与扩增
采用改良的CTAB法提取马尾松的总DNA。所得DNA经过1.0%的琼脂糖凝胶电泳进行检测,将样品稀释至150 ng/µL,备用。
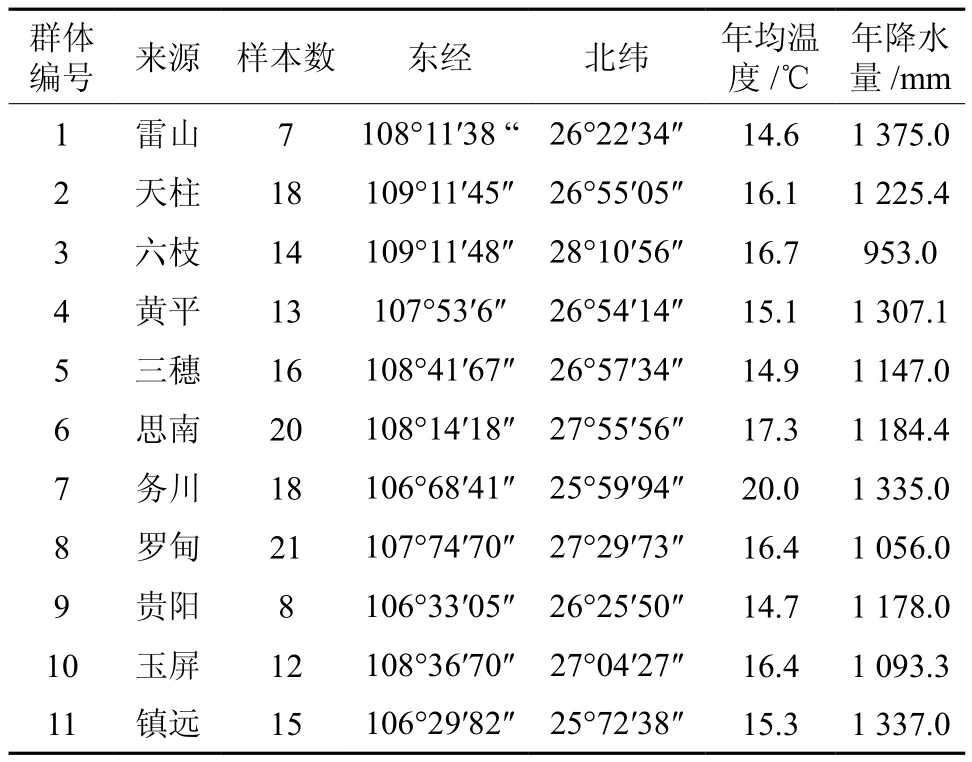
表1 贵州省马尾松天然群体主要地理及生态因子Table 1 The main geographic and ecological factors in natural populations of P. manssoniana in Guizhou province
1.3 引物筛选与PCR扩增
所用引物参照加拿大哥伦比亚大学UBC公司公布的第9套ISSR引物序列,由上海生工生物工程技术服务有限公司合成,筛选出15条多态性引物用于研究,引物序列见表2。 ISSR反应体系的总体积为20 μL, 内含Taq Polymerase 1 U,1×buffer,0.5 mmol dNTP,模板 DNA 约 150 ng,Primer 1 µmol。PCR 反应程序为:94 ℃变性5 min,然后进行40个循环[94 ℃变性45 s,50~60 ℃(视引物而定)退火45 s,72℃延伸90 s]。最后72 ℃延伸7 min,4 ℃终止反应。采用2%琼脂糖凝胶电泳,EB染色,凝胶成像分析系统拍照保存,人工读带。
1.4 电泳结果统计与分析
谱带统计时将具有相同迁移率的扩增片段按0/1系统纪录,有带记为1,无带记为0,对相同分子量的不考虑带的强弱;与空白对照反应有相同迁移率的带均视为污染带,不予记录。应用POPGENE 32软件分群体对供试材料进行遗传参数分析,分别计算不同群体多态位点百分率(PPB)、观测等位基因数(Na)、有效等位基因数(Ne)、Nei’s (1973)基因多样性指数 (H)、Shannon信息指数(I)等。
2 结果与分析
2.1 ISSR多态性引物筛选
通过对引物进行浓度、退火温度、循环次数等实验条件进行ISSR分析,获得15条PCR扩增条带重复性高、多态性好、条带清晰的引物(见表2)。并对11个地区的马尾松天然林样本进行ISSR-PCR扩增,扩增产物经琼脂糖凝胶电泳得到清晰的指纹图谱(见图1)。
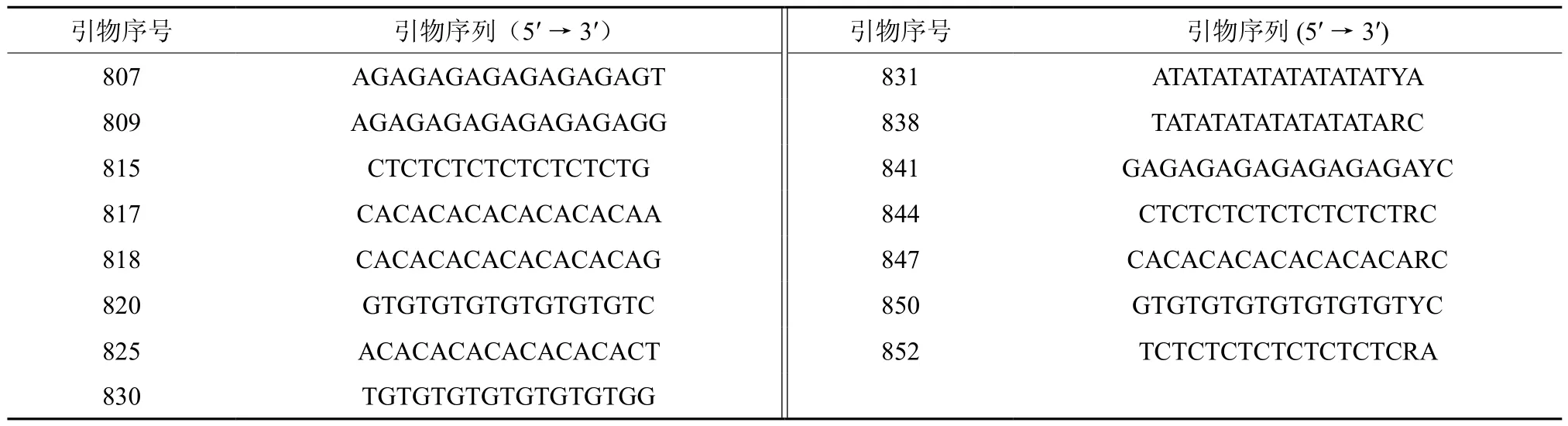
表2 马尾松遗传多样性分析ISSR引物Table 2 The primers of ISSR on the study genetic diversity of masson pine

图1 引物846-天然林ISSR-PCR扩增效果Fig. 1 ISSR-PCR of masson pine by Primer 846
2.2 贵州省马尾松天然群体遗传多样性分析
通过对贵州省马尾松11个天然群体样本进行ISSR分析,15条引物共扩增出110个条带,基因位点多态位点百分率变化介于35.45%~92.73%之间,平均值为71.56%。种源内观测等位基因数变化介于1.3545~1.9273之间,种源间观测等位基因数平均值为1.7107。种源内有效等位基因数变化介于1.1783~1.4612之间,种源间有效等位基因数为1.5601。种源内Nei’s基因多样性的变化介于0.1119~0.2813之间,种源间Nei’s基因多样性平均值为0.3308。种源内Shannon信息指数的变化介于0.1742~0.4332之间,种源间Shannon信息指数为0.5002。观察杂合度偏高,表明群体中杂合体偏多,纯合体偏少;群体间基因分化系数为0.3092,基因流为1.1170。表明贵州省马尾松天然林不同分布区的遗传多样度虽存在明显差异,但仍然保持较高的遗传多样性水平,其中罗甸、三穗、玉屏、镇远等种源遗传多样性较高,而贵阳种源遗传多样性相对较低。由于种源间基因交流频繁,贵州省马尾松变异主要存在于种源内。种源间气候、土壤等差异,在自然选择的作用下种源间也有了较大分化。聚类分析结果(见图2)表明,天柱和思南两个种源的遗传相似度最高,为0.9316。务川、雷山两个种源聚为一类,黄平与六枝2个种源聚为一类,其余7个种源的遗传相似度也比较高。
2.3 都匀马尾松种子园收集区遗传多样性分析
利用PopGen32软件对80个马尾松无性系进行遗传多样性分析,得到多态位点率为71.19%;观测等位基因数为1.7119,有效等位基因数为 1.4606,Nei’s基因多样性指数为 0.2633,Shannon信息指数为0.3925。该收集区遗传多样性水平要低于贵州省马尾松天然群体。
随着生活方式的改变,我国人群中存在MS的比例逐年增加,目前已报道的我国成人MS的比例为9.8%~17.8%[17],本研究中合并MS的前列腺癌患者占16.08%,与之前的报道相符。ZHANG等[12]的研究结果显示MS和前列腺癌的Gleason评分≥8以及较晚的病理分期有关。本研究的结果证实了MS和病理T分期的相关性,然而,未发现两组之间Gleason评分有明显差异,可能的原因是本研究采用的Gleason评分分组方式与之前的研究不同。为了减少其他临床终点对结果的干扰,我们采用竞争风险模型评估MS对生化复发的影响,结果显示合并MS的前列腺癌患者出现生化复发的概率是无MS者的1.86倍。
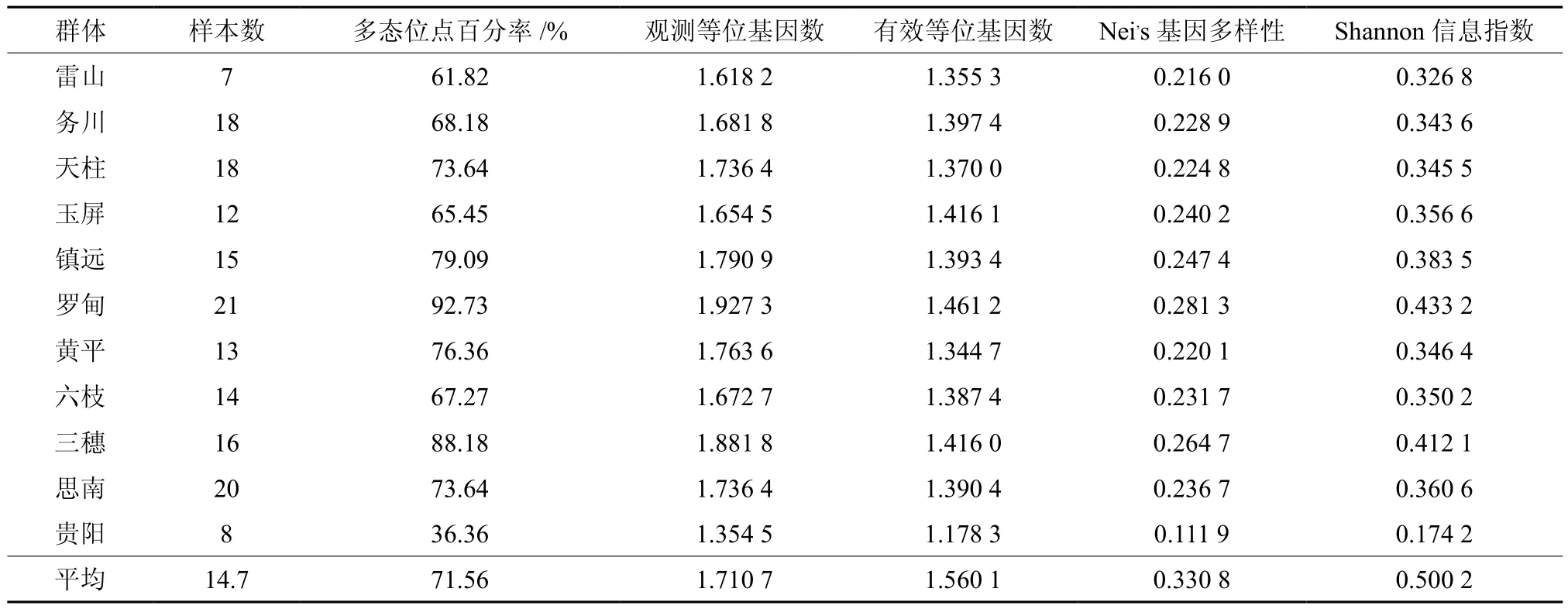
表3 贵州省马尾松天然林遗传多样性Table 3 The genetic diversity of natural groups of masson pine in Guizhou
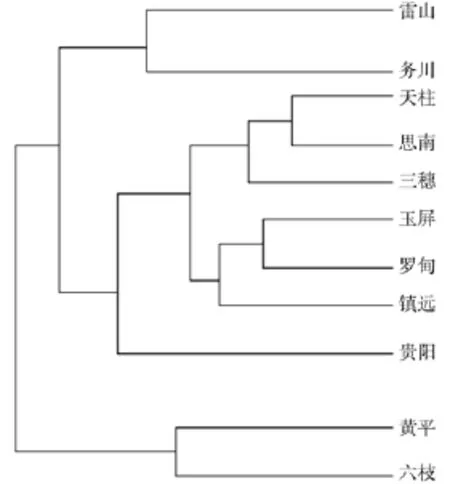
图2 马尾松天然林聚类图(UPGMA法)Fig.2 Dengrogram of eleven populations by ISSR
2.4 种子园马尾松遗传多样性分析
通过筛选的15条引物对种子园94个无性系样本进行ISSR-PCR分析,筛选的15条引物扩增的总条带数为121条,多态位点数为119条,多态位点比率为98.35%。每条引物扩增出的多态性条带数目平均为7.93条。条带片段大小在250~1500 bp之间。Nei’s基因多样度指数为0.3625,Shannon信息多样性指数为0.5326,表明建园亲本的遗传多样性水平较高,仍有进一步选择疏伐的空间。
2.5 种子园子代遗传多样性分析
通过筛选出的15条多态性ISSR引物对都匀马尾松子代测定林81个样品进行PCR扩增,共扩增出120个条带,其中多态性位点117个,多态位点百分率为97.50%,平均多态位点数为7.8条。其中观测等位基因数平均值为1.9750,有效等位基因数平均值为1.5454,Nei’s基因多样性指数均值为0.3267,Shannon信息指数均值为0.4946。表明种子园子代仍然存在较高的遗传多样性水平,人工选育过程中并没有导致遗传范围变窄。
3 讨论与小结
遗传多样性是生物界中最普遍的特征之一,遗传多样性的大小决定着物种或种群进化潜力和适应环境的能力。有效的种内遗传变异量也决定了物种在育种过程中的改良潜力。天然林资源长期进化过程中形成的庞大基因库,是遗传基础比较广泛的群体,蕴藏着巨大的生产潜力。马尾松在贵州省广泛分布,也是贵州省主要造林和用材树种,本研究利用ISSR分子标记对贵州省马尾松不同群体遗传多样性研究发现,天然林的Nei’s基因多样性指数为0.3308,Shannon信息指数为0.5002,遗传多样性略低于广西省的马尾松天然林。由于基因交流频繁,遗传多样性主要存在于种源内,在进行种质资源收集时可以少收集种源,可在遗传多样性相对较高的罗甸、三穗、玉屏等种源内多收集单株进行保存。
同广西、湖南马尾松种子园遗传多样性相比,贵州都匀马尾松种子园遗传多样性也略低,但同贵州省马尾松天然林相比,种子园遗传多样性有所升高,可能与建园材料的选择有关,早期天然林保存较好,种质资源丰富,这些天然林在地理上和遗传上有分化,由各天然林经过选择出的优树数量较多,建立的种子园便维持较高水平的遗传多样性,种子园子代也变异丰富。一般认为,在育种过程中加大人工选择强度有助于改良群体的遗传增益,同时会存在降低遗传多样性的风险,造成遗传资源选择范围变窄,严重影响林木继续改良的基础。但本研究结果与国内其它马尾松种子园研究结果再次验证了Wheeler & Jech的观点[20],即遗传学家们不用担心种子园由于群体数量有限可能导致遗传多样性较天然种群低的问题。王鹏良曾对马尾松无性系种子园连续3年的子代进行遗传多样性分析,结果发现种子园结实初期子代遗传多样性呈逐年上升趋势,随着种子园无性系的花粉量增加,子代中一些等位基因的频率会有所上升,对子代遗传多样性的贡献也逐步变大,子代遗传多样性会进一步上升,在种子园结实盛期,子代可能会有更高水平的遗传多样性[21]。本研究发现该种子园在结实初期子代遗传多样性略低于种子园亲本,现在林木郁闭,通风透光差,结实量进入衰退期,子代遗传多样性也有大幅度下降,需要结合子代测定、无性系开花结实习性等资料对种子园进行去劣疏伐,改善光照条件,同时进行辅助授粉以提高种子产量和遗传品质。
但该种子园后期营建的基因收集区由于材料来源于少数相邻县市,遗传多样性相对较低,为了长期改良需要,急需继续进行种质资源的收集、保存。
本研究小结如下:
(1)马尾松在贵州省分布广泛,也是贵州省的主要造林树种之一,马尾松天然林群体保持较高的遗传多样性。由于种源间基因交流频繁,贵州省马尾松变异主要存在于种源内。贵州省气候和生态条件的复杂多样性,使得马尾松种源间遗传多样性有了较大差异。
(2)贵州省都匀市马尾松良种基地1.5代无性系种子园亲本是在贵州省境内经过优树选择得到,来源广泛。种子园和子代仍然存在较高的遗传多样性水平,人工选育过程中并没有导致遗传范围变窄,种子园还有进一步选择的潜力。
参考文献:
[1] 崔博文,范付华,丁贵杰,等.不同地域马尾松优良无性系ISSR遗传多样性分析[J].种子,2016,5(9):41-45,50.
[2] 李元应.马尾松种质资源遗传多样性的ISSR分析[J].现代农业科技,2017(4):123-126.
[3] 王 茜,张冬林,杨模华,等.马尾松优良家系优株 ISSR 遗传距离分析[J].中南林业科技大学学报,2013,33(12):72-76.
[4] 李广军,郭彩萍,郭文娟.广西古蓬种源马尾松遗传多样性的ISSR分析[J].中南林业科技大学学报,2011,31(9):42-45.
[5] 李志辉,陈 艺,张冬林,等.广西马尾松天然林古蓬和浪水种源群体遗传多样性ISSR分析[J].中国农学通报,2009,25(16): 116-119.
[6] 冯源恒,李火根,杨章旗,等.马尾松桐棉种源天然群体遗传结构研究[J].广西植物,2016,36(11):1275-1281.
[7] 万爱华,徐有明,管兰华,等.马尾松无性系种子园遗传结构的RAPD分析[J].东北林业大学学报,2008,36(1):18-22.
[8] 龚 佳.马尾松实生种子园遗传多样性研究[D].南京:南京林业大学,2007:20-30.
[9] 张 薇,龚 佳,季孔庶.马尾松实生种子园遗传多样性分析[J].分子植物育种,2008,6(4):717-723.
[10] 陈德学.马尾松种子园无性系遗传多样性研究[D].南昌:南昌大学,2005.
[11] 杨玉洁,张冬林,杨模华,等.湖南桂阳马尾松种子园遗传多样性的ISSR分析[J].中南林业科技大学学报,2010,30(3):85-89.
[12] 朱必凤,陈德学,陈虞禄,等.广东韶关马尾松种子园遗传多样性分析[J].福建林业科技,2007,4(3):1-5.
[13] 张冬林,杨玉洁,杨模华,等.湖南城步马尾松种子园遗传多样性的ISSR研究[J].中南林业科技大学学报,2010,30(12):6-10.
[14] 艾 畅,徐立安,赖焕林,等.马尾松种子园的遗传多样性与父本分析[J].林业科学,2006,42(11):146-150.
[15] 谭小梅,周志春,金国庆,等.马尾松二代无性系种子园遗传多样性和交配系统分析[J].林业科学,2012,8(2):69-74.
[16] 杨玉洁,张冬林,杨模华,等.湖南桂阳马尾松种子园遗传多样性的ISSR分析[J].中南林业科技大学学报, 2010,30(3):85-89.
[17] 吴明长,徐嘉娟,朱亚艳,等.马尾松S R AP-PC R反应体系的建立及遗传多样性分析[J].贵州林业科技,2016,44(3):1-7.
[18] 张 一,储德裕,金国庆,等.马尾松1代育种群体遗传多样性的ISSR分析[J].林业科学研究,2009,22(6):772-778.
[19] 冯源恒,杨章旗,李火根,等.广西马尾松第一代育种群体遗传多样性[J].东北林业大学学报,2016,44(6):1-3.
[20] Wheeler N C, Jech K S. The use of electrophotic markers in seed orchard research[J]. New Forestry,1992,6:311-328.
[21] 王鹏良.马尾松无性系种子园多年份子代遗传多样性分析[D].南京:南京林业大学,2006.
猜你喜欢
现代妇产科进展(2022年1期)2022-12-07
中国生育健康杂志(2022年4期)2022-11-25
浙江林业科技(2022年4期)2022-07-16
绿色科技(2021年21期)2021-11-26
辽宁林业科技(2021年3期)2021-07-08
南方林业科学(2021年2期)2021-05-27
辽宁林业科技(2021年1期)2021-03-16
山西水土保持科技(2020年2期)2020-12-22
农家科技中旬版(2020年4期)2020-07-06
农业与技术(2016年22期)2017-03-07
