
灰胡杨光合生理指标与微气象环境因子的相关分析
2018-04-12 06:49施英俊
新疆农垦科技 2018年2期
施英俊
(兵团林业工作管理总站,新疆 乌鲁木齐 830011)

灰胡杨(Populus pruinosa Schrenk)是杨柳科杨属的植物,作为极端干旱区荒漠河岸林中仅有的较为古老和原始的树种之一,广泛分布于塔里木河流域,为我国首批确定的388种珍稀濒危植物中的渐危种。灰胡杨因长期生长在干旱的沙漠周边河流沿岸,因此在生理和生态功能上具备了耐干旱、耐盐碱、抗风沙等优良特性。正是由于其突出的生态适应性,目前灰胡杨作为天然林和人工林的建群种,在绿洲防风固沙、调节生态平衡、维持区域生物多样性和保障绿洲农牧业生产安全方面发挥了不可替代的作用。但是由于其生长区域内大规模的水土资源开发利用,从20世纪50年以后,一些区域河流水量减少,沿岸地下水位下降较快,导致了河流沿岸,及周边区域天然胡杨、灰叶胡杨林大面积退化死亡,其生态作用也急剧下降。因此,人们开始逐渐重新审视胡杨对区域环境的生态和经济价值,并开展了一系列的研究,其中一项重要工作就是关于胡杨生理生态学特性方面的研究。但在荒漠地区面积最大的新疆,胡杨、灰叶胡杨光合生理指标与微气象环境因子关系的研究报道还较少,也缺少系统研究。本研究通过对灰胡杨光合生理指标与环境因子的关系研究,分析其适应极端环境的生理生态特性及适应机制,为幼苗期间的抚育管理和更新复壮,以及进一步选择适宜的天然林保护和恢复技术措施提供科学的理论依据。
1 材料与方法
1.1 研究区概况
研究区位于巴音郭勒蒙古族自治州轮南镇(E84.25°、N41.15°),平均海拔高度 917 m,属暖温带大陆性干旱气候,夏季炎热,冬季寒冷,年日照时数为2 684 h左右,年平均气温为10.9℃,极端最低气温为-25.5℃,年平均日较差为14.6℃,无霜期202 d左右。年平均降水量为65.5 mm,年平均蒸发量为2 024 mm,平均风速1.8 m/s[1]。区域内胡杨林自然保护区是国内面积最大和保存较为完整的原始胡杨林之一。胡杨群落在不同时期变迁的河道周边分布,总体上各个群落组成结构简单、物种多样性低。乔木层主要有胡杨(Populus euphratica Oliv)和灰胡杨 (Populus pruinosa Schrenk)。林下灌木层以柽柳(Tamarix spp.)、铃铛刺(Halimodendron halodendron)等荒漠成分种类组成,草本层主要由骆驼刺(Alhagi sparsifolia)、 芦 苇 (Phragmites communis)、 甘 草(Glycyrrhiza inflata)等荒漠化草甸或盐渍化草甸的多年生植物种类组成。研究样地地下水位3~4 m,林分结构组成简单,林层分化不明显,林下植物种类较少,其中乔木层郁闭度0.45左右,灌木层盖度0.92%[2],土壤质地为砂壤土,pH平均为8.6左右,林分整体属于成熟林阶段。
1.2 材料的选取
研究区天然胡杨林内仅在地下水位3.5 m处有一块灰胡杨林,呈带状分布。2016年6月上旬在灰胡杨林内选择代表性的林分分别设置30 m×30 m的样方一块,在样方内每木捡尺,按径级选取8~10株生长健康无病害的灰胡杨作为实验材料。
1.3 测定项目和方法
光合生理参数测定:2016年6月和8月中旬,在晴朗无云的天气连续测定3 d,然后选择气象和数据采集状况最好的一天分析。测定方法为利用便携式光合作用测定仪(Li-6400P和Li-cor USA)测定灰胡杨叶的光合生理参数的日变化,在每个样方选择待测样本树冠中上部外围向阳新梢上的3、4位成熟叶,于当地时间08:00~20:00每小时测定1次,每株测定3~5个叶片。测定指标包括:(1)生理指标:净光合速率(Pn)、胞间 CO2浓度(Ci)、蒸腾速率(Tr)和气孔导度(Gs);(2)微气象参数:植物所测部位光合有效辐射强度(PAR)、大气温度(Ta)、空气相对湿度(RH)、叶气饱和水蒸汽压差(VPD)等[3]。
1.4 实验处理
试验数据采用Excel、DPS软件进行处理和分析。
2 结果与分析
2.1 灰胡杨光合生理指标的日动态变化
2.1.1 灰胡杨净光合速率的日变化
灰胡杨6月和8月的净光合速率日变化都呈现出双峰曲线M字形[4],灰胡杨净光合速率日动态变化趋势虽然大体相同,但是6月份的双峰曲线比8月份的明显。由于6月和8月光照强度和温度的不同造成了灰胡杨曲线峰值和谷值出现的时间不同。6月的2个峰值出现在12:00和16:00,大小分别为19.55 μmol·m-2·s-1和 14.66 μmol·m-2·s-1, 谷值出现在 15:00,大小为 6.97 μmol·m-2·s-1。 8 月份的 2个峰值出现在 13:00和 15:00,其大小分别为15.06 μmol·m-2·s-1和 14.92 μmol·m-2·s-1,谷值出现在14:00,大小为 12.22 μmol·m-2·s-1。 6 月的最大峰值比8月的大,而且2个月的双峰曲线的第2个峰值几乎相等。6月的净光合速率值在10:00~14:00以及16:00~20:00都比8月的大。
2.1.2 灰胡杨气孔导度的日变化
气孔导度是反映植物气孔行为的重要生理指标,它的开闭会对植物的光合作用和蒸腾作用产生一定的影响[5]。6月的气孔导度值均比8月小,8月的气孔导度日变化的变化幅度比6月大,6月和8月气孔导度日变化总势大致相同,但8月的气孔导度变化起伏不定呈现多峰,而6月的气孔导度变化相对平缓呈现单峰,8月有3个峰值,分别出现在10:00、15:00 和 19:00, 这 3 个峰值是依次降低的,大小为 0.998 mol·m-2·s-1、0.469 mol·m-2·s-1和 0.421 mol·m-2·s-1,有 2 个谷值出现在 14:00 和 16:00,大小为0.383 mol·m-2·s-1和 0.251 mol·m-2·s-1。6 月份仅有 1个峰值出现在 10:00, 大小为 0.525 mol·m-2·s-1,有1 个谷值出现在 17:00,大小为 0.142 mol·m-2·s-1。6月和8月的气孔导度在上午迅速上升,在10:00都达到最大值,其后随时间推移植物的气孔导度呈逐渐降低趋势,直至17:00以后才逐渐有所回升,这表明灰胡杨会通过降低气孔导度来适应中午的光强和高温。
2.1.3 灰胡杨胞间CO2浓度日变化
灰胡杨8月份的胞间CO2浓度日变化幅度比6月份小,二者变化总趋势大致相同。8月份的胞间CO2浓度日变化呈扁平的V字形,6月份的胞间CO2浓度日变化呈扁平的W字形,8月份的胞间CO2浓度值均比6月的大。8月份的胞间CO2浓度日变化无峰值,仅有1个谷值在16:00,大小为328 μmol·mol-1。 6 月份的胞间 CO2浓度日变化有 2个谷值和1个峰值,分别出现在13:00、17:00和15:00,大小分别为 241 μmol·mol-1、187 mol·mol-1和295 μmol·mol-1。 6 月和 8 月分别是从 08:00 的最大值 347 μmol·mol-1和 436 μmol·mol-1开始下降,都呈现出上午由于气孔导度和净光合速率的增大而下降,下午由于气孔导度和净光合速率的减小而上升的趋势[6]。
2.1.4 灰胡杨蒸腾速率的日变化
灰胡杨6月和8月的蒸腾速率日变化的总趋势大致相同,二者从08:00由于温度上升以及气孔导度的增大促进了蒸腾速率的迅速上升并达到最大值,中午则为了适应中午的光强和高温蒸腾速率而下降[7]。但是二者到达最大值的时间点不同,6月是在12:00到达最大值,8月是在11:00到达最大值。二者到达最大值以后都呈现出起伏不定的变化,6月份呈现双峰而8月份呈现多峰,但是总的趋势都是下降的。6月份共有2个峰值,出现在12:00和16:00,大小分别为 22.55 mmol·m-2·s-1和 17.57 mmol·m-2·s-1,共有 2个谷值,出现在14:00和19:00,大小分别为16.74 mmol·m-2·s-1和 8.32 mmol·m-2·s-1。 8 月有 5 个峰值,出现在 11:00、13:00、15:00、17:00 和 19:00,大小分别为 21.52 mmol·m-2·s-1、21.30 mmol·m-2·s-1、20.23 mmol·m-2·s-1、16.09 mmol·m-2·s-1和 15.88 mmol·m-2·s-1, 共有 4 个谷值出现在 12:00、14:00、16:00 和 18:00,其大小分别为 19.06 mmol·m-2·s-1、16.65 mmol·m-2·s-1、12.66 mmol·m-2·s-1和 15.29 mmol·m-2·s-1。
2.2 光合生理指标与微气象环境因子的关系
2.2.1 光合生理指标与微气象环境因子的相关分析
很多生态因子会影响灰胡杨的光合生理指标变化,而这些变化是内外因子综合作用的结果。而采用测定光合生理参数与环境因子数据进行相关性分析,探究微气象环境因子对灰胡杨光合生理指标的影响,可以揭示灰胡杨光合生理指标与其微气象因子之间的关系[8]。
2.2.1.1 6月灰胡杨光合生理指标与微气象环境因子的相关分析
6月灰胡杨光合生理指标与微气象环境因子间的相关系数及其显著性如表1,净光合速率Pn与光合有效辐射强度PAR呈极显著相关关系,相关系数为0.89,除此之外与其他微气象环境因子相关都不显著。气孔导度Gs与上述微气象环境因子相关都不显著。胞间CO2浓度Ci与叶气饱和蒸气压差VPD、温度Ta、空气相对湿度RH呈极显著相关,相关系数为-0.89、-0.81和0.81。蒸腾速率Tr与温度Ta、空气相对湿度RH呈显著相关关系,与光合有效辐射强度PAR呈极显著相关关系[9],相关系数为0.66、-0.59和0.77。由此可知,6月份光合有效辐射强度PAR是影响灰胡杨净光合速率Pn的唯一微气象环境因子,微气象环境因子对气孔导度GS的影响不显著。影响胞间CO2浓度Ci的微气象环境因子的大小顺序为:VPD>RH=Ta。影响蒸腾速率的微气象环境因子的大小顺序为:PAR>Ta>RH。
2.2.1.2 8月灰胡杨光合生理指标与微气象环境因子的相关分析

表1 6月灰胡杨光合生理参数与微气象环境因子的相关分析
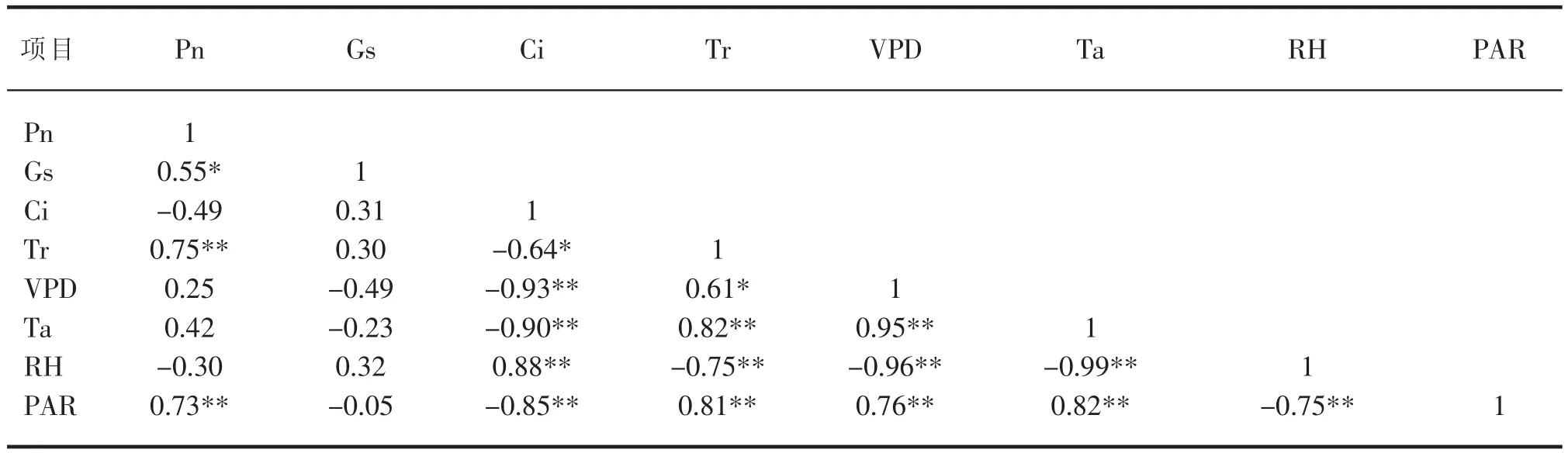
表2 8月灰胡杨光合生理参数与微气象环境因子的相关分析
8月的光合生理指标与微气象环境因子的相关系数及其显著性如表2,净光合速率Pn与光合有效辐射强度PAR呈极显著相关关系,相关系数为0.73。气孔导度Gs与上述微气象环境因子相关都不显著。胞间CO2浓度Ci与温度Ta、叶气饱和蒸气压差VPD、空气相对湿度RH、光合有效辐射强度PAR都呈极显著相关关系,相关系数为-0.90、-0.93、0.88和-0.85。蒸腾速率Tr与叶气饱和蒸气压差VPD显著相关,与温度Ta、空气相对湿度RH、光合有效辐射强度PAR呈极显著相关关系。由此可知,8月份灰胡杨影响净光合速率Pn的微气象环境因子主要是光合有效辐射强度PAR。微气象环境因子对气孔导度GS的影响不显著。胞间CO2浓度Ci与叶气饱和蒸气压差VPD、温度Ta、空气相对湿度RH、光合有效辐射强度PAR都呈极显著相关关系,相关系数为-0.93、-0.90、0.88和-0.85。蒸腾速率 Tr与叶气饱和蒸气压差VPD呈显著相关关系,与空气相对湿度RH、温度Ta、光合有效辐射强度PAR都呈极显著相关关系,相关系数为0.82、0.61、-0.75和0.81。由此可知,8月份影响灰胡杨净光合速率Pn的微气象环境因子主要是光合有效辐射强度PAR,微气象环境因子对气孔导度GS的影响不显著,影响胞间CO2浓度Ci的微气象环境因子的大小顺序为:VPD>Ta>RH>PAR。影响蒸腾速率的微气象环境因子的大小顺序为:Ta>PAR>RH>VPD。

表3 6月灰胡杨光合生理参数与微气象环境因子的偏相关分析
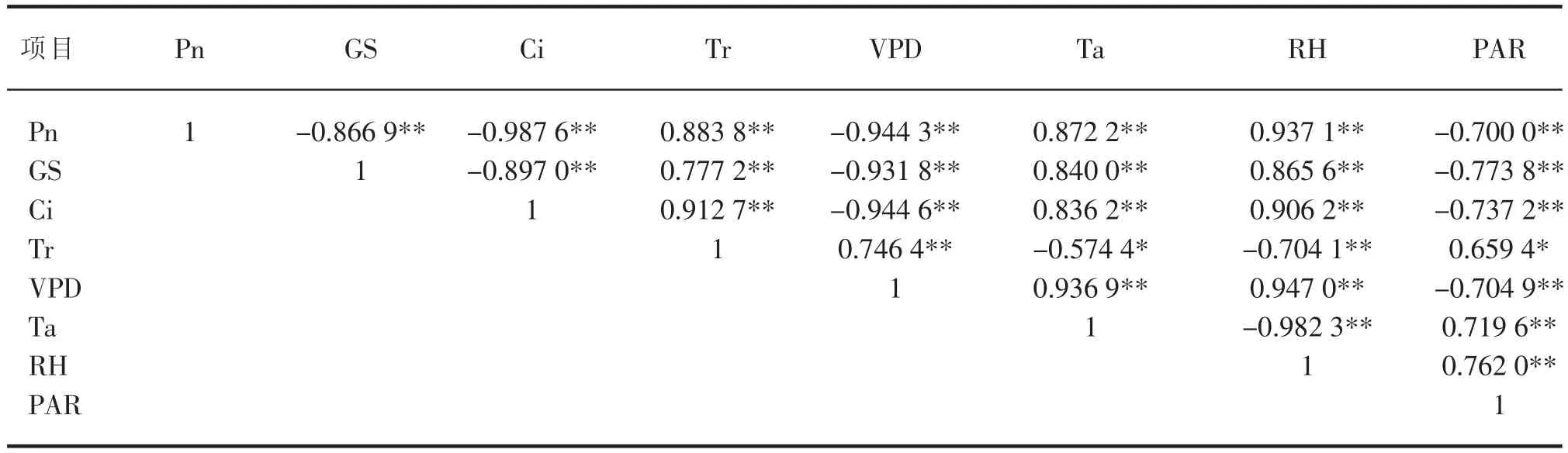
表4 8月灰胡杨的光合生理指标与微气象环境因子的偏相关分析
2.2.2 光合生理指标与微气象环境因子的偏相关分析
2.2.2.1 6月灰胡杨光合生理指标与微气象环境因子的偏相关分析
偏相关分析就是在研究2个变量之间的线性相关关系时控制可能对其产生影响的其他变量[9]。6月份灰胡杨的光合生理指标与微气象环境因子的偏相关系数及其显著性如表3,净光合速率与微气象环境因子的偏相关不显著。气孔导度与叶气饱和水蒸汽压差偏相关极显著,与空气相对湿度、光合有效辐射强度、温度偏相关显著,而胞间CO2浓度与微气象环境因子偏相关不显著。蒸腾速率仅与温度偏相关显著。由此可知,6月的微气象环境因子对灰胡杨的净光合速率和胞间CO2浓度的影响不明显,影响气孔导度的微气象环境因子的大小顺序为:VPD>PAR> RH>Ta。
2.2.2.2 8月灰胡杨光合生理指标与微气象环境因子的偏相关分析
8月份灰胡杨的光合生理指标与微气象环境因子的偏相关系数及其显著性如表4。净光合速率、气孔导度、胞间CO2浓度与净光合速率、气孔导度、蒸腾速率、胞间CO2浓度都呈偏向关极显著关系。蒸腾速率与叶气饱和蒸气压差、空气相对湿度呈偏相关极显著关系,与温度、光合有效辐射强度呈偏相关显著关系。由此偏相关系数大小及其显著性可知,8月份影响灰胡杨的净光合速率、气孔导度、胞间CO2浓度的微气象环境因子大小顺序都为:VPD>RH>Ta>PAR,影响蒸腾速率的微气象因子大小顺序为:VPD>RH>PAR>Ta。
2.2.3 主要光合生理指标净光合速率、气孔导度、蒸腾速率与微气象环境因子的多元逐步回归分析
简单的相关分析只能反映2个因素间表面而非本质的联系。而逐步多元回归分析方法能够有效地从众多影响因素中挑选出对各因子y的变化贡献较大的因子,并建立Pn、Gs、Tr与这些因子的最优回归方程[11]。选取叶气饱和蒸气压差VPD、温度Ta、空气相对湿度RH、光合有效辐射强度PAR这8个因子与各生理参数y进行多元逐步回归分析,得到逐步回归最优方程(见表5)。线性回归方程的R2一般要大于0.75才说明此拟合方程可信[9],经验证,表5中各方程的R2都大于0.75,各方程经显著性检验,相关系数显著性均小于0.01,表明灰胡杨各生理参数与微气象环境因子间的线性关系成立。

表5 主要光合生理参数与微气象环境因子的最优回归方程
根据表5最优回归方程各因子前系数的大小可得出各微气象环境因子对各光合生理参数影响的大小顺序。6月影响Pn的主要微气象环境因子的大小顺序为:VPD>RH>PAR,影响Gs的主要微气象环境因子的大小顺序为:VPD>Ta>RH>PAR,影响Tr的主要微气象环境因子的大小顺序为:Ta>RH>VPD。8月影响Pn的主要微气象环境因子的大小顺序为:RH>Ta>PAR,影响Gs的主要微气象环境因子的大小顺序为:VPD>RH>PAR,影响Tr的主要微气象环境因子的大小顺序为:RH>Ta>PAR。
3 讨论
受外界和内在因子的影响,灰胡杨的光合生理指标对环境的变化,尤其是对微气象环境因子的变化十分敏感。本研究区6月和8月灰胡杨Pn和6月的Tr的日变化呈双峰曲线,且出现了光合午休现象。从6月微气象环境因子与灰胡杨的Pn以及Tr的相关性分析结果可得,决定灰叶胡杨Pn日变化的主要微气象环境因子是PAR,决定灰叶胡杨Tr的日变化的微气象环境因子的大小顺序为:PAR>Ta>RH,这与前人对塔里木河中游灰胡杨研究结果一致[11]。
通过对灰胡杨6月和8月的光合生理指标与微气象环境因子的相关分析及其显著性知,PAR、Ta、RH是影响灰胡杨光合生理指标的主要微气象环境因子,它们通过直接和间接的交互作用,对灰胡杨的光合生理指标产生综合影响。这与前人对陕西省永寿县马莲滩流域的刺槐的研究结果相同[4]。在实际生存过程中,微气象环境因子之间相互联系,经常以某个因子为主进而影响其他因子,例如:PAR直接影响Ta和RH的变化,因此,无论是对Pn还是对Tr,PAR都是最主要的主导因子,其次是Ta和RH[11]。
一般认为Ta往往与PAR结合在一起共同决定叶气饱和水蒸气压差[6],灰胡杨6月和8月的Ta与VPD的相关极显著而且偏相关也极显著,说明了Ta对VPD的确有影响作用。由Gs对各微气象环境因子的最优回归方程知,VPD是对因变量Gs有直接影响的因子。所以Ta通过影响VPD间接的影响了Gs的变化。
由6月和8月Tr与RH相关分析和偏相关分析可知,RH显著影响植物的蒸腾速率Tr,即影响叶片失水和气孔涨缩,而气孔是CO2的进口,也是蒸腾过程中水汽主要的出口,因此,RH对Tr和Gs有着直接影响和间接影响的作用[6]。
4 结论
(1)6月和8月的净光合速率、气孔导度、胞间CO2浓度、蒸腾速率的日变化总趋势相同;(2)由相关分析得出,光合有效辐射强度PAR是影响净光合速率Pn的主要微气象环境因子;(3)由偏相关分析和逐步回归分析得出,影响气孔导度Gs的主要微气象环境因子是叶气饱和水蒸气压差VPD;(4)由相关分析和偏相关分析得出,影响胞间CO2浓度的主要微气象因子是叶气饱和水蒸气压差VPD、空气相对湿度RH和温度Ta;(5)通过相关分析、偏相关分析和逐步回归分析得出,叶气饱和水蒸气压差VPD、空气相对湿度RH、温度Ta是影响影响蒸腾速率Tr的主要微气象环境因子。
[1]韩路,王海珍,彭杰,等.塔里木荒漠河岸林植物群落演替下的土壤理化性质研究[J].生态环境学报,2010,19(12):2808-2814.
[2]韩路,王海珍,陈加利,等.塔里木荒漠河岸林干扰状况与林隙特征[J].生态学报,2011,31(16):4699-4708.
[3]周洪华,陈亚宁,李卫红,等.塔里木河下游胡杨气体交换特性及其环境解释[J].中国沙漠,2008,28(4):665-672.
[4]石贵玉,李明霞,梁士楚,等.桉树净光合速率日变化的季节动态研究[J].广西师范大学学报(自然科学版),2013,31(2):113-117.
[5]王曼.克里雅河流域水盐梯度下三种荒漠植物的生态响应研究[D].新疆大学,2014.
[6]郑元,赵忠,周靖靖,等.刺槐光合气体交换与环境因子的关系[J].西北农林科技大学学报(自然科学版),2011,39(4):81-88.
[7]杨泽粟.黄土高原植被生理过程和蒸散量计算方法及变化特征研究[D].兰州大学,2016.
[8]袁月.艾比湖流域胡杨光合与水分生理特征研究[D].新疆大学,2009.
[9]王曼.克里雅河流域水盐梯度下三种荒漠植物的生态响应研究[D].新疆大学,2014.
[10]周宝良,朱协飞,郭旺珍,等.棉花高品质种质系杂交后代纤维品质性状间的偏相关分析[J].棉花学报,2006,18(6):352-356.
[11]张志刚,李宏,DENNY WALTHER,等.塔里木河中游胡杨与灰叶胡杨气体交换特性对比研究[J].西北植物学报,2012,32(12):2506-2511.
猜你喜欢
农业工程学报(2022年3期)2022-04-16
西南农业学报(2021年7期)2021-08-23
节水灌溉(2021年5期)2021-05-26
生态学报(2021年6期)2021-04-25
农业工程学报(2021年3期)2021-04-15
节水灌溉(2020年9期)2020-09-16
农业与技术(2020年12期)2020-07-04
中国农业科技导报(2020年12期)2020-03-15
农学学报(2019年1期)2019-02-21
新课程·下旬(2017年7期)2017-08-14
