
Ku70基因稳定敲除HeLa细胞株的建立及其生物学功能研究
2018-04-09 03:37李达沈雪莲李少华丁红梅李慧黄皑雪耿介王超男白琛俊张坦董洁邵宁生
生物技术通讯 2018年2期
李达,沈雪莲,李少华,丁红梅,李慧,黄皑雪,耿介,王超男,白琛俊,张坦,董洁,邵宁生
军事医学研究院 军事认知与脑科学研究所,北京 100850
Ku蛋白是一种含量丰富的核蛋白,进化上高度保守,分布广泛,从细菌到人类均有表达[1-2]。人Ku蛋白是异二聚体,由Ku70和Ku80两个亚基组成。研究发现,Ku蛋白具有不同寻常的DNA结合特性,其以非序列依赖性的方式紧密结合在双链DNA的断端,是经典非同源末端连接(classi⁃cal non-homologous end joining,C-NHEJ)途径中的DNA末端结合因子,招募DNA依赖的蛋白激酶催化亚基(DNA-PKcs)与其结合形成DNA依赖的蛋白激酶全酶,催化双链断裂DNA的修复[2-7]。
此外,Ku蛋白还可能参与其他重要的生物学过程,比如Ku蛋白被证实与染色体端粒结构的维持有密切联系。研究发现在失去任意一个Ku蛋白亚基后,酿酒酵母就会出现端粒结构缺失的现象[8-9]。还有研究显示Ku70与细胞凋亡有直接联系,即Ku70能够与促凋亡因子Bax结合,抑制其促进细胞凋亡的作用[10]。然而,Ku蛋白的生物学功能尚未得到完全阐释,有待深入研究。
我们利用CRISPR/Cas9基因编辑技术破坏HeLa细胞Ku70基因开放读框,同时在其中插入潮霉素B抗性基因,然后通过潮霉素B抗性筛选获得Ku70基因稳定敲除的HeLa细胞株,再进一步利用此细胞株研究Ku70蛋白的生物学功能。
1 材料与方法
1.1 材料
HeLa细胞由本实验室保存;pCas-guide和pSilencer2.1-U6hygro质粒由军事医学研究院郑晓飞教授惠赠;感受态大肠杆菌DH5α、T4DNA连接酶、细胞基因组DNA提取试剂盒、琼脂糖凝胶DNA回收试剂盒、质粒小提试剂盒购自康为世纪生物科技有限公司;PCR平末端产物加A试剂盒、pBackZero-T载体为TaKaRa公司产品;pfuDNA聚合酶购自北京全式金生物技术公司;限制性核酸内切酶BamHⅠ、BsmBⅠ为Thermo Scientific公司产品;PRIME jet转染试剂为PolyPlus公司产品;潮霉素B为Roche公司产品;Ku70抗体购自Pro⁃teintech公司;GAPDH抗体购自中杉金桥公司;CCK-8试剂购自上海碧云天生物技术有限公司;2×SYBR Green Mix购自Toyobo公司;引物由生工生物技术公司合成。
1.2 构建pCas-gRNA质粒
用CRISPR DESIGN程序设计针对人Ku70基因的向导RNA(guide RNA,gRNA)序列(表1),合成编码gRNA的DNA序列,退火形成双链DNA。用限制性内切酶BamHⅠ、BsmBⅠ对pCas-guide载体和退火产物进行双酶切,胶回收酶切产物。将酶切后的pCas-guide载体和退火产物用T4DNA连接酶于20℃连接4 h,连接产物转化大肠杆菌感受态细胞DH5α,涂布于氨苄西林(Amp)抗性的细菌培养板上,37℃培养过夜。挑取细菌单克隆,菌液PCR鉴定后选取阳性克隆测序。

表1 合成的引物序列
1.3 构建供体DNA质粒pBackZero-T-Ku70
以HeLa细胞基因组为模板,Ku70-F1/Ku70-R1为引物扩增Ku70左同源臂片段Ku70-L,以Ku70-F2/Ku70-R2为引物扩增Ku70右同源臂片段Ku70-R;以pSilencer2.1-U6hygro质粒为模板,Hy-F/Hy-R为引物扩增潮霉素B抗性基因片段Hy;琼脂糖凝胶回收试剂盒分别回收片段Ku70-L、Ku70-R和Hy。以片段Ku70-L、Hy为模板,Ku70-F1/Hy-R为引物,重叠PCR扩增出Ku70-LHy;再以Ku70-L-Hy和Ku70-R为模板,Ku70-F1/ Ku70-R2为引物,扩增获得片段Ku70-KO,回收目的片段。用A-Tailing Kit在Ku70-KO片段平末端添加单个碱基A后,将该片段连接到pBackZero-T载体,转化感受态大肠杆菌DH5α并挑取单克隆菌株进行菌液PCR鉴定,选取阳性克隆测序。
1.4 细胞转染、抗性筛选与鉴定
将HeLa细胞以3×105/mL的密度接种于6孔细胞培养板中,细胞生长至30%~40%时转染0.8 μg pCas-gRNA质粒和1.2 μg pBackZero-T-Ku70质粒,转染后72 h收集细胞,以4×104/mL的密度接种至6孔板中,待细胞完全贴壁后加入潮霉素B(200 μg/mL),2~3 d更换一次含同浓度潮霉素B的新鲜培养基,培养至第7 d提取细胞基因组做PCR鉴定。以提取的细胞基因组为模板,Ku70左同源臂基因上的引物K-F和潮霉素B抗性基因上的引物K-R进行PCR扩增。之后将细胞扩大培养,冻存。
1.5 单克隆稳定细胞株筛选与鉴定
用有限稀释法筛选Ku70稳定敲除的HeLa单克隆细胞株,胰酶消化细胞,以新鲜培养基稀释细胞浓度至10/mL,将稀释的细胞悬液接种至96孔板,100 μL/孔,培养1周后显微镜下观察,选取含单克隆细胞的孔进行扩大培养。扩大培养至24孔板时改用含200 μg/mL潮霉素B的培养基继续培养,1周后收集细胞进行Western印迹鉴定。
1.6 Western免疫印迹实验
收集细胞,用RIPA提取细胞总蛋白,BCA蛋白定量,行12%SDS-PAGE,每泳道35 μg蛋白,然后湿转至PVDF膜,用5%脱脂奶粉室温封闭1 h;一抗室温孵育2 h,0.1%TBS-T洗3次;二抗室温孵育1 h,0.1%TBS-T洗3次;ECL曝光显影。
1.7 CCK-8法测定Ku70稳定敲除细胞株的增殖能力
将处于对数生长期的Ku70稳定敲除细胞和野生型HeLa细胞以3×104/mL的密度接种于96孔细胞培养板,每孔100 μL,分别在0、24、48、72、96 h加入CCK8试剂,每孔10 μL,继续培养1 h后检测D450nm值,制作细胞的96 h生长曲线。
1.8 Transwell方法检测Ku70稳定敲除细胞株的迁移能力
将处于对数生长期的Ku70稳定敲除细胞和野生型HeLa细胞以2×105/mL的密度种于Tran⁃swell小室的上室,下室加入500 μL完全培养基,培养12 h后擦去未穿膜的细胞,用4%多聚甲醛固定穿膜细胞,0.1%结晶紫染色后在显微镜下观察并拍照。
1.9 Ku70稳定敲除细胞的miRNA表达水平检测
用TRIzol法提取Ku70稳定敲除细胞和野生型HeLa细胞的总RNA,取1 μg RNA反转录为cDNA。qPCR反应体系包括1 μL cDNA、1 μL引物(表1)、10 μL 2×qPCR SYBR Green mix、8 μL ddH2O。反应条件:预变性95℃ 10 min,变性95℃ 20 s,退火55℃ 20 s,延伸72℃ 20 s,50个循环,采集熔解曲线。用 StrataGene公司的Mx3000P qPCR仪检测,Mx3000P软件分析结果。
1.10 统计分析
所有实验均重复3次以上,数据以x±s表示,统计分析使用SAS 9.2软件的Student'sttest,P<0.05视为具有统计学差异。
2 结果
2.1 设计并构建pCas-gRNA质粒
用 CRISPR DESIGN程 序(http://crispr.mit. edu/)设计gRNA序列。选取Ku70编码区100~300 bp序列输入对话框,从输出的结果中选择合适的靶位点序列。为了防止脱靶,我们共设计了3条gRNA序列。将合成的gRNA互补链退火形成双链,与双酶切的线性化pCas-guide连接,获得重组质粒pCas-gRNA。转化铺板后挑取单克隆菌落进行PCR鉴定,将阳性克隆测序(图1),结果显示3条gRNA序列均正确插入pCas-guide载体,pCasgRNA质粒构建成功。
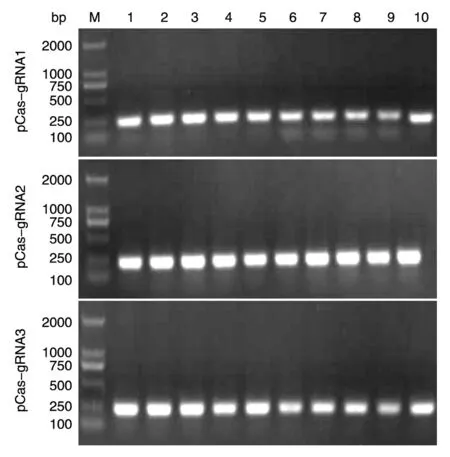
图1 pCas-gRNA质粒菌液PCR鉴定M:DNA marker;1~10:单克隆
2.2 构建供体DNA质粒pBackZero-T-Ku70
以HeLa细胞基因组为模板,分别以Ku70-F1/ Ku70-R1、Ku70-F2/Ku70-R2为引物,扩增长度分别为500和540 bp的Ku70左右同源臂Ku70-L、Ku70-R;以质粒pSilence2.1-U6 hygro为模板,Hy-F/Hy-R为引物,扩增长度为1026 bp的潮霉素抗性基因片段Hy;进而,以DNA片段Ku70-L、Hy为模板,Ku70-F1和Hy-R为引物,利用搭桥PCR扩增片段Ku70-L-Hy;再以Ku70-L-Hy和Ku70-R为模板,Ku70-F1和Ku70-R2为引物,扩增获得2066 bp的Ku70同源臂供体DNA片段Ku70-KO。琼脂糖凝胶切胶回收PCR产物中的目的片段,在其末端添加单个碱基A,再将该片段连接到pBackZero-T载体上,菌液PCR鉴定重组菌,将阳性克隆测序(图2),序列比对结果表明重组质粒序列正确,同源臂供体DNA载体pBackZero-TKu70构建成功。
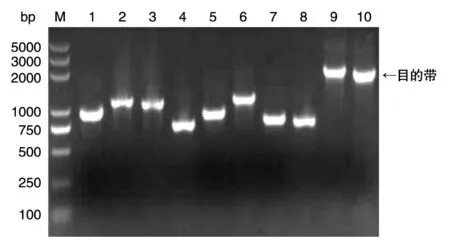
图2 pBackZero-T-Ku70质粒菌液PCR鉴定M:DNA marker;1~10:单克隆
2.3 筛选鉴定Ku70敲除的多克隆细胞株
将重组质粒pBackZero-T-Ku70分别与3种pCas-gRNA质粒共转染HeLa细胞,72 h后加入终浓度为200 μg/mL的潮霉素进行抗性筛选,15 d后收集细胞,提取细胞总蛋白,Western印迹检测Ku70蛋白的表达水平。结果显示,与对照细胞相比,转染质粒pCas-gRNA3的细胞Ku70条带明显减弱,提示gRNA3敲除效果最佳(图3)。
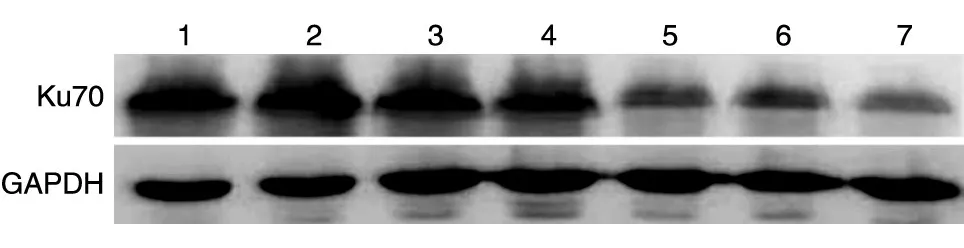
图3 Western印迹检测多克隆细胞株中Ku70的表达1:未转染的对照细胞;2:pBackZero-T-Ku70载体和pCas-gRNA1载体共转染的细胞;3:pBackZero-T-Ku70载体和pCas-gRNA2载体共转染的细胞;4~7:pBackZero-T-Ku70载体和pCas-gRNA3载体共转染的细胞
2.4 Ku70敲除细胞的基因组PCR鉴定
提取细胞基因组DNA,以K-F/K-R为引物进行PCR鉴定,结果见图4,共转染载体pBackZero-T-Ku70和pCas-gRNA3的细胞基因组DNA,经PCR扩增后在800 bp左右出现目的条带,而未转染的对照细胞没有此条带,表明潮霉素B抗性基因正确插入Ku70基因的特定位点。
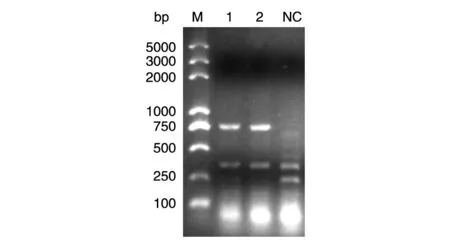
图4 基因组PCR鉴定敲除Ku70基因的单克隆细胞株1,2:共转染pBackZero-T-Ku70和pCas-gRNA3的细胞;NC:未转染的细胞
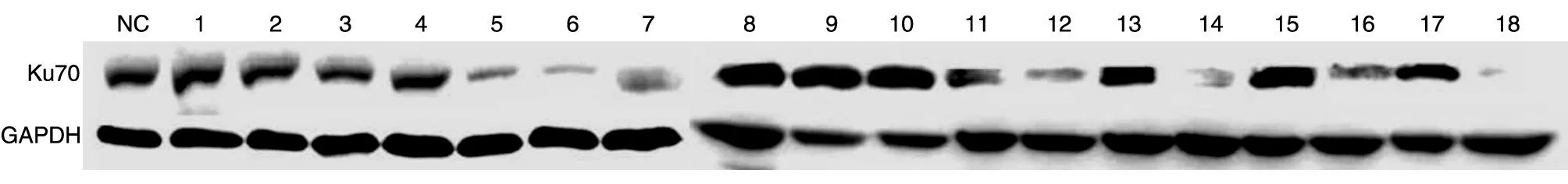
图5 敲除Ku70基因的单克隆细胞株Western印迹鉴定NC:野生型HeLa细胞;1~18:单克隆HeLa细胞株
2.5 分离并鉴定Ku70稳定敲除的单克隆细胞株
用有限稀释法筛选Ku70稳定敲除的HeLa单克隆细胞株。将细胞接种至96孔板,培养1周后选取单克隆细胞进行扩大培养,继而收集细胞,提取细胞提取总蛋白进行Western印迹鉴定(图5)。本次实验共获得18株细胞,其中14号和18号2株为敲除Ku70的单克隆细胞株。
2.6 敲除Ku70基因的稳定细胞株生物学功能检测
2.6.1 细胞增殖实验 用CCK-8法检测敲除Ku70基因的18号细胞株的增殖能力,得到18号Ku70稳定敲除细胞(KO)和野生型HeLa细胞(WT)的生长曲线(图6),与野生型比较,敲除Ku70基因的HeLa细胞增殖能力明显减弱。
2.6.2 细胞迁移实验 用Transwell方法检测18号细胞株的迁移能力,显微镜下观察并拍照(图7A);分别统计实验组和对照组穿膜的HeLa细胞数,并进行统计学分析(图7B)。实验结果表明,与野生型细胞相比,敲除Ku70基因的HeLa细胞迁移能力明显减弱。
2.6.3 实时荧光定量PCR分析敲除Ku70基因后HeLa细胞5种miRNA的表达水平 用TRIzol法提取HeLa细胞总RNA,反转录成cDNA,用qPCR仪检测5种miRNA的表达水平。结果见图8,与WT比较,敲除Ku70基因后hsa-miR-649、hsa-miR-562、hsa-miR-544a的表达水平显著上调,hsamiR-548a、hsa-miR-492水平没有显著变化。
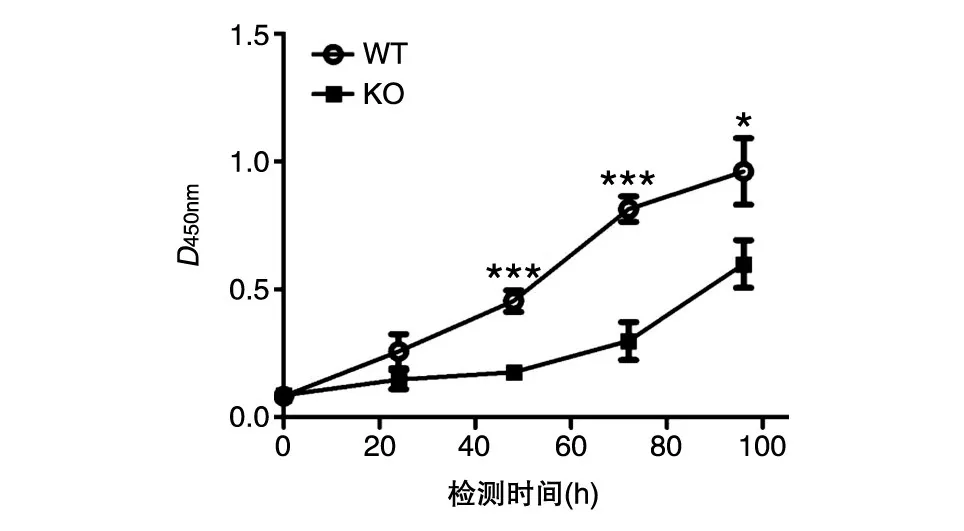
图6 Ku70基因稳定敲除细胞株生长曲线(n=3)WT:野生型HeLa细胞;KO:Ku70稳定敲除细胞;*P<0.05,***P<0.001
3 讨论
作为C-NHEJ途径中的重要因子,Ku蛋白的DNA断裂修复功能已经有了较为详尽的研究报道。与DNA断裂修复功能类似,Ku蛋白还在保持染色体端粒结构的完整性方面发挥重要作用。在酵母中的研究发现,Ku蛋白能够通过结合端粒酶RNA元件TLC1的48 nt颈环结构,协助招募端粒酶,完成端粒的延伸[11-12]。但随后又有研究表明,人Ku蛋白能够结合端粒酶47 nt的RNA元件hTR,而hTR与TLC1并没有相似序列[13]。Ku70还被证实能够通过结合细胞凋亡促进因子Bax,抑制细胞凋亡。在个体水平上,Ku蛋白还被证实与衰老有关。单独敲除Ku70或Ku80,或者二者都敲除的小鼠,均出现了类似的加速衰老现象[14]。猜测这可能与影响了C-NHEJ途径有关,因为衰老的大鼠神经元细胞内和阿尔兹海默患者脑组织内的C-NHEJ途径均明显减弱。缺失了Ku蛋白的作用后,C-NHEJ途径将进一步减弱,这或许最终导致细胞和个体衰老加速[15-16]。这些研?究结果提示,Ku蛋白的生物学功能十分广泛,还需要进一步挖掘。
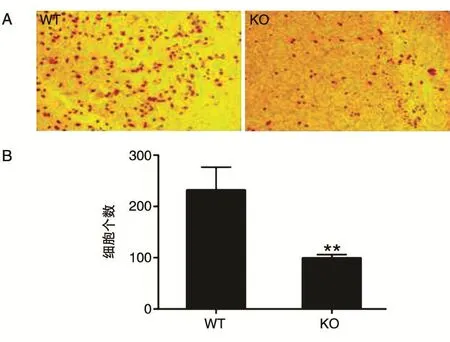
图7 Ku70基因稳定敲除细胞株迁移能力检测(n=3)WT:野生型HeLa细胞;KO:Ku70稳定敲除细胞;**P<0.01
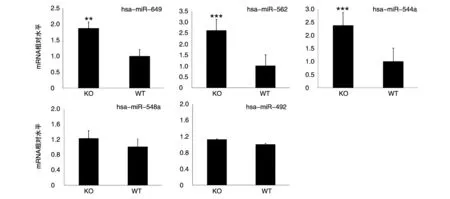
图8 qPCR检测敲除Ku70后HeLa细胞5种miRNA的表达水平(n=3)WT:野生型HeLa细胞;KO:Ku70稳定敲除细胞;**P<0.01,***P<0.001
Ku蛋白结合DNA的特异性早已确定,并且已经有研究证明Ku蛋白与DNA的结合并不是依赖特定序列或碱基的[17]。生物化学分析结果显示,Ku70亚基的3个结构域(ɑ/β结构域、DNA结合结构域、Ku80结合结构域)对于Ku蛋白聚集在双链断裂DNA末端是必需的[18]。细胞实验表明Ku70蛋白能够作为细胞内的模式受体识别外来的病毒DNA,进而介导Ⅲ型干扰素的产生[19]。此外,Ku蛋白的RNA结合特异性也被证实。比如上面提到的Ku蛋白在维持端粒结构的过程中,就发挥了招募端粒酶RNA元件的功能。有报道称Ku70蛋白能够与一类具有颈环结构并且带有一个突出基序的RNA特异结合,这表明Ku70与RNA的结合是空间特异性的[20-21]。
为了便于深入研究Ku70蛋白的生物学功能,我们构建了Ku70基因稳定敲除的HeLa细胞株,为后续实验奠定了基础。我们检测了Ku70稳定敲除细胞株的增殖和迁移能力等生物学功能,结果表明敲除Ku70基因后HeLa细胞增殖和迁移能力均有所减弱,提示Ku70可能参与了HeLa细胞的增殖和迁移过程。此外,我们前期的实验结果提示Ku70蛋白可能调节miRNA表达(结果未显示),而本研究中,我们尝试检测Ku70稳定敲除细胞株中几种可能被Ku70蛋白调节的miRNA表达水平。RT-qPCR结果显示,3种miRNA在敲除Ku70基因的HeLa细胞中明显上调,提示Ku70可能参与了这些miRNA的表达调控,相关的分子机制还有待进一步研究。
[1] Mimori T,Akizuki M,Yamagata H,et al.Characteriza⁃tion of a high molecular weight acidic nuclear protein recognized by autoantibodies in sera from patients with polymyositis-scleroderma overlap[J].Clin Invest, 1981,68(3):611-620.
[2] Mimori T,Steitz H J A.Characterization of the DNA-binding protein antigen Ku recognized by autoantibod⁃iesfrom patients with rheumatic disorders[J].Biol Chem, 1986,261(5):2274-2278.
[3] Vries E,Driel W,Bergsma W G,et al.HeLa nuclear protein recognizing DNA termini and translocating on DNA forming a regular DNA-multimeric protein com⁃plex[J].Mol Biol,1989,208(1):65-78.
[4] Griffith A J,Blier P R,Mimori T,et al.Ku polypep⁃tides synthesized in vitro assemble into complexes which recognize ends of double-stranded DNA[J].Biol Chem,1992,267(1):331-338.
[5] Grundy G J,Moulding H A,Caldecott K W,et al. One ring to bring them all-the role of Ku in mamma⁃lian non-homologous end joining[J].DNA Repair, 2014,17:30-38.
[6] Hammel M,Yu Y,Mahaney B L,et al.Ku and DNA-dependent protein kinase dynamic conformations and assembly regulate DNA binding and the initial non-homologousend joiningcomplex[J].BiolChem, 2010,285(2):1414-1423.
[7] Kragelund B B,Weterings E,Hartmann P R,et al. TheKu70/80 ringin non-homologousend-joining: easy to slip on,hard to remove[J].Front Biosci,2016, 21:514-527.
[8] Boulton S J,Jackson S P.Identification of a Saccharo⁃myces cerevisiae Ku 80 homologue:roles in DNA dou⁃ble strand break rejoining and in telomeric mainte⁃nance[J].Nucleic Acids Res,1996,24(23):4639-4648.
[9] Gravel S,Larrivee M,Labrecque P,et al.Yeast Ku as a regulator of chromo somal DNA end structure[J]. Science,1998,280(5364):741-744.
[10]Sawada M,Sun W,Hayes P,et al.Ku70 suppresses the apoptotic translocation of Bax to mitochondria[J]. Nat Cell Biol,2003,5(4):320-329.
[11]Stellwagen A E,Haimberger Z W,Veatch J R,et al. Ku interactswith telomeraseRNA topromotetelo⁃mere addition at native and broken chromosome ends [J].Genes Dev,2003,17(19):2384-2395.
[12]Fisher T S,Taggart A K,Zakian V A.Cell cycle-de⁃pendent regulation of yeast telomerase by Ku[J].Nat Struct Mol Biol,2004,11(12):1198-1205.
[13]Ting N,Yu Y,Pohorelic B,et al.Human Ku70/80 in⁃teracts directly with hTR,the RNA component of hu⁃man telomerase[J].Nucleic Acids Res, 2005,33(7): 2090-2098.
[14]Li H,Vogel H,Holcomb V B,et al.Deletion of Ku70,Ku80,or both causes early aging without sub⁃stantially increased cancer[J].Mol Cell Biol,2007,27 (23):8205-8214.
[15]Vyjayanti V N,Rao K S.DNA double strand break repair in brain:reduced NHEJ activity in aging rat neurons[J].Neurosci Lett,2006,393(1):18-22.
[16]Shackelford D A.DNA end joining activity is reduced in Alzheimer's disease[J].Neurobiol Aging,2006,27(4): 596-605.
[17]Walker J R,Corpina R A,Goldberg J.Structure of the Ku heterodimer bound to DNA and its implica⁃tions for double-strand break repair[J].Nature,2001, 412(6847):607-614.
[18]Koike M,Yutoku Y,Koike A.Accumulation of Ku70 at DNA double-strand breaks in living epithelial cells [J].Exp Cell Res,2011,317(17):2429-2437.
[19]Zhang X,Brann T W,Zhou M,et al.Ku70 is a nov⁃el cytosolic DNA sensor that induces type III rather than type I IFN[J].J Immunol,2011,186(8):4541-4545.
[20]Andrey N A,Ekaterina S K,Timofey S Z,et al.Hu⁃man Ku70 protein bindshairpin RNA and double stranded DNA through two different sites[J].Biochimie, 2017,132:85-93.
[21]Andrew B D,Karen J G,Jennifer S P,et al.RNA recognition by the DNA end-binding Ku heterodimer [J].RNA,2013,19(6):841-851.
猜你喜欢
江西农业学报(2021年4期)2021-04-20
中国现代医药杂志(2020年10期)2020-12-14
现代检验医学杂志(2016年3期)2016-11-15
西南医科大学学报(2015年1期)2015-08-22
医学研究杂志(2015年11期)2015-06-10
医学研究杂志(2015年3期)2015-06-10
特产研究(2015年1期)2015-04-12
中国当代医药(2015年16期)2015-03-01
中国当代医药(2015年9期)2015-03-01
中国医药导报(2015年27期)2015-02-28
