
电针对失神经大鼠骨骼肌自噬相关基因表达的影响
2018-03-30 08:36:47吴梦佳唐成林赵丹丹罗翱安荟羽谭程方邱丽
中国康复理论与实践 2018年3期
吴梦佳,唐成林,赵丹丹,罗翱,安荟羽,谭程方,邱丽
1.重庆医科大学中医药学院,重庆市400016;2.中医药防治代谢性疾病重庆市重点实验室,重庆市400016
周围神经损伤后,失神经支配的骨骼肌发生快速萎缩,肌蛋白分解率增加,骨骼肌质量迅速下降,极大影响骨骼肌功能的维持和恢复[1]。失神经骨骼肌萎缩发生时,自噬相关基因活性升高[2]。自噬-溶酶体系统通过调控骨骼肌细胞的代谢和稳态,对维持骨骼肌质量的稳定发挥重要作用[3]。临床研究表明,针灸对骨骼肌萎缩的治疗效果显著[4];电针能有效延缓失神经大鼠骨骼肌萎缩,但其具体机制仍有待明确[5]。本课题组前期研究表明,电针能通过上调失神经肌萎缩大鼠模型中胰岛素样生长因子-1(insulin-like growth factor-1,IGF-1)的表达、下调肌肉生长抑制素(myostatin)的表达,促进肌卫星细胞的增殖[6];并上调Bcl-2的表达,下调caspase-3等凋亡相关因子的表达[7],从而延缓失神经骨骼肌萎缩。本研究观察电针对失坐骨神经大鼠腓肠肌中自噬相关基因(autophagy-related gene,Atg)表达的影响,从肌萎缩后自噬活性水平的角度,探讨电针延缓失神经骨骼肌萎缩的可能机制。
1 材料与方法
1.1 实验动物及分组
8周龄清洁级健康雄性Sprague-Dawley大鼠21只,体质量270~290 g,重庆医科大学动物实验中心提供,动物生产许可证号SCXK(渝)2012-0002。饲养于SPF级动物房。
大鼠适应性喂养7 d后,将大鼠编1~21号,使用计算机自动生成21个两位数随机数字,每个编号对应一个随机数字,将随机数字从小到大依次排列序号,其中序号1~7为假手术组,8~14为模型组,15~21为电针组。实验过程中动物处置均按照重庆医科大学伦理委员会标准进行。
1.2 主要试剂与仪器
Trizol总RNA提取试剂盒、RR037A逆转录试剂盒、SYBR Premix Ex Taq TMII、ULK1、Atg13、Beclin1、Atg14、Atg7、Atg12、Atg5、Atg16L1、GAPDH引物合成:TAKARA公司。
SDZ-Ⅱ型电子针治疗仪:苏州医疗用品有限公司。汉医牌无菌针灸针:北京汉医医疗器械中心。柔软型实验大鼠固定器:温州原上草医疗科技有限公司。AL204型电子天平:瑞士METTLER TOLEDO公司。低温高速离心机:日本SIGMA公司。Thermo ND 2000超微量核酸蛋白测定仪:上海GENE公司。CFX PCR检测系统、T100TMPCR仪:美国BIO RAD公司。CellSens Standard图像采集软件、BX53普通正置显微镜:日本OLYMPUS公司。
1.3 造模方法
制备切断大鼠坐骨神经腓肠肌萎缩模型[8]。大鼠4%水合氯醛8 ml/kg腹腔注射麻醉,俯卧固定于手术台,无菌条件下右后肢手术区备皮。于大鼠右后肢坐骨结节下缘外侧切口6~8 mm,玻璃分针沿肌肉走向钝性分离肌肉与筋膜,找到并暴露坐骨神经;取中段切断,造成1.0 mm神经缺损。逐层缝合肌肉与皮肤,清洁伤口。
假手术组只暴露神经,不行坐骨神经截断术。
1.4 电针干预
造模1 d后,用大鼠固定器[9]固定电针组大鼠,选右侧足三里、环跳穴[10],使用直径0.25 mm、长13 mm针灸针,直刺5~7 mm;接电针仪,正极连接足三里,负极连接环跳,连续波,强度1.0 mA,频率2 Hz,以大鼠下肢稍震动为宜,每次10 min,每天1次,每周6次,连续干预8周。
假手术组和模型组每天同法固定,但不进行电针干预。
1.5 标本采集
各组大鼠于造模后8周取材。大鼠4%水合氯醛8 ml/kg腹腔注射麻醉,无菌条件下完整剥离双侧腓肠肌,去除多余肌腱头,PBS冷浴;吸干表面残留液体。术侧腓肠肌组织分为两份,一份快速修剪后置4%多聚甲醛中固定,用于形态学检测;一份锡箔纸包裹,放入液氮罐中速冻,-80℃冰箱保存待测。
1.6 检测方法
1.6.1 腓肠肌湿重比
大体观察双侧腓肠肌大小。观察后迅速完整剥离双侧腓肠肌,将多余血管及神经清除,于电子天平上称取肌湿重,以健侧作为对照,计算腓肠肌湿重比。
1.6.2 腓肠肌纤维截面积及直径
腓肠肌4%多聚甲醛固定后,蔗糖梯度脱水3 d,OTC包埋,制备厚8 μm冰冻切片。晾干,行HE染色,过二甲苯溶液2次,每次5 min;梯度乙醇浸泡4次,每次2 min,清水冲洗2 min;苏木素染色2 min,冲洗;浸于分化液中40 s,冲洗;伊红染色80 s,冲洗;梯度乙醇浸泡2次,每次2 min;二甲苯溶液浸泡4 min;树胶封片,晾干。200倍光学显微镜下观察切片,每张切片随机选取5个视野拍照,使用Image-Pro Plus 6.0图像分析软件计算腓肠肌纤维截面积和直径。
1.6.3 逆转录实时定量聚合酶链反应
腓肠肌组织样本约50 mg,提取组织上清,将Trizol液加入管中,剪碎组织,充分匀浆,离心,提取总RNA;异丙醇沉淀后离心,75%乙醇清洗3次后晾干,加入无酶水待检。清洗超微量核酸蛋白测定仪探头3次,记录总RNA 260/230和260/280值,计算稀释比例。-20℃取出SYBR Green,按说明书配制反应体系,合成反应体系,设置反应条件行逆转录:37℃15 min,85℃5 s,4℃+∞。加样,标板后上机行定量聚合酶链反应。使用CFX Manager 3.1软件读取Ct值,计算2-△△Ct。上下游引物序列见表1。
1.7 统计学分析
采用SPSS 22.0统计软件进行分析。实验结果均用(xˉ±s)表示,采用单因素方差分析;方差齐时,组间两两比较采用Bonferroni检验,方差不齐时采用Dunnett's T3检验。显著性水平α=0.05。
2 结果
2.1 腓肠肌湿重比
与假手术组相比,模型组和电针组术侧腓肠肌湿重比显著降低(p<0.001),电针组高于模型组(p<0.05)。见表2。
2.2 形态学
假手术组肌纤维边缘规则,形态正常;模型组肌纤维缩小,边缘不规则;电针组较模型组肌纤维大且边缘较规则。见图1。与假手术组相比,模型组和电针组术侧腓肠肌纤维截面积和直径显著降低(p<0.001),电针组显著高于模型组(p<0.001)。见表2。

表1 待测基因引物序列
2.3 聚合酶链反应
与假手术组相比,模型组术侧腓肠肌ULK1、Atg13、 Beclin1、 Atg14、 Atg7、 Atg12、 Atg5 和Atg16L1 mRNA表达显著升高(p<0.001);与模型组相比,电针组以上mRNA表达量均下降(p<0.05)。见表3。
3 讨论
根据靶器官肌肉损伤后的临床特征,失神经骨骼肌萎缩属传统中医学“痿症”范畴,主要表现为肌肉瘦削、酸困且痿软无力[11]。据“治痿独取阳明”的理论,选取足阳明胃经合穴足三里穴,以强筋健骨,补益气血;局部取足少阳胆经环跳穴,以通经活络,便利腰腿。尚画雨等[12]发现,针刺大鼠腓肠肌可有效下调大负荷运动后腓肠肌中Pink1和Parkin的表达,抑制线粒体自噬过度激活,减轻骨骼肌损伤,促进修复。提示电针延缓骨骼肌萎缩的治疗作用可能与调节骨骼肌细胞自噬活性有关。
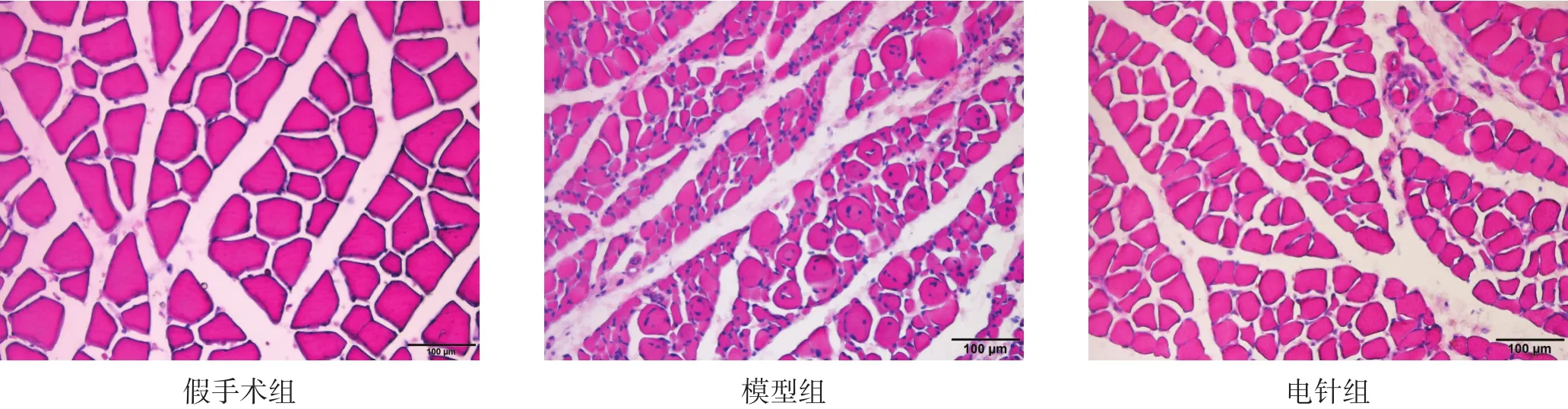
图1 大鼠术侧腓肠肌纤维病理学改变(HE染色,bar=100 μm)
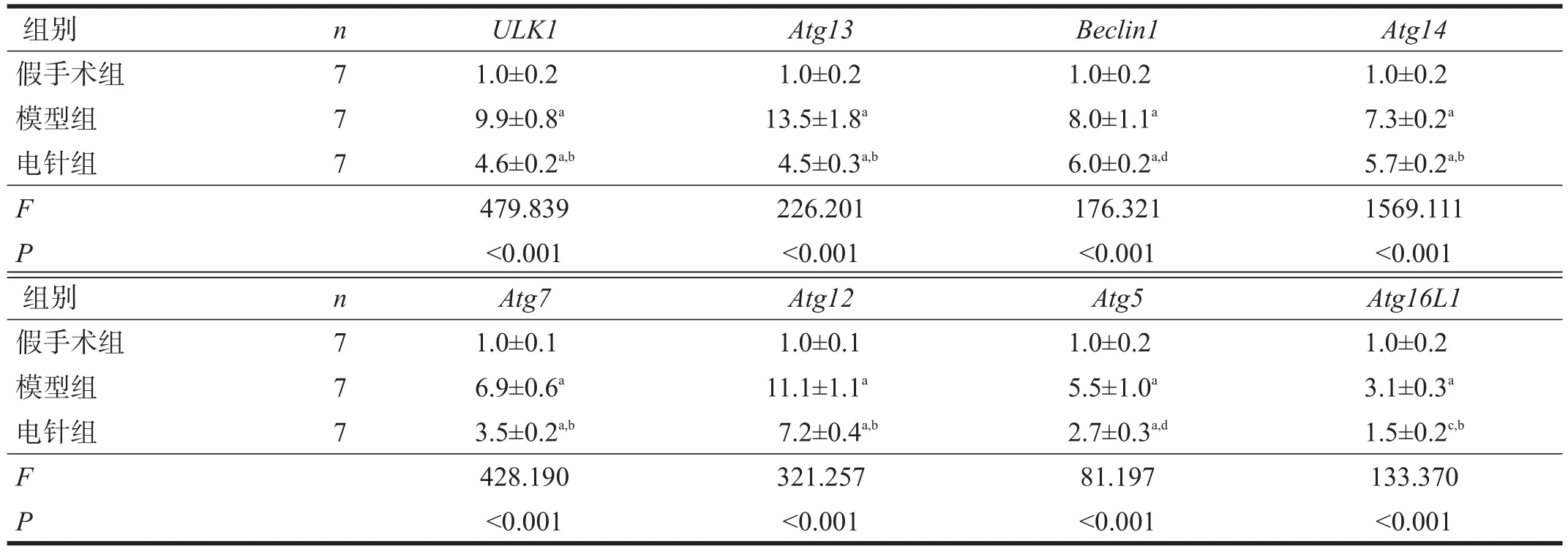
表3 各组大鼠自噬相关基因表达比较(2-△△Ct)
自噬-溶酶体系统是控制骨骼肌蛋白质代谢的重要途径之一[13],在肌肉体积控制及维持细胞自身稳态方面发挥重要作用[14]。自噬过程大致分为4个阶段:自噬诱导、自噬体形成、自噬溶酶体形成和内容物降解;自噬相关基因参与细胞自噬的全过程,发挥着重要作用[15]。ULK1是酵母Atg1在哺乳动物的同源基因,与调节成分Atg13共同构成复合物,增强Atg13稳定性,在自噬诱导阶段起关键作用[16]。Beclin1是酵母Atg6在哺乳动物的同源基因,与调节成分Atg14等参与小囊泡形成,其表达强度与自噬活性密切相关[17]。Atg5和Atg12是影响自噬体泛素样蛋白修饰的重要基因[18],其形成的复合体在自噬体膜的延伸过程中发挥重要作用,促使自噬小体不断形成。Atg7是辅助自噬体伸展扩张的重要基因。在自噬体形成期,Atg7通过水解ATP获能,进而激活Atg12,在Atg10的催化下,Atg12与Atg5结合,进而募集Atg16L1形成多聚复合体,有利于自噬体的伸展扩张[19],促进自噬活性增强。
李妍等[20]在慢性阻塞性肺疾病大鼠模型中发现,在萎缩的骨骼肌组织中,FoxO转录因子及自噬相关因子Beclin1、Atg5、Bnip3等升高,自噬-溶酶体系统被激活,骨骼肌萎缩加重;敲除小鼠Atg7以抑制自噬,也出现肌力下降、肌肉萎缩,并没有出现维持肌肉稳态的结果[21];而Laminin-2自噬缺陷小鼠出现肌萎缩和营养不良等症状,却是由于自噬过度激活所致[22]。有研究表明[23],胰岛素缺乏的糖尿病大鼠骨骼肌中哺乳动物雷帕霉素靶蛋白(mammalian target of rapamycin,mTOR)受到抑制,FoxO3被激活,导致自噬过度激活,骨骼肌发生萎缩,肌肉流失。耐力运动可以激活mTOR,降低自噬水平,减轻自噬过度激活,从而减少蛋白质等的过度降解,维持骨骼肌稳态[24]。Liu等[25]研究发现,KLHL20可以通过调控ULK1和Vps34复合物的代谢,控制自噬的终结,防止因过度自噬引起的肌肉损失。Sato等[26]发现,赖氨酸可减少C2C12肌管细胞中肌纤维蛋白的降解,延缓肌肉萎缩,其机制可能是通过激活mTOR,抑制自噬溶酶体系统活性的过表达。以上研究提示,自噬水平的高低与骨骼肌细胞稳态的维持紧密相关,不足或过度的自噬都不利于骨骼肌健康,自噬过程的关闭和防止自噬过度激活,对保持肌肉结构稳态,防治肌肉萎缩意义重大。
临床治疗时,由于不能及时到找适合的神经供体进行移植,等待手术的过程往往又比较漫长,不可逆的失神经肌萎缩将得以持续。维持骨骼肌细胞稳态,保持其相对活性,延缓骨骼肌萎缩是进行神经移植手术,提高移植后骨骼肌适应性的关键。本研究表明,失坐骨神经8周后,大鼠腓肠肌中自噬相关基因表达上调,自噬活性增强;电针干预后自噬活性下降,骨骼肌萎缩减轻。这一结果与其他研究团队通过手针、运动、药物等干预方式的研究结果一致[12,24-25],可为临床进行神经移植术、提高神经移植后骨骼肌功能恢复,争取宝贵的治疗时间。
自噬作为一种自我调节机制,在机体中发挥着“双刃剑”的作用[27]。针灸具有调节阴阳平衡的作用。在失神经1周和4周时,电针可通过上调自噬相关基因表达,促进坐骨神经修复[28]。因此,在较长时间失神经过程中,自噬对于萎缩骨骼肌的具体影响,以及电针如何对其进行调控以延缓失神经肌萎缩的具体机制,还有待进一步研究说明。
综上所述,我们推测,电针延缓大鼠失神经骨骼肌萎缩的机制可能是通过调控自噬相关基因的表达,影响肌萎缩的自噬活性,抑制自噬水平过度激活,从而维持肌细胞稳态来实现的。
[1]Lang F,Aravamudhan S,Nolte H,et al.Dynamic changes in the mouse skeletal muscle proteome during denervation-induced atrophy[J].Dis Model Mech,2017,10(7):881-896.
[2]Oâ Leary MFN,Hood DA.Denervation-induced oxidative stress and autophagy signaling in muscle[J].Autophagy,2009,5(2):230-231.
[3]Schiaffino S,Dyar KA,Ciciliot S,et al.Mechanisms regulating skeletal muscle growth and atrophy[J].FEBS J,2013,280(17):4294-4314.
[4]谢运华,李震华,李柳,等.针刺结合改良神经外膜束膜联合缝合术治疗周围神经损伤的临床观察[J].上海中医药杂志,2014,48(10):54-56.
[5]吴珍元,黄英如,冼华,等.电针对失神经骨骼肌萎缩及纤维化的影响[J].中国康复医学杂志,2016,31(2):177-182.
[6]高睿琦,唐成林,曹净,等.电针对失神经骨骼肌萎缩大鼠胰岛素样生长因子1、肌肉生长抑制素及肌卫星细胞增殖的影响[J].中国康复理论与实践,2016,22(11):1259-1263.
[7]高睿琦,唐成林,黄思琴,等.电针对失坐骨神经大鼠腓肠肌细胞凋亡及相关蛋白的影响[J].针刺研究,2017,42(4):302-307.
[8]彭建平,何继业,王栋梁,等.骨髓间充质干细胞成肌诱导后移植对延缓失神经肌肉萎缩的作用[J].中国组织工程研究,2008,12(25):4824-4828.
[9]张毅,唐成林,田源,等.电针联合饮食调控对非酒精性脂肪性肝病大鼠过氧化物酶体增殖物激活受体α和肝型脂肪酸结合蛋白的影响[J].针刺研究,2015,40(5):345-351.
[10]李忠仁.实验针灸学[M].北京:中国中医药出版社,2003:316.
[11]唐雪梅.《内经》痿证理论及其临床应用[J].南京中医药大学学报,2005,21(6):350-352.
[12]尚画雨,付玉,夏志,等.针刺对运动性骨骼肌损伤大鼠骨骼肌线粒体自噬的影响[J].中国病理生理杂志,2017,33(11):2038-2046.
[13]黄燕峰,魏连波.骨骼肌蛋白代谢及其调控机制研究进展[J].广东医学,2015,36(23):3728-3731.
[14]Whitehead NP,Kim MJ,Bible KL,et al.A new therapeutic effect of simvastatin revealed by functional improvement in muscular dystrophy[J].Proc Natl Acad Sci USA,2015,112(41):12864-12869.
[15]Mizushima N,Yoshimori T,Ohsumi Y.The role of Atg proteins in autophagosome formation[J].Annu Rev Cell Dev Biol,2011,27(1):107-132.
[16]Chang HJ,Chang BJ,Ro SH,et al.ULK-Atg13-FIP200 complexes mediate mTOR signaling to the autophagy machinery[J].Mol Biol Cell,2009,20(7):1992-2003.
[17]Matsunaga K,Saitoh T,Tabata K,et al.Two Beclin 1-binding proteins,Atg14L and Rubicon,reciprocally regulate autophagy at different stages[J].Nat Cell Biol,2009,11(6):385-396.
[18]Sheng Y,Song Y,Li Z,et al.RAB37 interacts directly with ATG5 and promotes autophagosome formation via regulating ATG5-12-16 complex assembly[J].Cell Death Differ,2017.doi:10.1038/s41418-017-0023-1.[Epub ahead of print].
[19]Itakura E,Mizushima N.Characterization of autophagosome formation site by a hierarchical analysis of mammalian Atg proteins[J].Autophagy,2010,6(6):764-776.
[20]李妍,韩锋锋,李艳利,等.自噬-溶酶体系统在慢性阻塞性肺疾病大鼠骨骼肌萎缩中的作用机制[J].中国呼吸与危重监护杂志,2015,14(6):534-540.
[21]Masiero E,Agatea L,Mammucari C,et al.Autophagy is required to maintain muscle mass[J].Cell Metab,2009,10(6):507-515.
[22]Bonaldo P,Sandri M.Cellular and molecular mechanisms of muscle atrophy[J].Dis Model Mech,2013,6(1):25-39.
[23]Lv P,Huang J,Yang J,et al.Autophagy in muscle of glucose-infusion hyperglycemia rats and streptozotocin-induced hyperglycemia rats via selective activation of m-TOR or FoxO3[J].PLoS One,2014,9(2):e87254.
[24]钱帅伟,丁树哲.运动性细胞自噬是调节骨骼肌代谢稳态的内置机制[J].体育科学,2015,35(10):55-65.
[25]Liu CC,Lin YC,Chen YH,et al.Cul3-KLHL20 ubiquitin ligase governs the turnover of ULK1 and VPS34 complexes to control autophagy termination[J].Mole Cell,2016,61(1):84-97.
[26]Sato T,Ito Y,Nedachi T,et al.Lysine suppresses protein degradation through autophagic-lysosomal system in C2C12 myotubes[J].Mol Cell Biochem,2014,391(1-2):37-46.
[27]Clavel M,Michaeli S,Genschik P.Autophagy:a double-edged sword to fight plant viruses[J].Trends Plant Sci,2017,22(8):646-648.
[28]赵伟,刘延祥,王占魁,等.自噬对电针治疗大鼠实验性坐骨神经损伤后神经再生的影响[J].天津中医药大学学报,2014,33(6):351-354.
猜你喜欢
分子诊断与治疗杂志(2023年8期)2023-09-07 12:00:34
大电机技术(2022年3期)2022-08-06 07:48:24
核科学与工程(2021年4期)2022-01-12 06:30:04
煤气与热力(2021年4期)2021-06-09 06:16:54
中华戏曲(2020年1期)2020-02-12 02:28:18
中成药(2018年10期)2018-10-26 03:41:40
中国运动医学杂志(2016年3期)2016-07-10 12:07:23
中国运动医学杂志(2016年3期)2016-07-10 12:07:23
磁共振成像(2015年5期)2015-12-23 08:52:53
医学研究杂志(2015年5期)2015-06-10 06:43:26
