
分段拟合分析温度对垃圾发酵的影响
2018-03-30 06:56:10刘盛涛郑有飞
中国沼气 2018年1期
刘盛涛, 郑有飞
(1.南京信息工程大学 环境科学与工程学院, 江苏 南京 210044; 2.江苏省大气环境与装备技术协同创新中心, 江苏 南京 210044)
随着我国城市化水平逐步提高,城市生活垃圾产量也在逐年增加。生活垃圾会造成一系列环境与社会问题,如产生恶臭气味、渗滤液、滋生病菌等。城市生活垃圾的卫生填埋是一种常用的垃圾处理方法,该方法通过采取必要的防护措施,以达到被处置废物与环境生态系统最大限度的隔绝的目的,是生活垃圾的最终处置手段。据统计,2015年我国城市生活垃圾清运量达1.91亿吨,其中1.15亿吨通过卫生填埋处理,占垃圾无害化处理总量的63.8%。
垃圾填埋场是CH4的重要排放源之一,此前众多学者对填埋场的CH4排放进行了大量观测。如刘鸿霆[1]研究某填埋场后,得出夜间到凌晨排放速率最低,中午最高,下午随时间递减的规律;杨雪[2]等,则通过实验得出了排放通量下午大于中午大于上午的结论,且释放通量变化较小;马占云[3]等,通过长期观测封场区甲烷排放情况得出了甲烷日变化不存在规律性的结论。这些结论存在较大差异,同时在采样现场对垃圾产气及气体迁移过程有较多影响因素,用此前对垃圾发酵过程的研究结论难以解释。
此前研究普遍认为垃圾发酵产气的最适温度分别是37℃和55℃[4-5]。有研究表明,温度对垃圾发酵的不同阶段有不同影响。李阳等人的研究发现垃圾发酵过程中25℃条件下,乙酸产量最大[6,15,17];侯贵光等人则根据有机物厌氧产生沼气的生化机理,建立两步一级反应耦合模型,认为垃圾发酵可分为两个主要阶段[7]。但目前对于温度对各个产气阶段影响的研究不多,因此笔者通过对垃圾产气曲线分段后分别进行回归分析,研究温度对各个产气阶段的影响。
1 材料与方法
1.1 垃圾来源及预处理
为避免垃圾组分变化对实验造成影响,实验将特定组分垃圾按一定比例混合成试验用垃圾,所用垃圾及所占比例如表1所示,该垃圾组分比例参照填埋场垃圾样品组分确定。其中厨余类垃圾采用新鲜的苹果、梨及芦柑代替,三种水果鲜重各占厨余组分的1/3。所有垃圾在实验前破碎成1~2 cm小块,混合均匀后备用。接种物选用此前相同组分垃圾发酵产生的渗滤液。

表1 实验垃圾组分 (%)
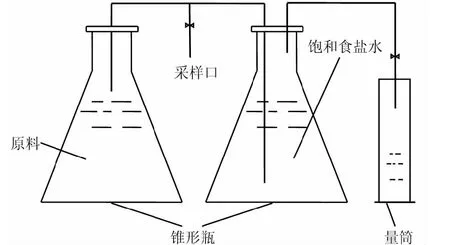
图1 实验装置
1.2 实验方法
称取125 g经处理的混合垃圾与500 mL锥形瓶中,接种物与垃圾按1∶10的质量比接种,混合均匀后置于恒温培养箱中培养,通过排水集气法收集气体,实验装置如图1所示。实验设置4个培养温度,分别为15℃,25℃,35℃及45℃,每个温度设置多个平行样,同时设置一组仅加入接种物的锥形瓶作为空白样。实验开始后每小时记录产气量,同时用针筒采集气样,通过气相色谱仪分析CO2及CH4浓度
2 结果与讨论
2.1 分段拟合
垃圾发酵过程可分为水解产酸、产甲烷及衰减阶段,各阶段反应转变需要一定时间,这为产气阶段分段造成困难。通常实验仅通过检测CH4浓度变化判断发酵阶段,但分段较为粗糙。本实验则通过分析发酵阶段转换造成的产气速率变化对垃圾发酵进行分段,同时通过检测CO2及CH4浓度进行确认,提高了分段的精度。笔者通过对多组平行样取平均值,对不同的产气阶段进行回归分析,通过最小二乘法估算产气阶段中满足线性关系的线性回归方程参数,并通过origin分析产气阶段中满足非线性关系的回归方程,以避免发酵过程中产气量波动对分析产生影响。
如下图所示,图2~图5中分别为15℃,25℃,35℃以及45℃条件下,垃圾的产气量峰值期、稳定期和衰减期的回归曲线。
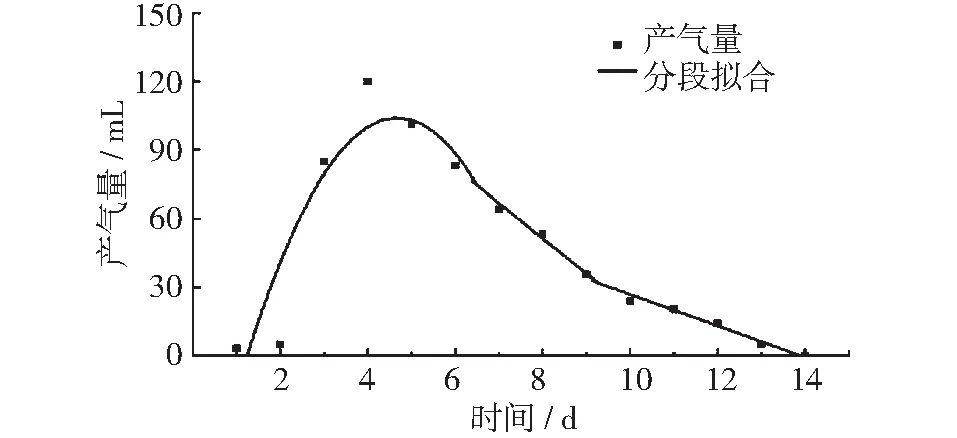
图2 15℃垃圾产气量及分段拟合曲线
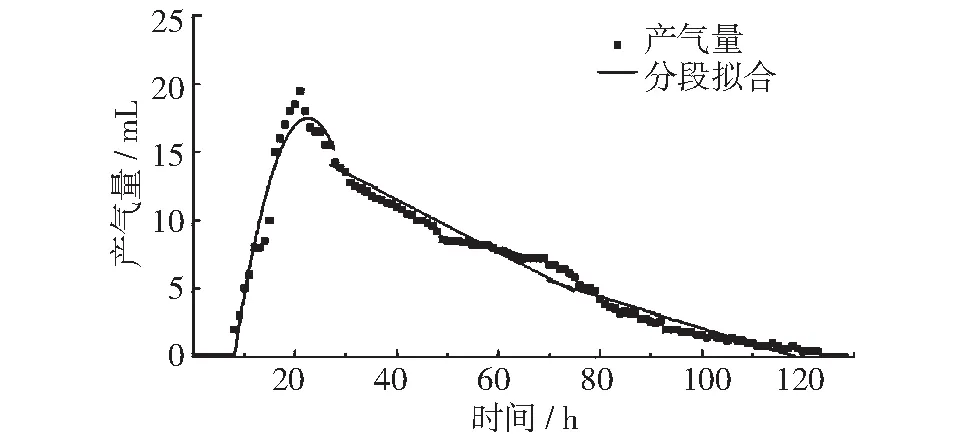
图3 25℃垃圾产气量及分段拟合曲线

图4 35℃垃圾产气量及分段拟合曲线
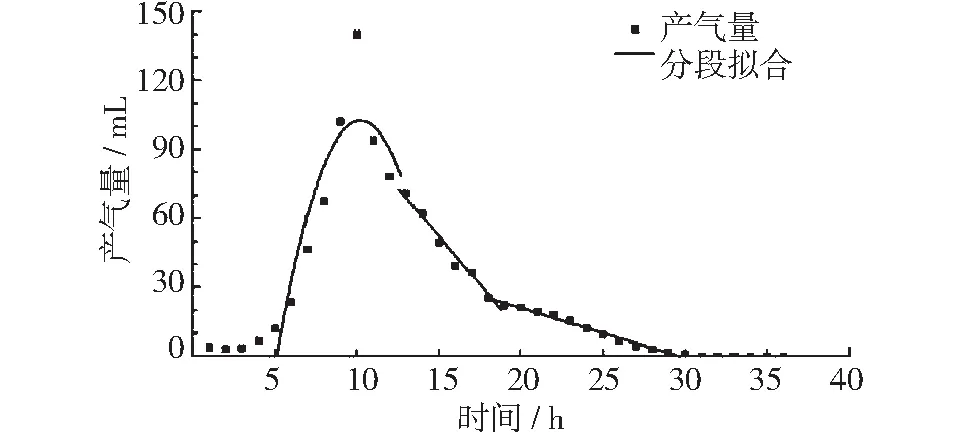
图5 45℃垃圾产气量及分段拟合曲线
拟合结果的回归方程如下:
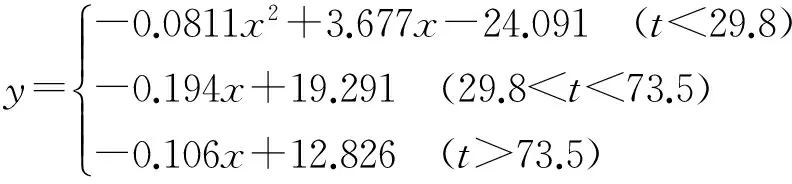
(2)
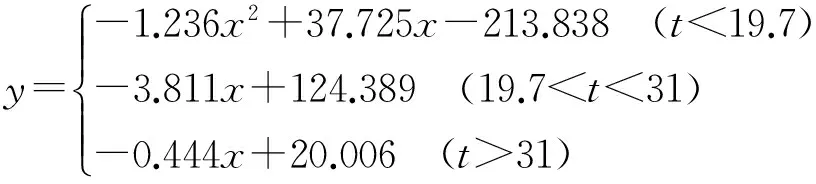
(3)

(4)
拟合结果的p值及r2如表2所示。
由各个产气阶段p值可知,各个产气阶段的拟合曲线与实际产气情况是显著相关的。除15℃的峰值期的拟合曲线0.05>p>0.01外,其他阶段的拟合曲线p值均小于0.01,与实际产气量,表现出高度显著相关性;而15℃峰值期的拟合曲线与实际产气量也表现出显著相关性。此外,根据拟合曲线的r2同样可以认为,拟合曲线对实际产气量的拟合有较好的相关性,与实际产气曲线相符。因此可以认为拟合效果较好,拟合曲线能准确反映实际产气量的变化。
2.2 温度与不同阶段产气量的关系
通过对拟合曲线积分可得垃圾发酵各阶段的产气量,各阶段产气量叠加如图6所示。为了进一步了解温度对不同阶段产气量的影响,笔者分别对各阶段产气量进行分析。

表2 拟合曲线的p值及r2
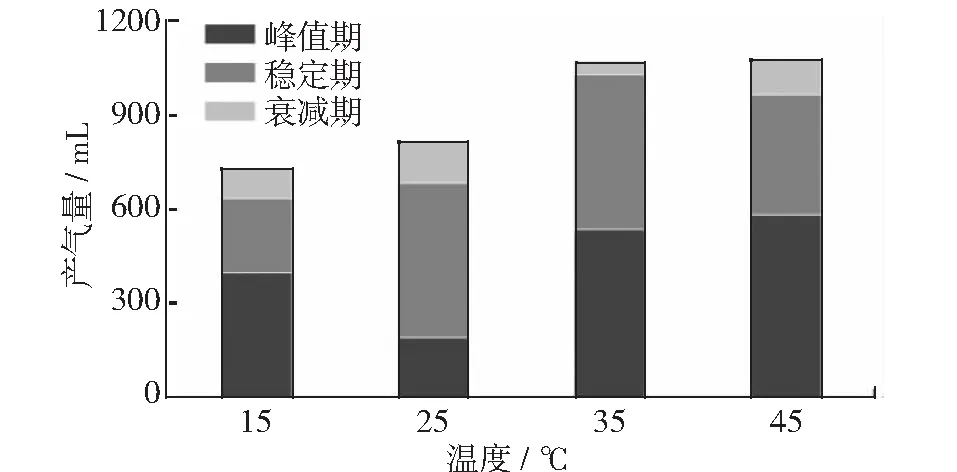
图6 温度与累积产气量
由图7~图10所示,分别为峰值期、稳定期、衰减期的累积产气量以及总累积产气量与温度的关系曲线。
垃圾发酵过程主要可以分为:水解产酸阶段、产甲烷阶段以及衰减阶段。其中,图7为不同温度条件下峰值期的产气量曲线,对应为水解产酸阶段,产气量先随温度升高而下降,在25℃达到极小值,随后上升,在35℃达到极大值。温度较低时,尤其是15℃以下的温度会影响水解产酸阶段的启动,即影响整个发酵过程的启动。在实验的低温段即15℃和25℃两组实验,虽然25℃的峰值期累积产气量较15℃的峰值期累积产气量有小幅减少,但与其他两组实验相比,这两组实验的峰值期累积产气量差别不大,由此可以推断水解产酸阶段在低温段时启动缓慢,产气量也较少,并且产生的乙酸等产甲烷阶段所需原料较少,不利于后续发酵的进行。当温度升高至35℃时,峰值期累积产气量迅速上升,水解产酸反应强烈,峰值期累积产气量是15℃与25℃峰值期累积产气量的2倍;而温度升高至45℃时,峰值期累积产气量有所下降,但与15℃及25℃相比仍处于较高水平,由此可以推断,水解产酸阶段即发酵峰值期的适宜温度在35℃~45℃,在这一温度段内,垃圾发酵的水解产酸阶段反应强烈,有机物分解产生大量的CO2及乙酸等后续产甲烷阶段所需原料,有利于发酵的继续进行。为了进一步确定温度与产气量关系,笔者尝试对其进行线性及非线性回归拟合,发现拟合方程的r2分别为0.526及0.537。这一结果表明峰值期累积产气量与温度有关,但由于样本数量原因无法拟合出相应的关系函数。有关研究表明,这一阶段的产气主要来自垃圾发酵产酸产生的CO2,乙酸和CO2所含有机碳占原总有机碳量的比例分为:乙酸2/3,二氧化碳1/3[7]。李阳[6,15]等人的研究发现,温度控制在25℃~35℃时餐厨垃圾发酵可获得较高乙酸产量;而赵宋敏[8]等人的研究则发现,温度在20℃~37℃时餐厨垃圾发酵产生的CO2随温度升高而升高,37℃时厨余垃圾发酵产酸量最大,超过37℃后CO2及乙酸产量迅速下降。以上研究结果与本实验结果相吻合。但由于样本较少的原因,确定温度与峰值期产气量关系函数需要通过进一步缩小各实验组之间的温度差距,进行多组实验。
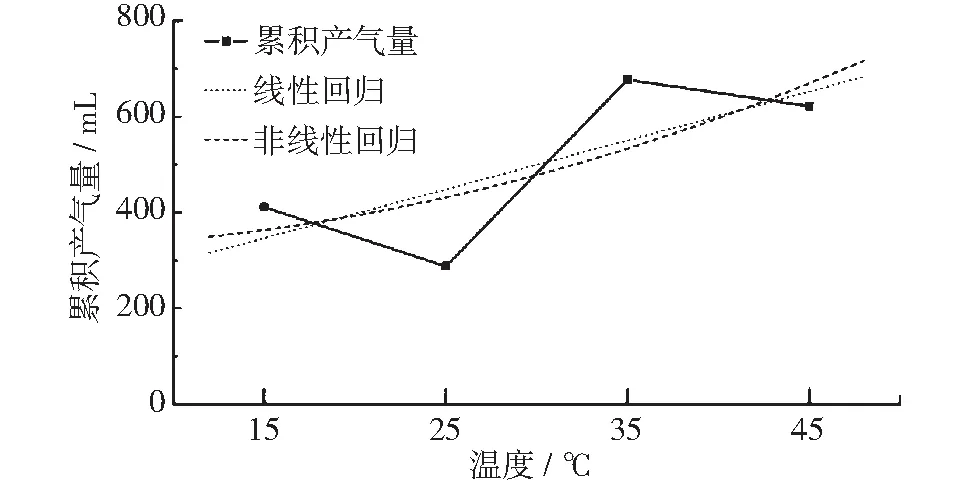
图7 峰值期累积产气量与温度关系
图8为不同温度条件下稳定期的产气量曲线,对应为产甲烷阶段,产气量随温度升高先升高再降低,在25℃达到极大值。对其进行非线性回归拟合,拟合方程的r2为0.816,这表明稳定期累积产气量与温度有较强相关性。通过拟合方程可以推算出稳定期累积产气量在温度达到31℃时达到最大值。这一阶段产气主要由甲烷菌利用此前反应产物代谢产生CH4和CO2,此时的有机酸是甲烷菌的营养限制因素,可假设为一级反应[9]。实验结果则显示,乙酸产量最低的25℃实验组,在这一阶段的产气量最高;拟合结果也表明产气量最高点在31℃左右,并非上一阶段的产气峰值温度。赵宋敏等人及马宗虎等人的研究结果表明,在整个厌氧发酵阶段,pH值并没有随温度变化发生较大波动[8,10],由此可排除pH值对产气量的影响。因此可以认为,该阶段的最佳产气温度应在31℃左右。
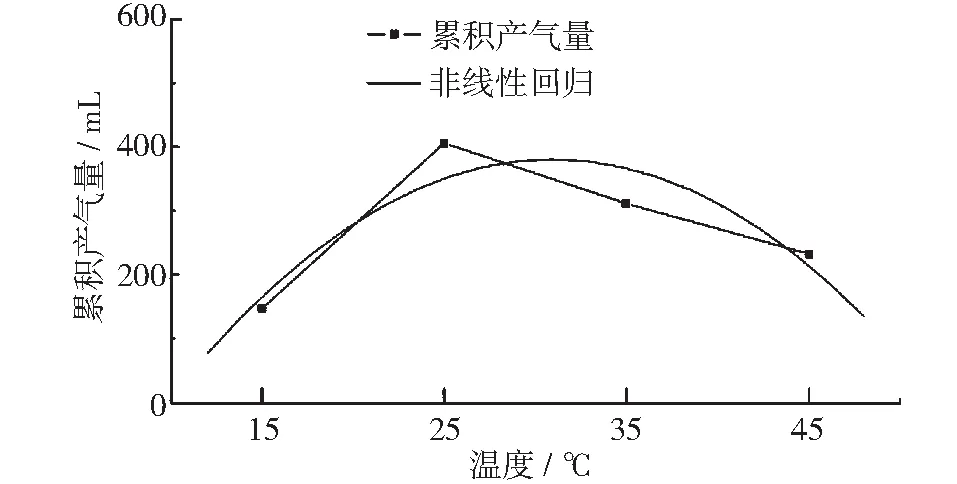
图8 稳定期累积产气量与温度关系
图9为不同温度条件下衰减期的产气量曲线,与峰值期相反,产气量先随温度升高而升高,在25℃达到极大值,随后下降,在35℃达到极小值后迅速上升。对其进行线性及非线性回归拟合发现拟合方程的r2仅0.255及0.141。这一结果表明,温度与衰减期产气量的相关性较弱。此外,比较相同温度下平行样的衰减期产气量发现,样本间衰减期产气量有很大波动,标准差是峰值期、稳定期产气量的5倍,不确定性较大。综上判断,衰减期的累积产气量与温度没有显著关联。其原因在于衰减期的产气基于之前两个阶段产气的剩余物质以及菌种情况,而每组的平行样之间虽然前两个产气阶段产气速率差距不大,但仍存在不符差异,这些差异会在产气后期的衰减期被放大,从而导致衰减期产气量的波动。
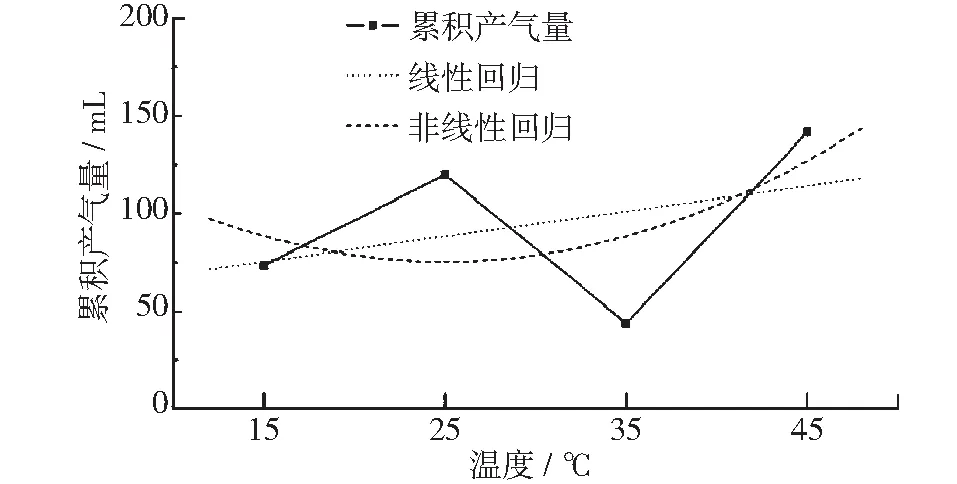
图9 衰减期累积产气量与温度关系
图10为不同温度条件下总累积产气量曲线。产气量随温度升高先升高再降低,在35℃时达到极大值。通过多项式拟合得出的拟合方程的r2为0.937,拟合结果非常接近观测值。通过拟合方程可推算出累积产气量在38.37℃时达到最大值。该结果与实际观测值相符,与此前文科军[11],Y C Song[12],吴满昌[13]等多人的研究结果相同。
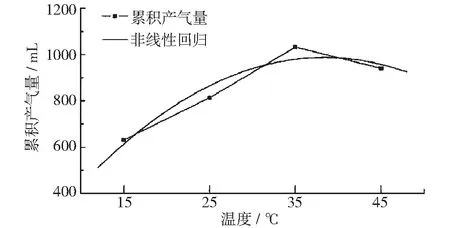
图10 总累积产气量与温度关系
以上结果表明,温度对垃圾发酵的不同产气阶段有不同影响,不同时期的最大产气温度有较大区别。其中稳定期产气量与温度相关性最为显著,在31℃时产气量最大;峰值期产气量与温度关系需要进一步实验探讨;衰减期产气量与温度相关性较弱,不确定性较大。
这一结果在一定程度上解释了国内外对填埋场填埋气排放观测结果的差异。此前诸多学者对填埋场的填埋气排放进行了观测研究[1-3,14,18],但结果有较大差异,尤其在温度对填埋气排放的影响上分歧较大。而本实验结果表明,温度对填埋气排放的影响取决于填埋场内填埋垃圾的产气阶段。垃圾体在不同的产气阶段所适宜的最佳产气温度有很大差别,但填埋场填埋垃圾时间跨度长,组分复杂,因为填埋场中垃圾往往处于不同的产气阶段,并且各地填埋场的温度条件存在很大差异,从而导致各地填埋场的产气速率存在较大差异。而对填埋场进行观测时,往往只能观测某一小段时间的填埋场表层气体排放情况,在这一小段时间里,填埋内部垃圾同时处于不同产气阶段,但相对处于较稳定状态,因而导致观测到的温度对填埋气排放量的影响有较大差异。
2.3 温度与不同阶段产气时间的关系
温度变化对垃圾发酵产气时间的影响相对比较明确,即随着温度的升高,可以显著加快垃圾发酵产气的过程,提高发酵产气效率。根据拟合曲线得出的产气曲线峰值及峰值时间如表3所示。
由表3可知,随温度升高,各组实验样品达到产气峰值所示时间迅速缩短,由此推断峰值期水解产酸阶段的菌种活跃度随温度升高而迅速升高;而各组实验样品的峰值高度随温度升高迅速升高也同样表明峰值期水解产酸阶段的菌种活跃度随温度升高而迅速升高。这一推论与此前由图7得出的结论相符,但略有差异,其原因推断为不同温度条件下,各个产气阶段产气时间在整个产气过程中所占的比例存在不同。
不同温度下,各产气阶段占总产气时间的比例如图11~图13所示。
如图11所示,虽然随着温度上升,峰值期的产气时间迅速缩短,但峰值期的产气时间在整个发酵产气过程中所占的比例各不相同。总体来看4个实验温度条件下,峰值期的产气时间在整个发酵产气过程中占30%左右;其中25℃条件下,峰值期的产气时间所占比例最小,仅为13%。结合该产气阶段累积产气量占总产气量比例来看,峰值期的产气时间比与累积产气比存在相关性,其中峰值期产气时间比,最小的25℃实验组峰值期累积产气比也最低,仅35%。而峰值期产气时间比略低于15℃及45℃的35℃实验组峰值期累积产气比略高于这两个实验组。结合之前温度与峰值期的累积产气量相关性的推论可以认为,15℃的低温条件下,微生物活动受抑制,水解产酸过程启动缓慢,因而15℃条件下峰值期所占时间最大;而在25℃条件下,虽水解产酸过程同样启动缓慢,但该温度条件下适宜甲烷菌的生长繁殖,因而发酵过程在水解产酸过程启动不久后很快进入稳定产甲烷阶段[6,15~17]。35℃及45℃的温度条件较适宜峰值期阶段的微生物活动,因而这两个温度条件下,峰值期的产气时间占总发酵过程时间较多,而35℃更接近峰值期的最佳产气温度,因而该阶段产气量占总产气量的比例也最大。
图12则更加验证了此前的推论,其中25℃条件下,稳定期的产气时间比在4个实验温度中最大,达到40.5%,并且在25℃条件下,稳定期累积产气量占总产气量的比值也远大于其他3个实验温度,达到49.9%;而15℃及45℃条件下,由于甲烷菌在过低及过高的温度中活动受抑制,因为在这两个温度条件下,稳定期的产气时间比较低,仅为21.4%和22.6%,而这两个温度条件下的稳定期累积产气量占总产气量的比值同样很低,仅为23.2%和23.3%;而35℃条件下,虽然其稳定期的产气时间比低于25℃条件,但也维持在较高水平,为35.9%,同时在这一温度条件下,其稳定期的累积产气量占总产气量的30%。
由图13可知,4个温度条件下,衰减期的产气时间占总发酵产气时间的40%左右,相差不大。其中25℃条件下,衰减期的产气时间占总发酵产气时间的比值最大,达46.5%;而35℃条件下,衰减期的产气时间占总发酵产气时间的比值最小,为38.5%;同时25℃及35℃条件下衰减期的累积产气量占总累积产气量的比值分别为衰减期累积产气比的最大值和最小值,分别为14.7%及4.2%。结合此前两个发酵阶段可知,而35℃条件下,峰值期水解产酸阶段以及稳定期的产甲烷阶段反应相对其他几个阶段更充分,实验样品中残留的可分解有机物含量较少,因而其衰减期产气时间较短,产气量相对其他3个温度条件也较少。
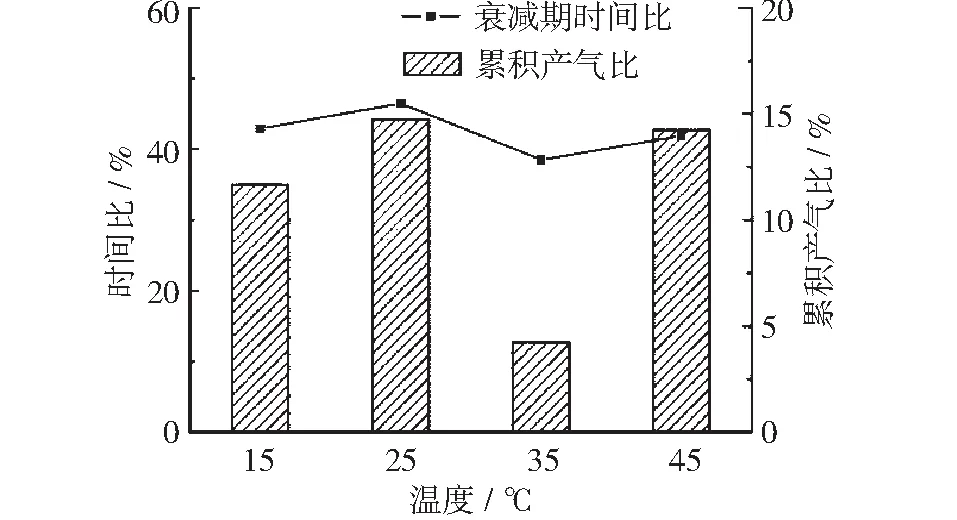
图13 产气衰减期时间比与累积产气比
通过分析不同温度条件下不同阶段的产气时间比及产气量比,更进一步验证此前的结论,即不同温度条件对不同产气阶段的产气时间,累积产气量均有不同影响。随温度升高,各实验组的总体发酵时间迅速减少,但产气总量总体上升。不同温度条件下,各产气阶段的产气时间占总产气时间的比例也存在较大差异,其中25℃条件下峰值期产气时间比最小,而稳定期产气时间比最长;并且在25℃条件下,其峰值期的累积产气量占总产气量的比值也最小,而稳定期的累积产气量占总产气量的比值也最大;可以认为在这一温度条件下,水解产酸菌种的活性与甲烷菌活性存在明显差异,这一温度条件对水解产酸菌种有明显抑制,但对甲烷菌活性有明显促进作用。
3 结论
(1)通过产气速率变化对垃圾产气曲线进行分段回归拟合的结果能准确反映实际产气变化,相关度高,拟合效果好。
(2)不同产气阶段受温度影响不同。稳定期产气量与温度关系最为明确,并且通过拟合数据发现31℃时稳定期产气量最大;峰值期产气量与温度关系需通过实验进一步探讨;衰减期产气量与温度相关性较弱,存在很大不确定性。
(3)不同产气阶段时间受温度影响同样不同。其中25℃条件下峰值期产气时间在总产气时间中比例最低为13%;稳定期产气时间在总产气时间中比例最高为40.5%;35℃条件下衰减期的产气时间在总产气时间中比例最小为38.5%。
(4)温度对不同产气阶段的影响集中体现在对不同产气阶段主要菌种活性的影响以及不同产气阶段产气时间的影响。
[1] 刘鸿霆,李启彬,刘 丹,等.垃圾填埋场甲烷释放速率的研究[J].安徽农业科学,2009,37(23):11151-11153.
[2] 杨 雪,岳 波,等.准好氧填埋场作业台阶表面CH4气体的释放通量研究[J].环境工程技术学报,2011,1(9):383-388.
[3] 马占云,李海玲,岳 波,等.生活垃圾填埋场覆盖层温室气体CH4和CO2释放规律及其相关性研究[J].环境工程技术学报,2014,4(5):399-405.
[4] Castrillon L. Anaerobic thermophilic treatment of cattlemanure in UASB reactors[J]. Waste Management Research, 2002, 20:350-356.
[5] Lier Van J B. New perspective in anaerobic digestion[J].Water Science and Technology, 2001, 43(1):1-18.
[6] 李 阳,周 涛,陈善平,等.温度对接种酵母菌和醋酸菌餐厨垃圾微氧发酵产乙酸的影响[J].中国环境科学,2016,36(1):175-180.
[7] 侯贵光,陈家军,张 娜,等.沼气产生的两步一级反应耦合模型[J].环境科学研究,2008,21(5):195-199.
[8] 赵宋敏,李定龙,戴肖云,等.温度对厨余垃圾厌氧发酵产酸的影响[J].环境污染与防治,2011,33(3):44-47.
[9] 王学东,王慧利,樊德方. 咪唑烟酸在土壤中的微生物降解及其代谢物分析[J].环境科学研究,2004,17(6):42-45.
[10] 马宗虎,冯小清,田 立,等.发酵浓度对餐厨垃圾厌氧消化特性的影响[J].中国沼气,2015,33(4):36-41.
[11] 文科军,杨 丽. 厨余有机垃圾产沼量的条件分析研究[J].环境科学与技术,2008,31(9):92-94.
[12] Y C Song, et al. Mesophilic and thermophilic temperature co-phase anaerobic digestion compared with single-stage mesophilic and thermophilic digestion of sewage sludge[J].Water Research, 2004,38:1653-1662.
[13] 吴满昌,孙可伟,李如燕,等.不同反应温度的城市生活垃圾厌氧发酵研究[J].化学与生物工程,2005,9:28-30.
[14] Ishigaki T, Yamada M, Nagamori M, et al. Estimation of methane emission from whole waste landfill site using correlation between flux and ground temperature[J].Environmental Geology, 2005, 48(7): 845-853.
[15] Melikoglu Mehmet, Lin Carol Sze Ki, Webb Colin. Analysing global food waste problem: pinpointing the facts and estimating the energy content[J].Central European Journal of Engineering, 2013,3(2):157-164.
[16] 焦刚珍,李 明,赵由才.不同来源污泥接种餐厨垃圾厌氧发酵产氢效果研究[J].环境污染与防治,2009,31(2):19-22.
[17] Li Yang, He Dongwei, NiuDongjie, et al. Acetic acid production from food wastes using yeast and acetic acid bacteria microaerobic fermentation[J].Bioprocess and Biosystems Engineering, 2015,38(5):863-869.
[18] Abichou T, Chanton J, Powelson D, et al. Methane flux and oxidation at two types of intermediate landfill covers[J].Waste Management, 2006, 26(11):1305-1312.
猜你喜欢
环境卫生工程(2021年5期)2021-11-20 05:45:24
环境卫生工程(2021年4期)2021-10-13 06:52:24
环境卫生工程(2020年3期)2020-07-27 01:19:18
Defence Technology(2019年6期)2020-01-07 09:11:04
中国当代医药(2019年20期)2019-09-25 05:03:40
农业机械学报(2019年1期)2019-02-15 07:30:26
中国美容医学(2016年4期)2016-07-25 06:16:52
中国石油大学学报(自然科学版)(2015年2期)2015-11-10 06:07:43
石油化工应用(2014年8期)2014-03-11 17:40:02
华西口腔医学杂志(2013年2期)2013-05-10 01:29:54
