
饲料铜水平对方格星虫稚虫生长的影响
2018-03-10 02:57:07许明珠董兰芳杨家林
水产科学 2018年1期
许明珠,童 潼,张 琴,董兰芳,蒋 艳,杨家林
( 广西壮族自治区海洋研究所,广西海洋生物技术重点实验室,广西 北海 536000 )

方格星虫(Sipunculusnudus)为北部湾特色经济海产品和近年来重点发展增养殖的经济动物之一,它生长快、病害少、活动范围小,味道鲜美,营养丰富,养殖面积和市场需求量逐年增大[7]。因此,研究有效促进方格星虫生长、适宜人工养殖的饲料,以提高方格星虫产量来满足日益增长的市场需求十分重要。目前有关方格星虫对饲料脂肪、蛋白、糖、硒、铁和锰的需求已有研究[8-13],而对饲料铜的研究却鲜见报道。本试验测定了由同一批受精卵孵出、规格相同、体质健康的方格星虫稚虫摄食含蛋氨酸铜饲料的质量日增加、组织铜含量和消化酶活性,研究了微量元素铜对方格星虫稚虫生长的影响,为研发规模化养殖方格星虫人工配合饲料提供参考。
1 材料与方法
1.1 试验饲料
蛋氨酸铜含铜量17%,由潍坊康科润生物技术有限公司友情提供,于基础饲料中分别添加蛋氨酸铜0、4、8、16、32 mg/kg和64 mg/kg(表1),制成6种试验饲料。经GB 13885—2003方法测定,饲料(以干物质计算)中铜的实际含量分别为2.7、7.5、11.2、19.8、30.4 mg/kg和52.1mg/kg。饲料制作参照文献[14]的方法进行,制成150目微颗粒饲料,标号装袋备用。
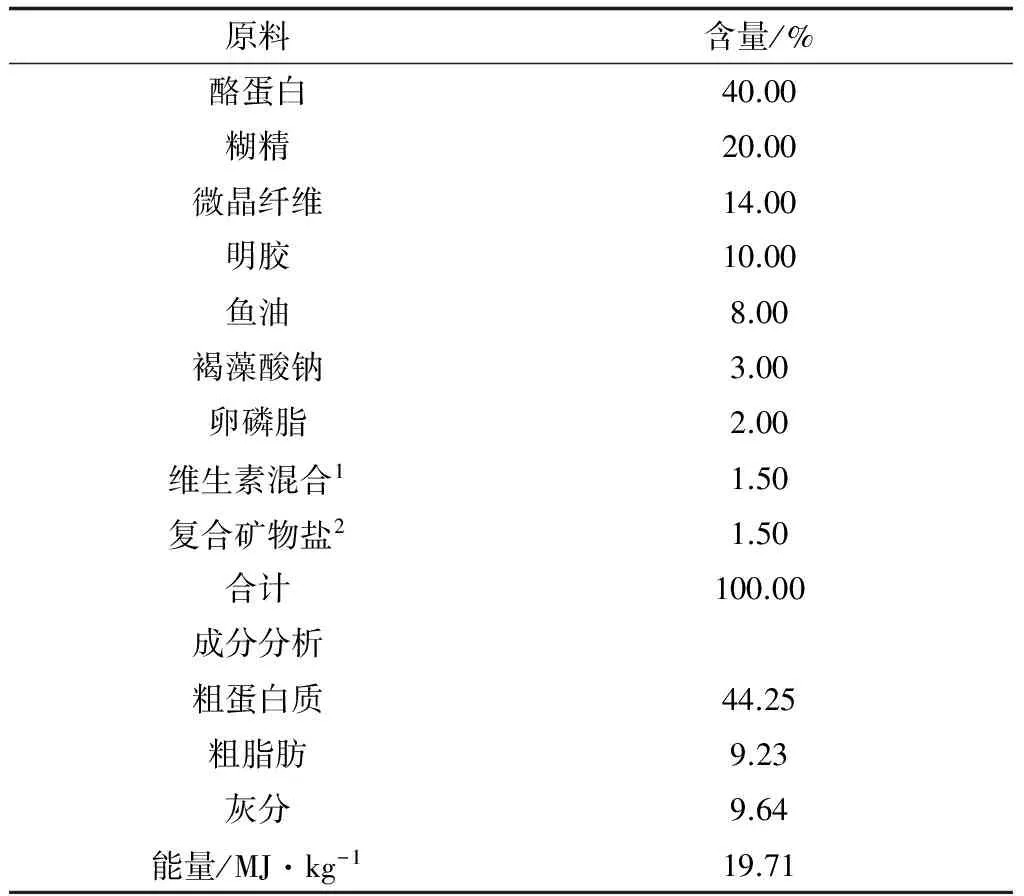
表1 试验饲料基础配方及营养成分分析(干物质)
注:1.每千克饲料维生素混合物提供:VD2400 IU,VE100 mg,VK1 mg,VC70 mg,VB10.5 mg,VB27 mg,VB66 mg,VB121 mg,D-泛酸32.6 mg,烟酸28 mg,生物素1 mg,叶酸7.5 mg,肌醇440 mg.2.每千克饲料矿物质预混料提供:CaCO33000 mg,ZnSO4·7H2O 300 mg,CaH2PO44125 mg,MnSO4·H2O 270 mg,NaCl 1200 mg,Na2SeO3·H2O 0.75 mg,KCl 750 mg,MgSO4·7H2O 1525.5 mg,FeSO4·7H2O 750 mg,AlSO4·6H2O 6 mg,CoCl2·6H2O 45 mg,KI 6 mg,沸石粉3021.75 mg.
1.2 试验设计与饲养管理
养殖试验于2013年8月至9月在广西海洋研究所海水增养殖实验基地进行。试验用稚虫取自广西海洋研究所培育的同批人工孵化苗种。正式试验前,先给试验稚虫投喂基础饲料14 d,使之逐渐适应试验饲料和养殖环境。驯养结束后,随机选出健康无病、活力较强、体质量相近[(18.46±0.11) mg]的稚星虫7200条,饲养于18个底层铺有约10 cm细沙的65 cm×55 cm×45 cm水槽中,每个水槽放养400条稚星虫。本试验采用自然光照,自然水温(26~30 ℃),二级过滤海水(盐度 18~22),24 h连续微充氧(溶解氧>5.0 mg/L),日换水两次(9:00和17:00),换水后过量投喂,保证沙子表面有少量未食用完全的饵料。养殖过程中每日巡视两次,及时清除和记录死亡的个体。
1.3 样品采集与指标测定
养殖结束后,将停食吐沙2 d的各组方格星虫收集,保存于-80 ℃超低温冰箱中。
平均终末体质量/mg=m/n
质量日增加/mg·d-1=(mt-m0)/t
式中,m和n分别为试验结束时每个水族箱中稚虫的总体质量(mg)和总数量(条),m0和mt分别为星稚虫平均初始体质量(mg)和平均终末体质量(mg),t为试验天数(d)。
饲料、虫体、体壁(解剖剪剪开后去除内脏和体腔液所得)均采用GB 13885—2003方法进行测定。将样品于550 ℃马福炉中灰化,用浓盐酸溶解残渣并稀释定容,然后导入原子吸收分光光度计(4510F,上海仪电分析)的空气—乙炔火焰中,测定铜元素的吸光度,并与元素校正溶液的吸光度比较定量。体腔液中铜含量采用化学法测定,试验原理为:在pH=9的氨溶液中,铜离子与二乙基二硫代氨基甲酸钠作用,生成摩尔比为1∶2的黄棕色胶体配合物,在452 nm处测定吸光值,标准曲线法定量铜离子的含量。
消化酶的活性的测定方法:参照南京建成生物工程研究所试剂盒的组织匀浆方法,将稚星虫整体匀浆进行酶液制备。蛋白酶活性采用Folin-酚法;脂肪酶活性和淀粉酶活性采用南京建成生物工程研究所试剂盒测定。以牛血清蛋白作标准,用考马斯亮蓝法测定酶液蛋白含量。酶活力用比活力(U/mg)表示。
本试验数据采用SPSS 19.0进行方差分析,若差异达到显著,则进行Tukey′s多重比较,显著性水平为0.05。
2 结果与分析
2.1 方格星虫摄食含不同铜水平饲料时的质量日增加
不同铜水平显著影响方格星虫稚虫的终末体质量和质量日增加(P<0.05),铜含量为11.2 mg/kg时终末体质量和质量日增加最高(0.68±0.02)mg/d,显著高于铜含量为2.7、19.8、30.4 mg/kg的试验组(P<0.05),但与铜含量为7.5、52.1 mg/kg的试验组差异不显著(P>0.05)(表2)。铜含量最低(2.7 mg/kg)的试验组稚虫的质量日增加显著低于其余各组(P<0.05)。
2.2 方格星虫摄食含不同铜水平饲料时组织的铜含量
饲料铜水平均显著影响方格星虫稚虫体中体壁铜含量和虫体铜含量(P<0.05)(表3)。稚星虫的体壁铜含量和虫体铜含量均随着饲料中铜含量的提高而升高。当饲料铜含量为52.1 mg/kg时,稚星虫的体壁铜含量和虫体铜含量均达到最大值,且其体壁铜含量显著高于较低(2.7 mg/kg)铜含量组(P<0.05);虫体铜含量显著高于2.7、7.5 mg/kg和11.2 mg/kg铜含量组。各处理组稚虫体腔液铜含量为2.32~4.53 μg/g,组间差异不显著(P>0.05)。

表2 方格星虫摄食含不同铜水平的饲料时的质量日增加(平均值土标准差)
注:同行数据肩标无字母或相同字母表示差异不显著(P>0.05),标不同小写字母表示差异显著(P<0.05).下同.
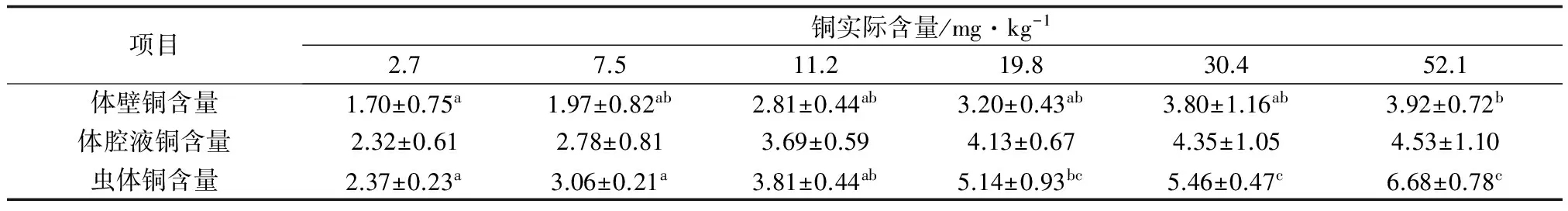
表3 饲料铜水平对方格星虫稚星虫组织铜的影响(平均值土标准差) μg/g
统计分析表明,体壁铜含量(y)与饲料铜含量(x)的回归方程为(图1):
y=0.8431ln(x)+0.683(r2=0.9351)
体腔液铜含量y1与饲料铜含量x1(2.7、7.5、11.2、19.8 mg/kg)的线性回归方程为(图2):
y1=0.1094x1+2.1031(r12=0.9138)
体腔液铜含量y2与饲料铜含量x2(19.8、30.4、52.1 mg/kg)的线性回归方程为(图2):
y2=0.0118x2+3.935(r22=0.9371)
由图2可知,当饲料铜含量(x)小于18.69 mg/kg时,体腔液铜含量(y)符合y1方程式;当饲料铜含量(x)大于18.69 mg/kg时,体腔液铜含量(y)符合y2方程式。
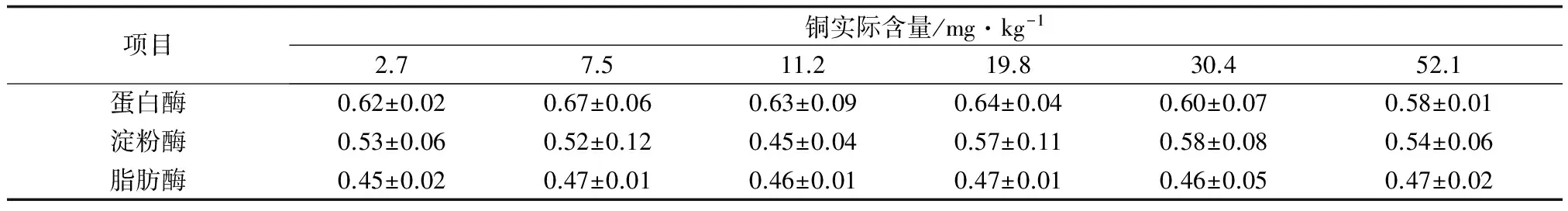
2.3 方格星虫稚虫摄食含不同铜水平饲料时消化酶的活性
饲料的铜水平对方格星虫稚虫的蛋白酶、淀粉酶、脂肪酶活性均无显著影响(P>0.05)(表4)。各处理组稚星虫蛋白酶活性为0.58~0.67 U/mg,淀粉酶活性为0.45~0.58 U/mg,脂肪酶活性为0.45~0.47 U/mg。

图1 方格星虫稚虫体壁铜含量与饲料铜含量的回归分析
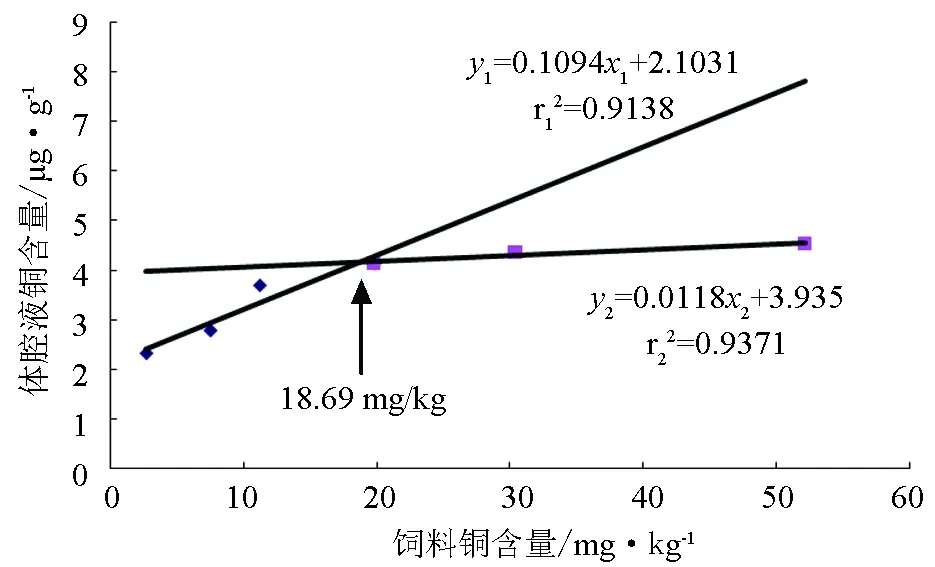
图2 方格星虫稚虫体腔液铜含量与饲料铜含量的回归分析
3 讨 论
3.1 饲料铜水平对方格星虫稚虫质量日增加的影响
已有研究表明,罗非鱼(Oreochromis)和草鱼(Ctenopharyngodonidellus)对饲料中谷氨酸铜[15]、斑节对虾(Penaeusmonodon)对饲料中蛋氨酸铜[16]的生物利用度大于硫酸铜;凡纳滨对虾(Litopenaeusvannamei)对饲料中各种铜源的生物利用度为蛋氨酸铜>富马酸铜>硫酸铜>碱式氯化铜[17-18];而异育银鲫(Carassiusauratusgibelio)对饲料中各种铜源的生物利用度则为碱式氯化铜>氨基酸铜络合物>硫酸铜[19]。
本试验中,在铜含量为11.2 mg/kg时,方格星虫的质量日增加最大,继续增加饲料铜的含量,稚星虫的质量日增加下降,然后缓慢上升趋于平稳。这与军曹鱼研究结果类似[23]。究其原因可能是方格星虫没有肝脏等器官,解毒能力较弱,铜元素和锌元素在生物体内具有拮抗作用,过度升高饲料中铜含量会影响生物对锌元素的吸收,因此高含量的饲料铜可能在一定程度上抑制方格星虫的生长[24-26]。
3.2 饲料铜水平对方格星虫稚虫体成分组织铜含量的影响
在其他水产动物的研究中,饲料中铜含量升高对凡纳滨对虾的肝胰脏中铜含量有显著影响,但对肌肉中铜含量均无显著影响;饲料铜水平为10、30、50 mg/kg时,硫酸铜和蛋氨酸铜为铜源极显著影响凡纳滨对虾肝胰脏的铜含量[27-28]。叶超霞等[29]研究高铜水平饲料的影响时发现,高铜水平饲料组的斜带石斑鱼(Epinepheluscoioides)全鱼和肝胰脏中的铜含量显著高于铜适量组。本研究中,饲料铜含量对方格星虫体壁铜含量和虫体铜含量有显著影响,对体腔液铜含量无显著影响,与上述研究结果略有出入。一般水产动物都将体内大部分铜储存于肌肉、骨骼和肝脏等器官组织中,方格星虫属于低等水生动物,体内并无骨骼和肝胰脏,其体内铜主要储存于体壁肌肉细胞和体腔液中,因此可能造成饲料铜含量的变化对体壁铜含量以及虫体铜含量影响显著[30],但是各试验组体腔液中铜元素含量差异不显著的原因还有待进一步研究。
4 结 论
试验结果表明,饲料铜含量变化显著影响方格星虫的质量日增加、体壁铜含量和虫体铜含量(P<0.05),但各组体腔液铜含量和消化酶活性差异不显著(P>0.05)。综合质量日增加和饲料成本,建议方格星虫稚虫饲料铜水平为7.5~11.2 mg/kg。
[1] 杨凤. 动物营养学[M].北京:中国农业出版社, 2009:84-102.
[2] 刘汉超, 叶元土, 蔡春芳, 等. 幼鱼阶段团头鲂对饲料中铜的需要量[J]. 动物营养学报, 2014, 26(11):3469-3477.
[3] Marc B, Anne-Kathrine L, Amund M. Effects of elevated dietary copper concentrations on growth, feed utilization and nutritional status of Atlantic salmon (SalmosalarL.) fry[J]. Aquaculture, 1999(174):167-181.
[5] 陈四清, 季文娟, 潘生弟. 黑鲷幼鱼对Zn、Cu的营养需要[J]. 中国水产科学, 1998, 5(2):52-56.
[6] Kanazawa A, Teshima S, Sasaki M. Requirements of the juvenile prawn for calcium, phosphorus, magnesium, potassium, copper, manganese, and iron[J].Memories of the Faculty of Fisheries, 1984(33):63-71.
[7] 李凤鲁,孔庆兰,史贵田,等.中国沿海方格星虫属(星虫动物门)的研究[J].青岛海洋大学学报, 1990, 20(1):93-99.
[8] 张琴, 童万平, 董兰芳, 等. 饲料中脂肪水平对方格星虫稚虫生长性能、体组成及消化酶活性的影响[J].渔业科学进展, 2011, 32(6):99-106.
[9] 张琴, 童万平, 董兰芳, 等. 饲料蛋白水平对方格星虫稚虫生长和体组成的影响[J]. 渔业科学进展, 2012, 33(1):86-92.
[10] 许明珠, 张琴, 童万平, 等. 饲料糖水平对方格星虫稚虫生长、体组成和消化酶活性的影响[J]. 动物营养学报, 2013, 25(3):534-542.
[11] 许明珠, 张琴, 童潼, 等.饲料中硒含量对方格星虫稚虫生长、体成分、组织硒含量及相关酶活性的影响[J]. 动物营养学报, 2015, 27(6):1733-1739.
[12] 许明珠, 张琴, 童潼, 等.饲料铁水平对方格星虫稚虫生长性能、体成分、酶活性及组织铁含量的影响[J].动物营养学报, 2014, 26(11):3325-3331.
[13] 许明珠, 张琴, 童潼, 等. 饲料中锰含量对方格星虫稚虫生长性能、体成分、体腔液中锰超氧化物歧化酶活性及组织锰含量的影响[J]. 动物营养学报, 2015, 27(10):3077-3083.
[14] Blair T, Castell J, Neil S, et al. Evaluation of microdiets versus live feeds on growth, survival and fatty acid composition of larval haddock (Melanogrammusaeglefinus)[J]. Aquaculture, 2003(225):451-461.
[15] 李云峰, 覃宗华, 彭险峰.谷氨酸铜对罗非鱼和草鱼生长性能血清生化指标和体组成的影响[J]. 中国饲料, 2013(18):34-36.
[16] 阳会军, 谭北平, 方怀义. 饲料中添加蛋氨酸铜和硫酸铜对斑节对虾生长和存活的影响[J]. 饲料工业, 2001(22):15-16.
[17] 郭志勋, 陈毕生, 徐力文, 等. 蛋氨酸铜和硫酸铜在凡纳滨对虾饲料中的应用效果比较[J]. 南方水产, 2005, 1(2):56-61.
[18] 周萌, 王安利, 曹俊明. 饲料中不同形式的铜及添加量对凡纳滨对虾(Penaeusvannamei)生长性能、血清铜蓝蛋白和生长激素水平的影响[J]. 海洋与湖沼, 2010, 41(4):577-582.
[19] Shao X P, Liu W B, Xu W N, et al. Effects of dietary copper sources and levels on performance, copper status, plasma antioxidant activities and relative copper bioavailability inCarassiusauratusgibelio[J]. Aquaculture, 2010,308(1/2):60-65.
[20] 乔永刚, 谭北平, 麦康森, 等. 军曹鱼对饲料中铜需要量的研究[J]. 中国海洋大学学报:自然科学版, 2013, 43(4):34-41.
[21] 赵宇江. 水产动物饲料中铜的营养与毒性的研究进展[J].饲料博览:技术版, 2009(1):28-31.
[22] 钱剑, 王哲, 刘国文. 铜在动物体内代谢的研究进展[J]. 动物医学进展, 2003, 24(2):55-57.
[23] 丁培君, 李丽, 邹本革. 铜源饲料添加剂研究进展[J]. 中国饲料添加剂, 2011(12):14-17.
[24] 吴鹏飞, 王嫣, 顾志峰, 等. 光裸方格星虫消化道的形态和组织学观察[J]. 渔业科学进展, 2010, 31(1):74-79。
[25] 廖伏初, 何望, 黄向荣, 等. 水产动物中铜的营养作用及研究进展[J]. 江西饲料, 2008(3):21-23.
[26] Hilton J W. Interrelationship between vitamins,inorganic salts and feed ingredients in feed of fishes[J].Aquaculture, 1989(79):223-244.
[27] 郭志勋, 陈毕生, 徐力文, 等. 饲料铜的添加量对南美白对虾生长、血液免疫因子及组织铜的影响[J]. 中国水产科学, 2003, 10(6):526-528.
[28] 董晓慧,杨原志,郑石轩,等.饲料中不同铜源和水平对凡纳滨对虾生长、免疫和组织铜含量的影响[J].大连水产学院学报, 2007,22(5):377-384.
[29] 叶超霞, 刘永坚, 田丽霞, 等. 饲料中高水平铜对斜带石斑鱼(Epinepheluscoioides)生长和铜、铁、锰、锌含量的影响[J]. 海洋与湖沼, 2013,44(3):606-610.
[30] 邓中日,黄勃,方再光.裸体方格星虫体壁形态和组织学结构[J].湖南城市学院学报:自然科学版, 2007, 16(1):62-66.
猜你喜欢
猪业科学(2021年3期)2021-05-21 02:05:42
南方农业学报(2020年10期)2020-01-21 15:36:41
中国临床医学影像杂志(2019年2期)2019-04-25 06:15:50
当代化工研究(2016年9期)2016-03-20 16:22:13
大连海洋大学学报(2015年2期)2016-01-26 07:44:38
焊接(2015年7期)2015-07-18 10:59:15
应用化工(2014年9期)2014-08-10 14:05:08
安徽农业科学(2014年3期)2014-04-29 16:58:15
故事作文·高年级(2009年11期)2009-12-11 10:24:24
故事作文·高年级(2009年9期)2009-12-11 10:24:18
