
3种砧木嫁接对柠檬幼树光合特性的影响
2018-01-30 06:30刘红明王绍华郑玉龙龙春瑞李进学付小猛沈正松高俊燕
经济林研究 2018年1期
刘红明,王绍华,郑玉龙,龙春瑞,李进学,付小猛,沈正松,高俊燕
(1.云南省农业科学院热带亚热带经济作物研究所,云南 保山 678000;2.大理大学 农学与生物科学学院,云南 大理 671003)
砧木作为接穗的营养运输通道,对矿质养分的运输起着极其重要的作用,对果树树体生长、果实成熟期、产量和经济性状均有重要影响[1],还可提高嫁接品种的抗逆性和适应性[2-4]。同时,果树的“砧—穗”组合还会影响植株呼吸、矿质营养吸收和光合等生理生化过程[5-6]。这已在柑橘[7-9]、桃树[10]、苹果[11]、梨[12]和猕猴桃[13]等果树上得到证实。砧木对嫁接果树品种的影响是一个受内外因素综合影响且非常复杂的生理生化过程[14-15],目前多采用田间调查嫁接植株生长数据来研究及推测砧木对接穗的作用[16-17],较少涉及砧木对嫁接品种光合性能和叶片光合色素含量影响的研究。一般认为,果树叶片光合性能与其生产潜力呈正相关[18]。
目前在柠檬生产上应用最多的砧木是枳壳,枳壳砧在生长后期表现嫁接不亲和、不耐土壤瘠薄,在生产中还存在着抗逆性(抗病性和抗旱性)差等缺点,成为制约柠檬增产和品质改善的重要因素[19-20]。因此选育新型优良砧木刻不容缓。但柠檬砧穗互作机制尚缺乏系统研究,在光合特性方面的研究尤为缺乏,而这正是评价其生态适应性的基础[21-22]。目前,关于柠檬砧木的研究主要集中在树体生长和营养代谢等领域[23-25],专家学者开展不同砧木对樱桃[26]、核桃[27]、苹果[28]和李[29]等果树光合特性影响的研究较多。但不同砧木嫁接对柠檬光合特性影响的研究鲜见报道。本试验选用3种砧木,对不同砧木嫁接云柠1号柠檬和4号柠檬的光合特性进行研究。测定3种不同砧木嫁接柠檬的光合日变化和叶绿素相对含量,从光合生理的角度比较不同砧木对柠檬光合性能的影响,旨在为筛选优良柠檬嫁接砧木提供参考。
1 材料与方法
1.1 试验地概况
试验于 2016 年在云南省农科院热带亚热带经济作物研究所德宏州瑞丽市科研基地进行,试验柠檬品种和砧木见表1。试验前,取本地耕层(0~30 cm)的土壤,去除杂质后粉碎,混合均匀,取样进行测定分析,其理化性质为:有机质含量为2.24 g/kg、pH值为5.72、全氮含量为0.256 g/kg、全磷含量为0.098 g/kg、全钾含量为1.824 g/kg、碱解氮含量为69.26 mg/kg、速效磷含量为26.47 mg/kg、速效钾含量为208.42 mg/kg、交换性镁含量为197.46 mg/kg、有效锰含量为20.47 mg/kg、有效铁含量为57.66 mg/kg、有效锌含量为1.11 mg/kg、有效铜含量为2.29 mg/kg、有效硼含量为0.23 mg/kg。所属区域为南亚热带季风气候类型,最近10 a平均气温 18.4~21.0 ℃,最高气温和最低气温分别为38.8、-2.1 ℃,年积温6 400~7 300 ℃;年日照2 281~2 453 h;年降雨量1 400~1 700 mm。

表1 试验材料Table1 Test materials
1.2 试验设计
试验采用 LCpro-SD 全自动便携式光合测定仪(英国ADC公司生产),于2016年9—11月分别对不同砧木嫁接柠檬幼树光合作用日变化进行田间测定。测定时选取树冠中上部南面的当年成熟叶片,叶龄相对一致。选择晴朗无云天气,从 8:00—18:00 每隔2h 测定1次,每个品种测定3 株树,每株5片叶。主要测量指标包括:净光合速率(Pn)、气孔导度(Gs)、胞间 CO2浓度(Ci)、蒸腾速率(Tr)、叶面温度(Tl)、光合有效辐射(Pa)等。测定时每片叶片重复测定3次,取其平均值。采用植物营养速测仪TYS-4N(浙江托普公司生产)测定叶片叶绿素相对含量(SPAD值)和叶片氮素含量,测定叶片的前中后3个不同的位置,计算其平均值,每个处理测6株树,每棵树重复3次。
1.3 数据处理
水分利用效率由计算公式Wu=Pn/Tr计算得到,光能利用效率:Lu(%)=Pn·100/Pa,叶肉瞬时羧化效率:Ce=Pn/Ci。采用Microsoft Excel 2013 和 SPSS 13.0 进行数据统计及相关性分析。
2 结果与分析
2.1 光合有效辐射日变化情况
由图1可知,光合有效辐射(Pa)整体出现先上升后下降的倒 “U” 型图,Pa在138~1 969 μmol·m-2s-1之间波动,在 8:00—14:00 之间上升,14:00到达峰值,之后逐渐下降。
2.2 叶面温度与外界温度光合日变化比较

图1 光合有效辐射日变化Fig.1 Diurnal change of photosynthetically active radiation (Pa)
相关分析表明,叶面温度(Tl)与外界温度均随光合有效辐射的变化而变化,二者显著正相关。由图2可以看出,叶面温度始终高于外界温度,二者温度在8:00—14:00之间持续上升,并在14:00 达到峰值,随后温度逐渐降低。

图2 叶面温度与外界温度日变化比较Fig.2 Comparison of diurnal changes of leaf surface temperatures (Tl) and external temperatures (Tc)
2.3 不同砧木嫁接柠檬胞间 CO2 浓度日变化比较
由图3可以看出,胞间CO2浓度(Ci)在8:00—10:00 之间持续下降,Ci在 229 ~ 439 μmol·mol-1之间变化,10:00之后3种砧木嫁接柠檬Ci下降趋势明显降低。4号NM/SY在10:00—14:00时之间略微上升,其变化范围在0~6 μmol·mol-1之间,其余类型在10:00—12:00均呈现下降。12:00—14:00YN1/SY、YN1/YDLM呈现上升,在该时间段4号NM/YDLM迅速降低,并在14:00达到谷值。14:00—16:00时4号NM/YDLM出现第一次上升,在 16:00 达到峰值 284 μmol·mol-1,其余砧木嫁接柠檬在此期间逐渐下降。16:00之后不同砧木嫁接的柠檬胞间CO2浓度又逐渐升高。
2.4 不同砧木嫁接柠檬气孔导度日变化比较

图3 3种砧木嫁接的柠檬胞间CO2浓度日变化比较Fig.3 Comparison of diurnal changes of intercellular CO2 concentrations (Ci) in lemon grafted with three kinds of rootstocks

图4 3种砧木嫁接的柠檬气孔导度日变化比较Fig.4 Comparison of diurnal changes of stomatal conductances (Gs) in lemon grafted with three kinds of rootstocks
由图4可知,气孔导度(Gs)在10:00时达到全天最高,且在该时刻枳壳嫁接柠檬高于另外2种砧木嫁接。在10:00—14:00之间气孔导度降低,这是由于外界温度上升导致气孔闭合以减少蒸腾作用的结果。气孔导度在16:00时出现第二个峰值,可以看出气孔导度日变化与净光合速率曲线均呈现“双峰”型,在光合“午休”期间,气孔导度降至最低。可以看出枳壳嫁接柠檬在8:00—12:00之间气孔导度相对较高,酸柚嫁接柠檬气孔导度处于相对最低值。14:00时之后印度莱檬嫁接4号柠檬气孔导度大于其它几种砧木嫁接,枳壳砧木嫁接柠檬除了比印度莱檬嫁接4号柠檬气孔导度低以外,均高于其余砧木。
2.5 不同砧木嫁接柠檬净光合速率日变化比较
由图5可以发现,不同砧木嫁接柠檬净光合速率日变化为典型的“双峰”型,第一个峰值出现在10:00左右,其最大值从大到小依次为:YN1号/ZK>4号NM/ZK>4号NM/SY> YN1号/SY>YN1号/YDLM>4号NM/YDLM,最大净光合速率在11.08~13.14 mol·m-2s-1之间变化。第二个峰值出现在16:00左右;第一个峰值之后,随着外界温度和叶面温度的升高,不同砧木嫁接的柠檬净光合速率逐渐降低,并在14:00左右出现光合“午休”现象。从该图中还可以看出,在光合“午休”期间,用枳壳砧木嫁接的4号柠檬、云柠1号净光合速率仍大于其余砧木嫁接的柠檬。

图5 3种砧木嫁接的柠檬净光合速率日变化比较Fig.5 Comparison of diurnal changes of net photosynthetic rates (Pn) in lemon grafted with three kinds of rootstocks
2.6 不同砧木嫁接柠檬蒸腾速率日变化比较
由图6可以发现,3种砧木嫁接云柠1号蒸腾速率曲线呈现倒“U”型,而3种砧木嫁接4号柠檬蒸腾速率曲线呈现“双峰”型。在3种砧木嫁接云柠1号和4号柠檬中,8:00—12:00,枳壳砧木嫁接的柠檬蒸腾速率相对最低。在14:00时光合“午休”期间,蒸腾速率从大到小依次为:印度莱蒙>枳壳>酸柚,16:00左右枳壳砧木嫁接柠檬蒸腾速率均处于相对最低。
2.7 不同砧木嫁接柠檬水分利用率、光能利用率、叶肉瞬时羧化效率日变化比较
从图7可见,3种砧木嫁接柠檬光能利用率在8:00—10:00之间快速下降,直到光合午休时达最低值,之后缓慢回升。在一天中光合速率最高时(10:00)4号柠檬和云柠1号嫁接枳壳的光能利用率最高。
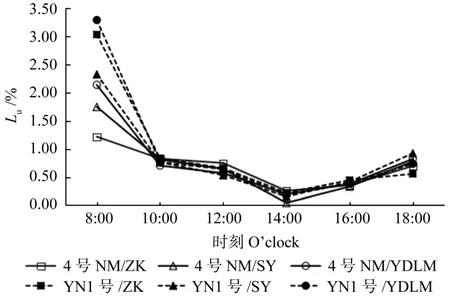
图7 3种砧木嫁接的柠檬光能利用率日变化比较Fig.7 Comparison of diurnal changes of light use eff i ciency (Lu) in lemon grafted with three kinds of rootstocks
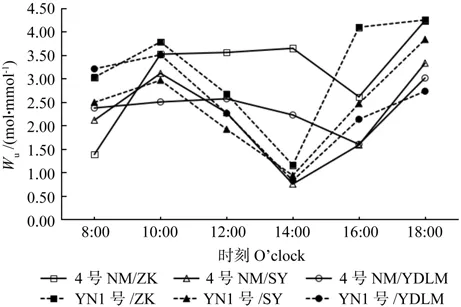
图8 3种砧木嫁接的柠檬水分利用率日变化比较Fig.8 Comparison of diurnal changes of water use eff i ciency (Wu) in lemon grafted with three kinds of rootstocks
从图8可以看出,3种不同砧木嫁接的柠檬在10:00左右出现一个水分利用率的高峰,其中云柠1号嫁接枳壳的水分利用率最高,达3.78%;随后逐渐下降,到中午光合午休时(14:00)达到一天中的最低值,其中4号柠檬嫁接枳壳和印度莱檬例外,在16:00前呈一条平缓曲线,到16:00时才达到一天中的最低值。由图9可以发现,不同砧木嫁接柠檬叶肉瞬时羧化效率随时间变化曲线和净光合速率变化曲线趋势一致,均为典型的“双峰”型,第一和第二峰值分别出现在10:00和16:00,但和净光合速率不一样的是,第一峰值叶肉瞬时羧化效率最高的是云柠1号嫁接印度莱檬;所有砧木嫁接柠檬在光合午休期间(14:00)叶肉瞬时羧化效率均达到最低值,其中4号柠檬嫁接酸柚在所有处理中最低。
2.8 不同砧木嫁接柠檬叶片SPAD值及氮含量比较
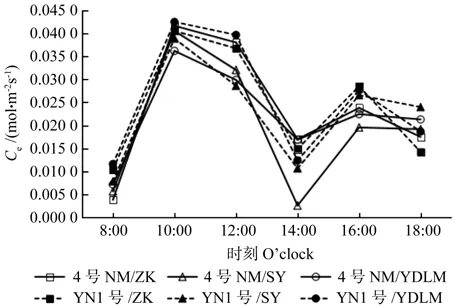
图9 3种砧木嫁接的柠檬叶肉瞬时羧化效率日变化比较Fig.9 Comparison of diurnal changes of instantaneous carboxylation eff i ciency (Ce) in lemon grafted with three kinds of rootstocks
不同砧木嫁接柠檬叶片SPAD值及氮含量见表2。从表2可以看出,枳砧/云柠1号SPAD值和叶片氮含量均为最高,和嫁接在印度莱檬上相比差异显著,与嫁接在酸柚上相比差异则不显著;4号柠檬嫁接在枳壳与嫁接在酸柚上的SPAD值和叶片氮含量相比,差异不显著,与嫁接在印度莱檬上相比差异显著。4号柠檬和云柠1号分别嫁接在枳壳和酸柚上的SPAD值和叶片氮含量相互间差异不显著,仅4号柠檬和云柠1号分别嫁接在印度莱檬上相比差异显著。
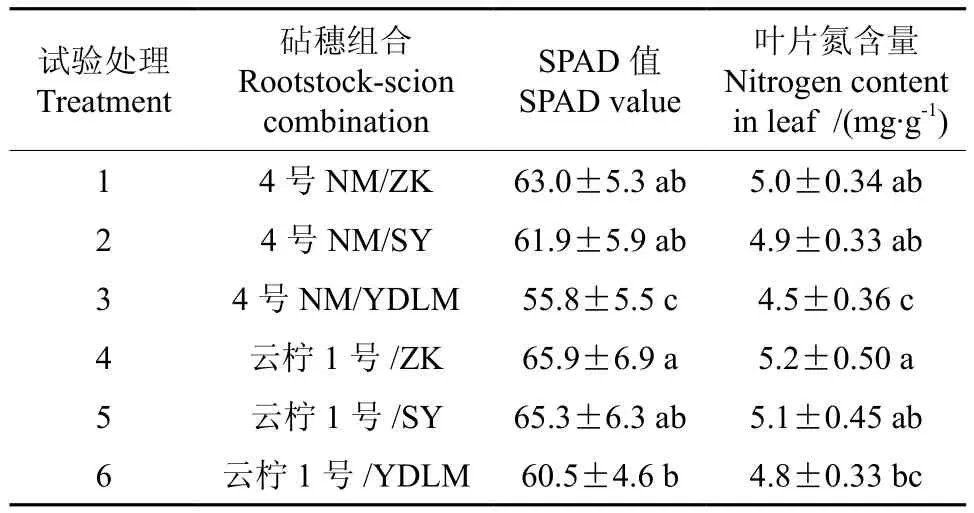
表2 不同砧木嫁接柠檬叶片SPAD值及氮含量比较Table2 Comparison of SPAD values and nitrogen contents of grafted lemon leaves with different rootstocks
3 讨论与结论
果树砧木对嫁接品种光合性能有较大影响,尤其是对光合速率的影响最大[30]。张建光等[11]的研究表明:M26和M72种苹果砧木对供试品种叶片的光合性能显著高于Bud2。赵红军等[31]对不同砧木香玲核桃的光合特性的研究表明:黑核桃砧香玲核桃叶片的最大光合速率显著高于本砧核桃,其主要原因是黑核桃砧接穗叶片的叶绿素a、叶绿素b和总叶绿素含量显著高于本砧核桃。刘凤芹[32]研究发现,不同砧穗组合间南果梨叶片的气孔导度、胞间 CO2浓度均达到显著差异。在温州蜜柑上的研究表明,枳砧净光合速率(Pn)、蒸腾速率(Tr)、水分利用率(Wu) 等均为最高,光合性能表现最优[8],这与本研究结果一致;廖玲等[33]对黄果柑的研究表明,在供试的砧木中,香橙砧具有最佳的光合性能,不同种类砧木嫁接的黄果柑净光合速率均表现为“双峰”型;而在脐橙上的研究则表明,不同砧木嫁接的脐橙间净光合速率差异显著,其光合速率日变化有“单峰”、“双峰”、“多峰”等多种曲线类型[34]。本试验中,3种砧木对柠檬幼树光合性能的影响较大,其中枳砧/云柠1号柠檬的光合性能最好,其最大净光合速率(Pmax)、气孔导度(Gs) 、水分利用效率(Wu)、光能利用效率(Lu)和叶片叶绿素含量(SPAD值)均最高。嫁接的柠檬幼树净光合速率均表现为“双峰”型曲线,具有明显的光合“午休”现象,这可能是由中午光合辐射过强、温度过高导致气孔导度和胞间 CO2浓度急速下降引起。这和胡利明与何海洋等人[8,35]对枳砧温州蜜柑幼树的研究结果一致。
叶绿素是绿色植物叶片中参与光合作用的主要色素,它参与光能的吸收、传递和转化,是植物光合作用的基础。叶片的光合速率与叶绿素含量高低密切相关[36],植株功能叶中叶绿素含量的高低在很大程度上反映了植株的光合性能和营养状况。本研究中,具有最大净光合速率的枳砧/云柠1号也同样具有最高的SPAD值和叶片氮含量,而在琯溪蜜柚上的研究显示,叶绿素含量与Pn间相关性不明显[37],原因可能与其选取的特殊测定时间(采果后)有关。
本研究表明,不同砧穗组合的光合性能有较大差异,这些差异必然会导致生产性能的不同。由于果树是多年生植物,育苗和建园时必须认真考虑选择适宜的砧穗组合。就本试验而言,枳砧/云柠1号组合具有最高的净光合速率、最高的SPAD值和叶片氮含量,同样具有最高的光能利用率和水分利用率,叶肉瞬时羧化效率也较高,若仅从叶片光合性能和叶片叶绿素含量考虑,枳砧/云柠1号为最适砧穗组合。但影响砧木对接穗品种光合特性的因素较多,除了砧木本身的遗传因素外,还受树体栽培环境等外界因素的影响。因此,有关砧木影响柠檬光合作用的机理尚需作进一步的系统研究。此外,生产上评价砧木的指标较多,除光合性能外,生长和结果特性、抗逆性和果实品质也是重要的评价指标,本研究仅研究了砧穗组合的光合特性,下一步计划将生长特性和结果特性与光合性能结合起来研究,为全面、系统地评价筛选柠檬砧木提供科学依据。
[1] 符泽华, 段伟华, 渠心静, 等. 不同品种接穗对油茶高接换冠后产量及经济性状的影响[J]. 中南林业科技大学学报, 2017,37(10): 75-79.
[2] 李小红, 周 凯, 谢 周, 等. 不同葡萄砧木对矢富罗莎葡萄嫁接苗光合作用的影响[J]. 果树学报, 2009, 26(1): 90-93.
[3] Smith M W, Shaw R G, Chapman J C, et al. Long-term performance of ‘Ellendale’ mandarin on seven com-mercial rootstocks in subtropical Australia[J]. Scientia Horticulturae,2004, 102(1): 75-89.
[4] Garciasanchez F, Perezperez J G, Botia P, et al. The response of young mandarin trees grown under saline conditions depends on the rootstock[J]. European Journal of Agronomy, 2006, 24(2):129-139.
[5] 朱丽华, 章文才. 砧木对柑桔嫁接幼树早果影响的生理生化研究[J]. 园艺学报, 1991, 18(4): 296-302.
[6] 郁万文, 曹福亮, 汪贵斌, 等. 不同接穗无性系对银杏嫁接树生长影响及材用无性系初选[J].中南林业科技大学学报,2016, 36(3): 1-7.
[7] Morinaga K, Ikeda F. The effects of several rootstocks on photosynthesis, distribution of photosynthetic product, and growth of young Satsuma mandarin trees[J]. Engei Gakkai Zasshi, 1990, 59 (1): 29-34.
[8] 胡利明, 夏仁学, 周开兵, 等. 不同砧木对温州蜜柑光合特性的影响[J]. 园艺学报, 2006, 33(5): 937-941.
[9] Jover S, Martinez-Alcantara B, Rodriguez-Gamir J. Inf l uence of rootstocks on photosynthesis in navel orange leaves: effects on growth, yield, and carbohydrate distribution[J]. Crop Science,2012, 52 (2):836-848.
[10] Tsipouridis C, Thomidis T. Effect of14peach rootstocks on the yield, fruit quality, mortality, girth expansion and resistance to frost damages of May Crest peach variety and their susceptibility on Phytophthora citrophthora[J]. Scientia Horticulturae, 2005,103(4): 421-428.
[11] 张建光, 刘玉芳, 施瑞德. 不同砧木上苹果品种光合特性比较研究[J]. 河北农业大学学报, 2004, 27(5): 31-33.
[12] 姜卫兵, 俞开锦, 高光林, 等. 梨不同砧穗组合光合特性[J].园艺学报, 2002, 29(6): 569-570.
[13] 龚弘娟, 叶开玉, 蒋桥生, 等. 4种砧木嫁接的红阳猕猴桃光合特性比较[J]. 南方农业学报, 2014, 45(10): 1825-1830.
[14] 吕 斌, 陈学年. 不同砧木先锋橙叶片POD活性与树体生长势的关系[J]. 西南农业学报, 1999, 12(2): 63-67.
[15] 欧 毅, 毛启才, 曹照春, 等. 不同砧木锦橙叶片过氧化物酶活性与植株生长结果的关系[J]. 云南农业大学学报, 1994,9(1): 23-27.
[16] 赵同生. 毛樱桃与李的嫁接亲合性及致矮效果[J]. 河北果树,2003(4): 12-12.
[17] 周怀军, 张洪武, 张晓曼, 等. 不同砧木IAA酶活性与李树树体生长势的关系[J]. 河北林业科技, 2002(6): 1-3.
[18] 马玉坤, 贾永芳, 马静芳, 等. 不同砧木对红地球葡萄光合日变化的影响[J]. 江苏农业学报, 2012, 28(3): 638-642.
[19] 周开兵, 郭文武, 夏仁学, 等. 两种柑橘体细胞杂种砧木利用价值和砧穗互作生化机制的探讨[J]. 园艺学报, 2004, 31(4):427-432.
[20] 周开兵, 夏仁学.中国柑橘砧木选择研究进展与展望[J]. 中国农学通报, 2005, 21(1): 213-217.
[21] Huang W D, Wu L K , Zhan J C. Effect of weak light on the peroxidation of membrane-lipid of cherry leaves[J]. Acta Botanica Sinica, 2002, 44(8): 920-924.
[22] Wu L K, Huang W D, Zhan J C. Effects of low light intensity on sweet cherry fruit setting and quality[J]. Journal of China Agricultural University, 2002, 7(3): 69-74.
[23] 周东果, 董美超, 李进学, 等. 嫁接不同柠檬接穗对植株生长的影响[J]. 江苏农业科学,2014, 42(7): 151-154.
[24] 洪 林, 谢永红, 刘正富. 砧木对菲诺47柠檬树体生长、果实品质及叶片氮磷钾含量的影响[J].南方农业学报, 2012,43(8): 1173-1178.
[25] 胡 敏, 兰 翔, 何玉广, 等. 不同砧木对云柠1号柠檬叶片养分含量的影响[J]. 湖北农业科学, 2014, 53(13): 3066-3069.
[26] 秦嗣军, 吕德国, 杜国栋, 等. 5种樱桃砧木光合特性的初步研究[J]. 园艺学报, 2006, 33(4): 813-816.
[27] 孟丙南, 张俊佩, 裴 东, 等. 不同砧木对核桃光合特性的影响[J]. 经济林研究, 2013, 31(2): 32-37.
[28] 宋 莎, 江旭生, 韩秀梅, 等.不同苹果砧木叶绿素含量与光合速率研究[J]. 江苏农业科学, 2015, 43(3): 149-150.
[29] 李欧毅, 王 进, 谢永红, 等. 不同砧木对李叶片光合效能和生理生化指标的影响[J]. 西南农业大学学报(自然科学版),2006, 28(3): 428-431.
[30] 赵登超, 侯立群, 韩传明, 等.我国核桃光合作用研究进展[J].山东林业科技, 2011, 41(4): 107-111.
[31] 赵红军, 魏海蓉, 姜 涛, 等. 黑核桃砧和本砧香铃核桃的光合特性初探[J].山东农业科学, 2007(3): 53-55.
[32] 刘凤芹, 王春枝, 刘凤杰, 等. 不同砧穗组合对南果梨光合特性的影响[J].安徽农业科学, 2010, 38(31): 17373-17374.
[33] 廖 玲, 曹淑燕, 荣 毅, 等.不同砧木对黄果柑光合特性、光合作用关键酶及其基因表达的影响[J]. 浙江农业学报,2016, 28(5): 769-775.
[34] 梅正敏, 罗世杏, 伊华林, 等. 不同砧木嫁接对桂脐1号脐橙幼树生长及光合特性的影响[J]. 南方农业学报, 2014, 45(3):434-441.
[35] 何海洋, 彭方仁, 张 瑞, 等. 不同品种美国山核桃嫁接苗光合特性比较[J].南京林业大学学报(自然科学版), 2015,39(4): 19-25.
[36] 赵 平, 曾小平, 彭少麟, 等.海南红豆(Ormosia pinnata)夏季叶片气体交换、气孔导度和水分利用效率的日变化[J]. 热带亚热带植物学报, 2000, 8(1): 35-42.
[37] 易干军, 姜小文, 霍合强, 等. 琯溪蜜柚光合特性的研究[J].园艺学报, 2003, 30(5): 519-524.
猜你喜欢
农业工程学报(2022年3期)2022-04-16
生态学报(2021年6期)2021-04-25
农业工程学报(2021年3期)2021-04-15
节水灌溉(2020年9期)2020-09-16
农业与技术(2020年12期)2020-07-04
农学学报(2019年1期)2019-02-21
现代园艺(2017年19期)2018-01-19
新课程·下旬(2017年7期)2017-08-14
中成药(2017年6期)2017-06-13
发明与创新(2016年5期)2016-08-21
