
江蓠、浒苔、藻渣和菌渣替代鱼粉对大菱鲆幼鱼生长性能、血清和肝脏生化指标、体组成和肠道组织结构的影响
2018-01-23 06:21梁萌青徐后国卫育良
动物营养学报 2018年1期
郭 斌 梁萌青 徐后国 卫育良
(1.中国水产科学研究院黄海水产研究所,青岛 266071;2.上海海洋大学水产与生命学院,上海 201306)
近年来,全球水产养殖产量增长迅速,养殖规模的扩大增加了对水产饲料的需求。鱼粉是水产饲料中蛋白质的主要来源,但近年由于过度捕捞、环境污染及厄尔尼诺现象等不良气候的影响,鱼粉资源日益减少,鱼粉价格不断上涨[1]。因此,对于水产饲料行业,节约鱼粉的用量并开发合适的蛋白质替代源显得极为重要。
植物蛋白质因其来源广、产量稳定、价格低廉等优点,一直作为替代鱼粉的蛋白质来源,传统的植物蛋白质虽然在水产饲料中的应用研究已有很多[2-5],但由于植物蛋白质含有抗营养因子[6],并缺乏鱼类所需的一些必需氨基酸和促生长成分,因此不能完全替代鱼粉。海藻是生活在海洋中含有叶绿素的一类自养植物,与陆生植物相比,海藻含有丰富的矿物质、维生素、海藻多糖、不饱和脂肪酸、游离氨基酸以及具有诱食作用的一些物质,可以补充陆生植物蛋白质大量替代鱼粉所造成的营养缺乏[7-8]。我国海域辽阔,海藻资源十分丰富,但这些海藻资源还未充分利用,如果可以充分利用海藻中的蛋白质资源,对于解决水产饲料的蛋白质来源有着重要意义。因此,本研究以常见的2种低值海藻(江蓠、浒苔)和2种工业下脚料(海带提取褐藻胶后的藻渣、土曲霉发酵生产衣康酸后的菌渣)与植物蛋白质配合成混合植物蛋白质替代大菱鲆幼鱼饲料中部分鱼粉,探究其对大菱鲆幼鱼生长性能、血清和肝脏生化指标、体组成和肠道组织结构的影响,以期为海藻及工业下脚料在大菱鲆饲料中的应用提供参考。
1 材料与方法
1.1 试验饲料
试验共配制5种等氮等脂的试验饲料,首先以鱼粉为主要蛋白质源、鱼油为脂肪源配制含60%鱼粉基础饲料(鱼粉组,作为对照组),然后分别将10%的江蓠、浒苔、藻渣(海带提取褐藻胶后的废渣)、菌渣(土曲霉发酵生产衣康酸后的废渣)与植物蛋白质谷朊粉、玉米蛋白粉、豆粕配合后替代基础饲料中35%的鱼粉。所有原料测定常规营养成分后,粉碎机粉碎,过80目筛网,按配方称量原料,逐级混匀,然后加鱼油混匀,加30%水搅拌均匀,用制粒机制成直径为2 mm的颗粒饲料,55 ℃鼓风干燥12 h后置于-20 ℃冷库保存。江蓠、浒苔、藻渣和菌渣的常规营养成分见表1,试验饲料组成及营养水平见表2,试验饲料氨基酸组成见表3。
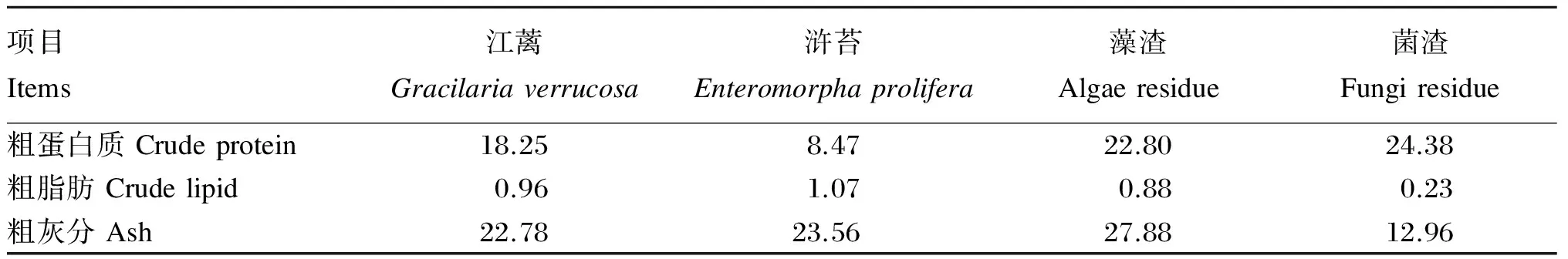
表1 江蓠、浒苔、藻渣和菌渣的常规营养成分(干物质基础)
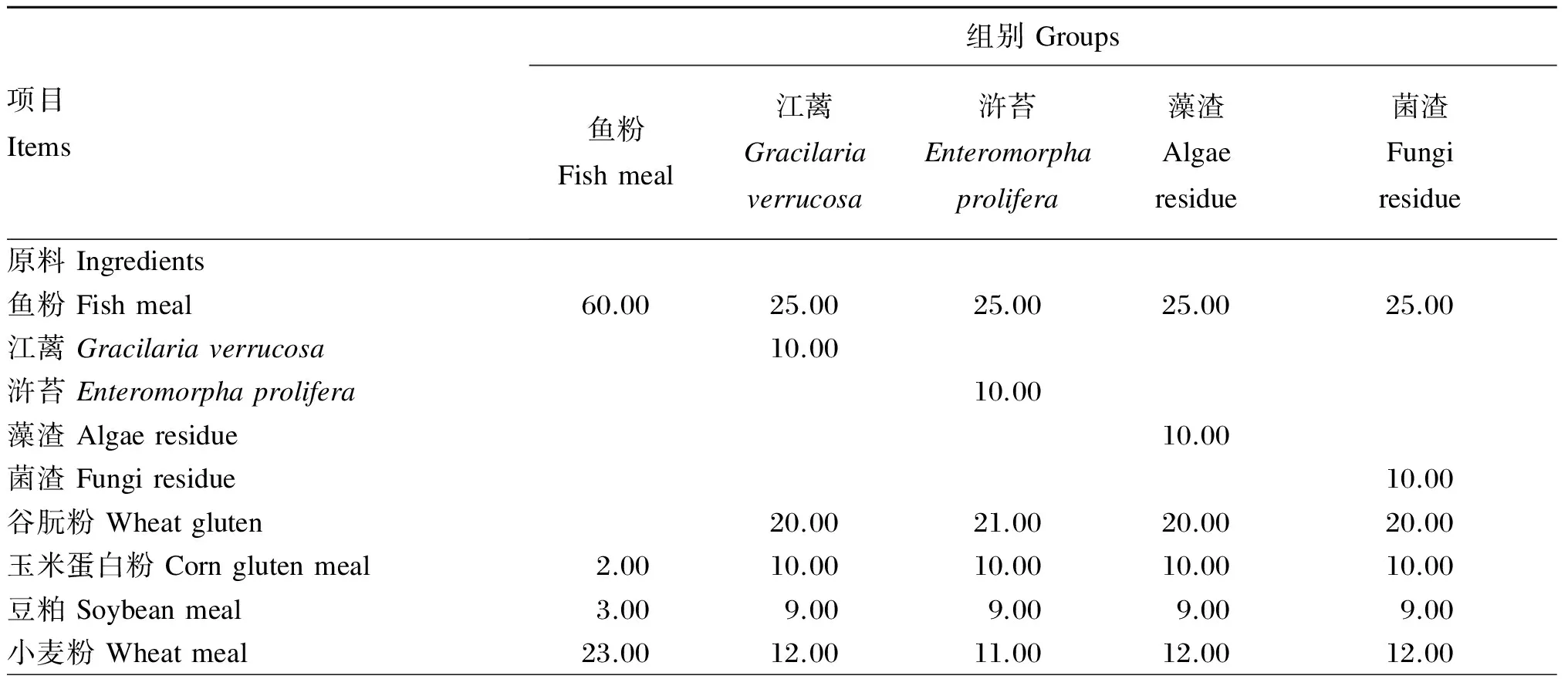
表2 试验饲料组成及营养水平(干物质基础)
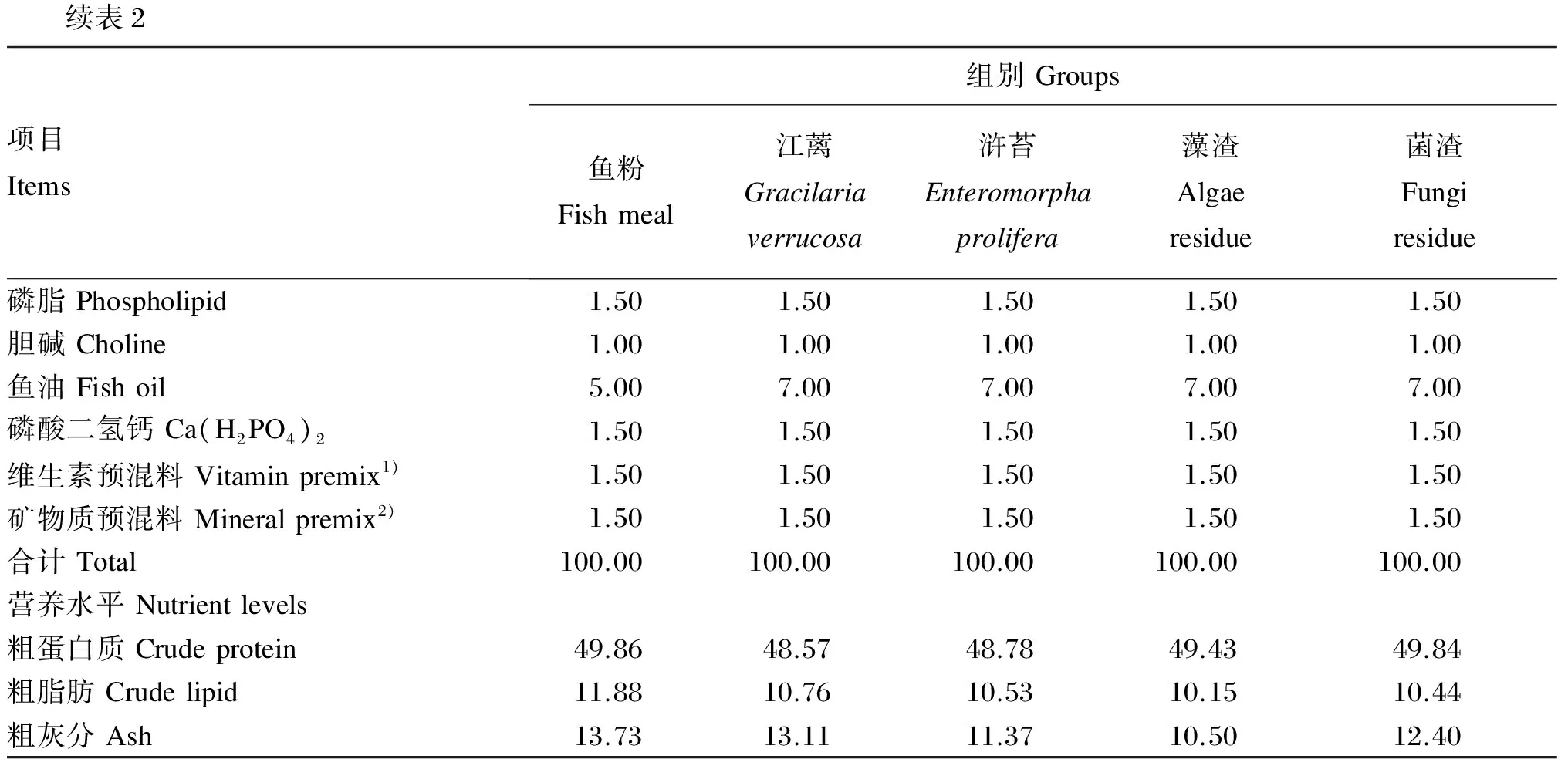
续表2项目Items组别Groups鱼粉Fishmeal江蓠Gracilariaverrucosa浒苔Enteromorphaprolifera藻渣Algaeresidue菌渣Fungiresidue磷脂Phospholipid1.501.501.501.501.50胆碱Choline1.001.001.001.001.00鱼油Fishoil5.007.007.007.007.00磷酸二氢钙Ca(H2PO4)21.501.501.501.501.50维生素预混料Vitaminpremix1)1.501.501.501.501.50矿物质预混料Mineralpremix2)1.501.501.501.501.50合计Total100.00100.00100.00100.00100.00营养水平Nutrientlevels粗蛋白质Crudeprotein49.8648.5748.7849.4349.84粗脂肪Crudelipid11.8810.7610.5310.1510.44粗灰分Ash13.7313.1111.3710.5012.40
1)每千克维生素预混料含有 Contained the following per kg of vitamin premix:VA 375 000 IU,VD375 000 IU,VE 3 000 mg,VK3900 mg,VB1600 mg,VB2600 mg,VB6600 mg,VB123.7 mg,D-泛酸钙D-calcium pantothenate 2 400 mg,烟酸胺 niacinaminde 4 500 mg,叶酸 folic acid 185 mg,D-生物素D-biotin 7.5 mg,肌醇 inositol 3 000 mg,VC 10 500 mg。
2)每千克矿物质预混料含有Contained the following per kg of mineral premix:Zn 1 750 mg,Mn 1 050 mg,Cu 410 mg,Fe 1 150 mg,Co 60 mg,I 50 mg,Se 15 mg。
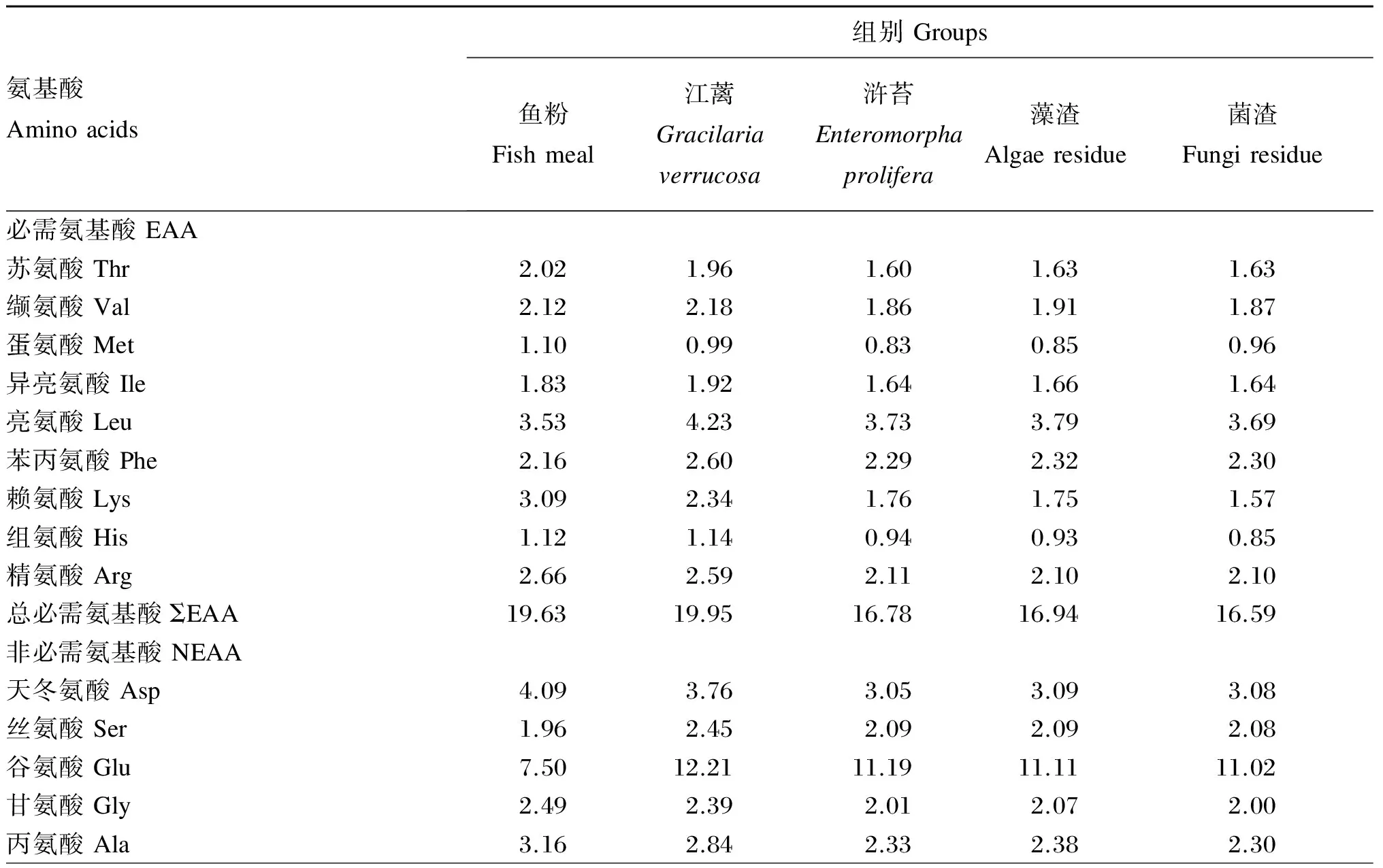
表3 试验饲料氨基酸组成(干物质基础)
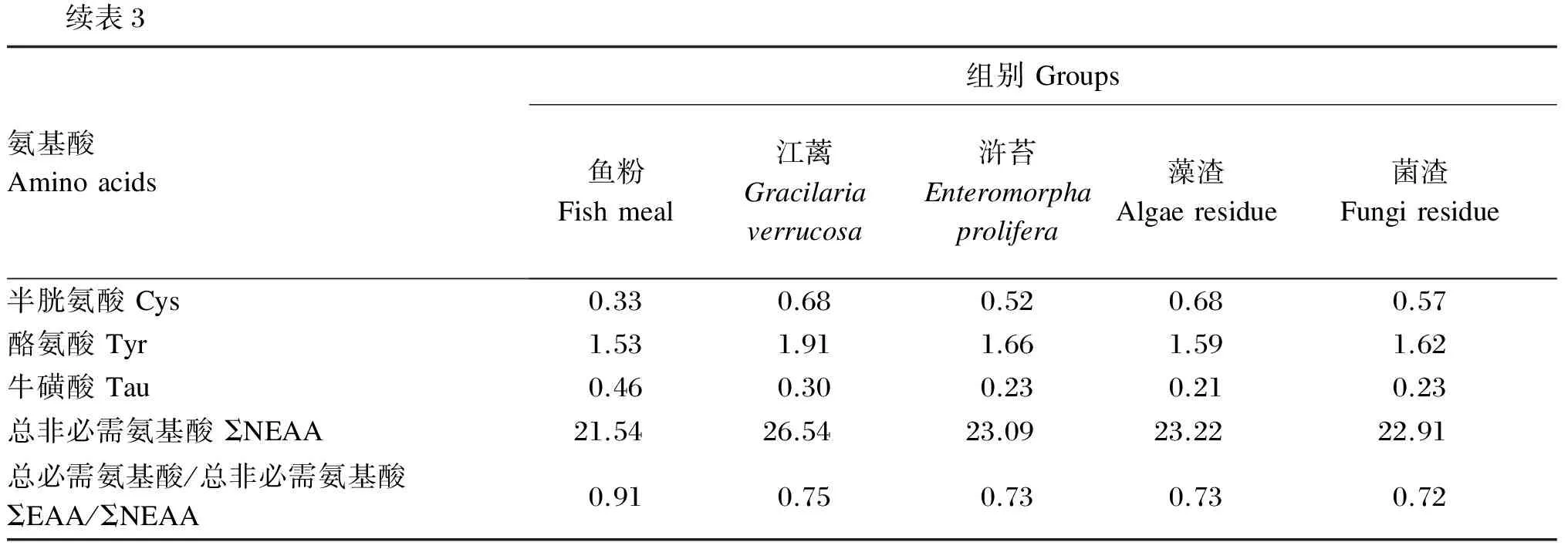
续表3氨基酸Aminoacids组别Groups鱼粉Fishmeal江蓠Gracilariaverrucosa浒苔Enteromorphaprolifera藻渣Algaeresidue菌渣Fungiresidue半胱氨酸Cys0.330.680.520.680.57酪氨酸Tyr1.531.911.661.591.62牛磺酸Tau0.460.300.230.210.23总非必需氨基酸ΣNEAA21.5426.5423.0923.2222.91总必需氨基酸/总非必需氨基酸ΣEAA/ΣNEAA0.910.750.730.730.72
1.2 饲养管理
养殖试验在山东省烟台市天源水产有限公司进行,试验所用大菱鲆幼鱼取自天源水产有限公司。试验开始前先使用对照组饲料暂养1周,使其适应养殖环境和饲料的大小、硬度。试验采用自然光照,养殖模式为流水养殖,水源为深井海水,水温保持在12~14 ℃,溶氧浓度在5.5 mg/L左右,盐度在35‰左右,pH 7.5~8.0。
试验开始前停食24 h,然后随机选取健康、规格一致的大菱鲆幼鱼,初始体重为(16.00±0.11) g,随机分配到15个容积为150 L的塑料养殖桶中,每个桶中投放25尾。将15个养殖桶随机分为5组,每组3个养殖桶,每组随机投喂1种试验饲料。养殖试验期间每天早、晚各表观饱食投喂1次,投喂0.5 h后对每个桶内的残饵计数,根据每100粒饲料的平均质量,计算残饵质量。试验时间为2016年9月至2016年11月,养殖周期共77 d。
1.3 样品采集与分析
试验开始前随机取20尾大菱鲆幼鱼作为初始鱼,用于常规营养成分分析。试验结束后,饥饿24 h,对每桶鱼进行计数、称重。每桶随机取3尾鱼,尾静脉取血,用1%肝素钠抗凝,4 ℃静置4 h后3 500 r/min离心10 min取上层血清,置于液氮中保存;采血后的鱼称重并测量体长,解剖分离内脏团和肝脏并称重,用于计算肥满度、肝体比和脏体比,取肝脏置于液氮中保存;每尾鱼去掉皮,取背部同一部位的肌肉置于液氮中保存,用于肌肉氨基酸组成的测定。每桶另随机取3尾鱼用于体成分分析。每桶再随机取1尾鱼,解剖后取其前肠和中肠,置于Davidson’s固定液(95%乙醇330 mL、甲醛220 mL、冰醋酸115 mL、纯水335 mL),固定24 h后转移至70%乙醇中保存待测。
饲料和鱼体常规营养成分分析参考AOAC(1995)的方法。其中,水分含量测定采用105 ℃烘干至恒重法,粗蛋白质含量测定采用凯氏定氮法(VELP凯氏定氮仪,UDK-142 Automatic Distillation Unit,意大利),粗脂肪含量测定采用索氏抽提法,以石油醚作为抽提液(SOXTEC2050 FOSS脂肪测定仪,瑞典),粗灰分含量采用马弗炉(550 ℃)灼烧6 h测得。
血清中甘油三酯(TG)、总胆固醇(TC)、高密度脂蛋白胆固醇(HDL-C)、低密度脂蛋白胆固醇(LDL-C)、丙二醛(MDA)和总胆汁酸(TBA)含量,超氧化物歧化酶(SOD)、酸性磷酸酶(ACP)、碱性磷酸酶(AKP)、谷草转氨酶(GOT)、谷丙转氨酶(GPT)活性以及肝脏中GOT、GPT活性均采用南京建成生物工程研究所生产的试剂盒进行测定。
饲料和肌肉中氨基酸组成的测定参考GB/T 18246—2000的方法,使用L-8900全自动氨基酸分析仪(Hitachi,日本)测定。检测了饲料和鱼体肌肉中9种必需氨基酸、7种非必需氨基酸和牛磺酸的含量,色氨酸因酸水解破坏而未检测。
肠道组织结构分析:采用石蜡包埋和苏木精-伊红(HE)染色的方法制作石蜡切片,并在显微镜(80i,Nikon,日本)下观察,拍照,利用Photoshop CC 2017软件测量黏膜皱襞高度、上皮细胞高度和微绒毛高度。
1.4 计算公式
成活率(survival rate,SR,%)=100×
终末鱼尾数/初始鱼尾数;摄食率(feed intake,FI,%/d)=100×
摄食饲料干重/[试验天数×(初始体重+
终末体重)/2];增重率(weight gain rate,WGR,%)=100×
(终末体重-初始体重)/初始体重;特定生长率(special growth rate,SGR,%/d)=
100×(ln终末体重-ln初始体重)/试验天数;饲料效率(feed efficiency ratio,FER)=
(终末体重-初始体重)/摄食饲料干重;蛋白质沉积率(protein productive value,PPV,%)=
100×鱼体蛋白质沉积量/总饲料蛋白质摄入量;蛋白质效率(protein efficiency ratio,PER)=
(终末体重-初始体重)/总饲料蛋白质摄入量;肝体比(heaptosomatic index,HSI,%)=
100×肝脏重/体重;脏体比(viscerosomatic index,VSI,%)=
100×内脏团重/体重;肥满度(condition factor,CF,g/cm3)=
100×体重/体长3。
1.5 数据分析
试验数据使用SPSS 17.0软件进行处理,采用单因素方差分析(one-way ANOVA)程序进行方差分析,若存在显著差异(P<0.05)则采用Duncan氏法进行组间的多重比较。结果以平均值±标准误(mean±SE)表示。
2 结 果
2.1 江蓠、浒苔、藻渣和菌渣替代鱼粉对大菱鲆幼鱼生长性能和形体指标的影响
江蓠、浒苔、藻渣和菌渣替代鱼粉对大菱鲆幼鱼生长性能的影响如表4所示。各组大菱鲆幼鱼的存活率均介于92%~96%,组间无显著差异(P>0.05);江蓠、藻渣、菌渣组的增重率和特定生长率与鱼粉组均无显著差异(P>0.05),浒苔组显著低于鱼粉组(P<0.05),菌渣和浒苔组显著低于藻渣组(P<0.05),江蓠组与浒苔、藻渣、菌渣组均没有显著差异(P>0.05);浒苔和藻渣组的摄食率显著高于鱼粉和江蓠组(P<0.05),菌渣组显著高于江蓠组(P<0.05),菌渣、江蓠组与鱼粉组无显著差异(P>0.05);鱼粉与江蓠组的饲料效率和蛋白质效率无显著差异(P>0.05),但显著高于浒苔、藻渣、菌渣组(P<0.05);鱼粉和江蓠组的蛋白质沉积率显著高于浒苔和藻渣组(P<0.05),江蓠组与菌渣组无显著差异(P>0.05)。
鱼粉组脏体比显著低于藻渣组(P<0.05),与江蓠、浒苔和菌渣组无显著差异(P>0.05);各组的肝体比和肥满度无显著差异(P>0.05)。
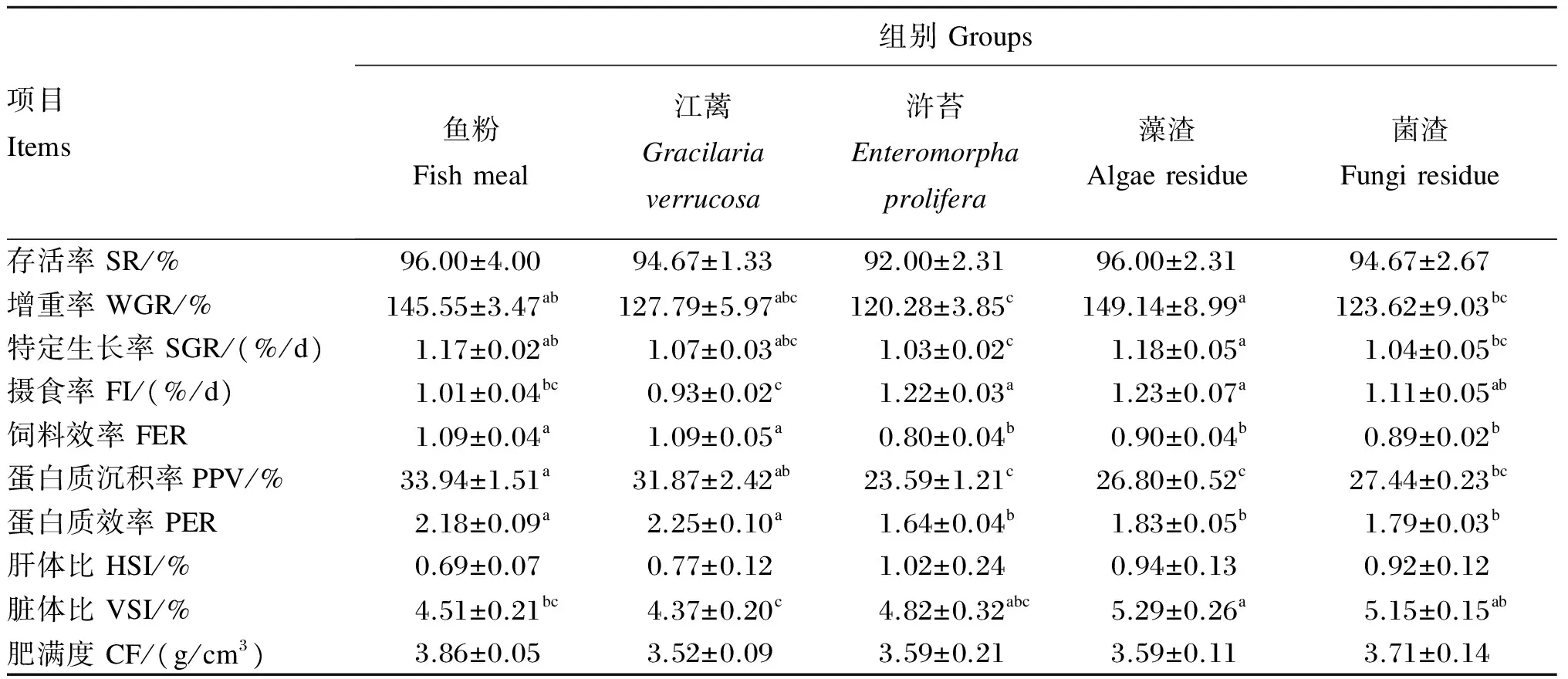
表4 江蓠、浒苔、藻渣和菌渣替代鱼粉对大菱鲆幼鱼生长性能和形体指标的影响
同行数据肩标无字母或相同字母表示差异不显著(P>0.05),不同字母表示差异显著(P<0.05)。下表同。
In the same row, values with no letter or the same letter superscripts mean no significant difference (P>0.05), while with different letter superscripts mean significant difference (P<0.05). The same as below.
2.2 江蓠、浒苔、藻渣和菌渣替代鱼粉对大菱鲆幼鱼血清和肝脏生化指标的影响
江蓠、浒苔、藻渣和菌渣替代鱼粉对大菱鲆幼鱼血清TG、TC、HDL-C、LDL-C和TBA含量的影响如表5所示。各组血清中TG、LDL-C和TBA的含量无显著差异(P>0.05);浒苔组血清中TC含量显著低于藻渣组(P<0.05),显著高于鱼粉和菌渣组(P<0.05),与江蓠组无显著差异(P>0.05);浒苔和藻渣组血清中HDL-C含量显著高于鱼粉和菌渣组(P<0.05),与江蓠组无显著差异(P>0.05)。
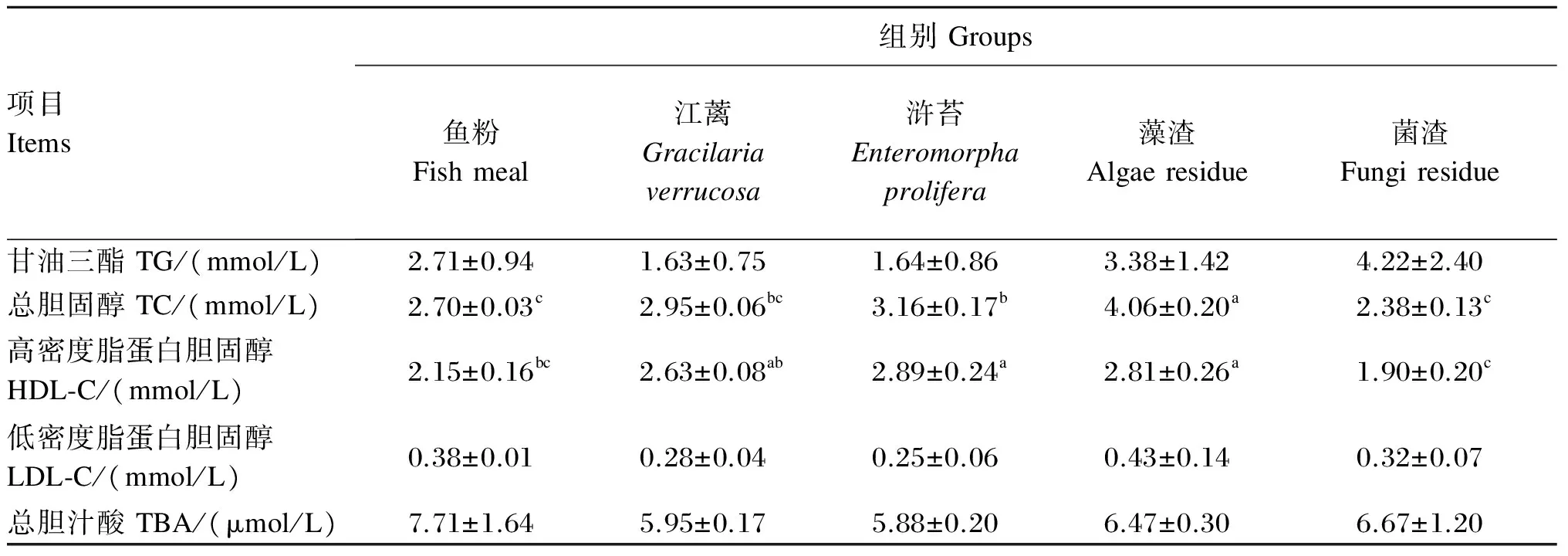
表5 江蓠、浒苔、藻渣和菌渣替代鱼粉对大菱鲆幼鱼血清甘油三酯、总胆固醇、高密度脂蛋白、低密度脂蛋白和总胆汁酸含量的影响
江蓠、浒苔、藻渣和菌渣替代鱼粉对大菱鲆血清和肝脏中GPT和GOT活性的影响如表6所示。各组血清中GPT和GOT活性和肝脏中GOT活性无显著差异(P>0.05);鱼粉组肝脏GPT活性显著高于菌渣和江蓠组(P<0.05),与浒苔和藻渣组无显著差异(P>0.05)。
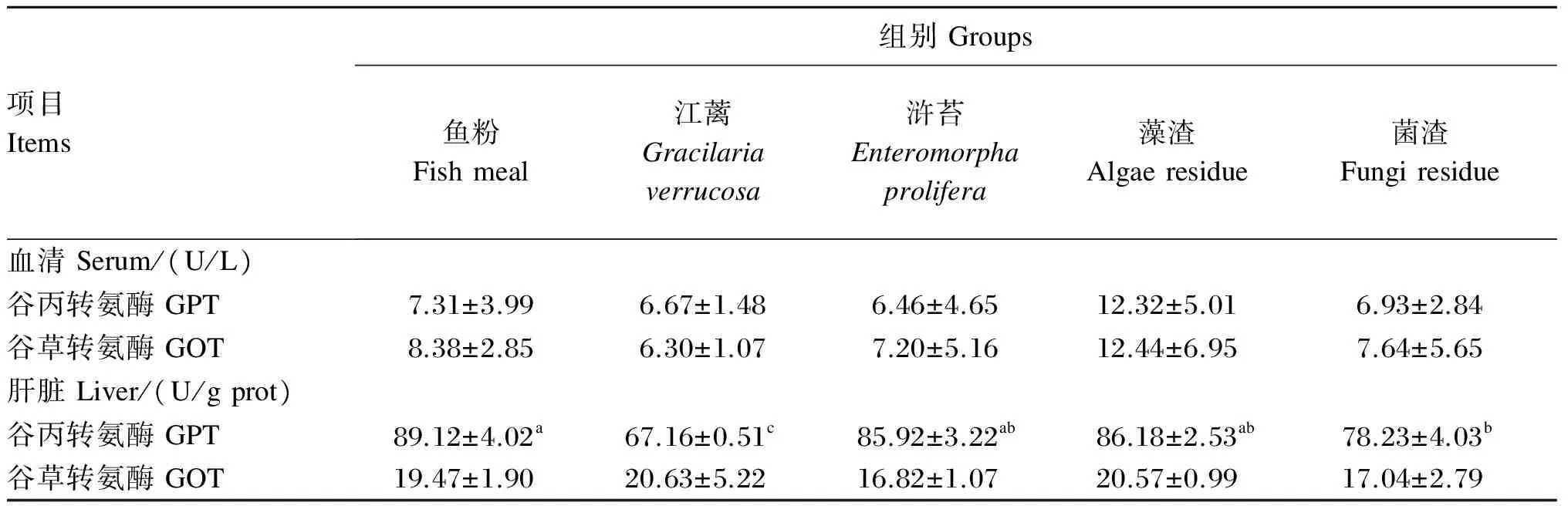
表6 江蓠、浒苔、藻渣和菌渣替代鱼粉对大菱鲆幼鱼血清和肝脏中谷丙转氨酶和谷草转氨酶活性的影响
江蓠、浒苔、藻渣和菌渣替代鱼粉对大菱鲆幼鱼血清非特异性免疫和抗氧化指标的影响如表7所示。江蓠、浒苔、藻渣和菌渣组血清中ACP活性与鱼粉组均无显著差异(P>0.05),藻渣组显著高于浒苔组(P<0.05);各组血清中AKP活性和MDA含量无显著差异(P>0.05);江蓠、浒苔、藻渣和菌渣组血清中SOD活性与鱼粉组均无显著差异(P>0.05),江蓠和藻渣组显著高于浒苔和菌渣组(P<0.05)。
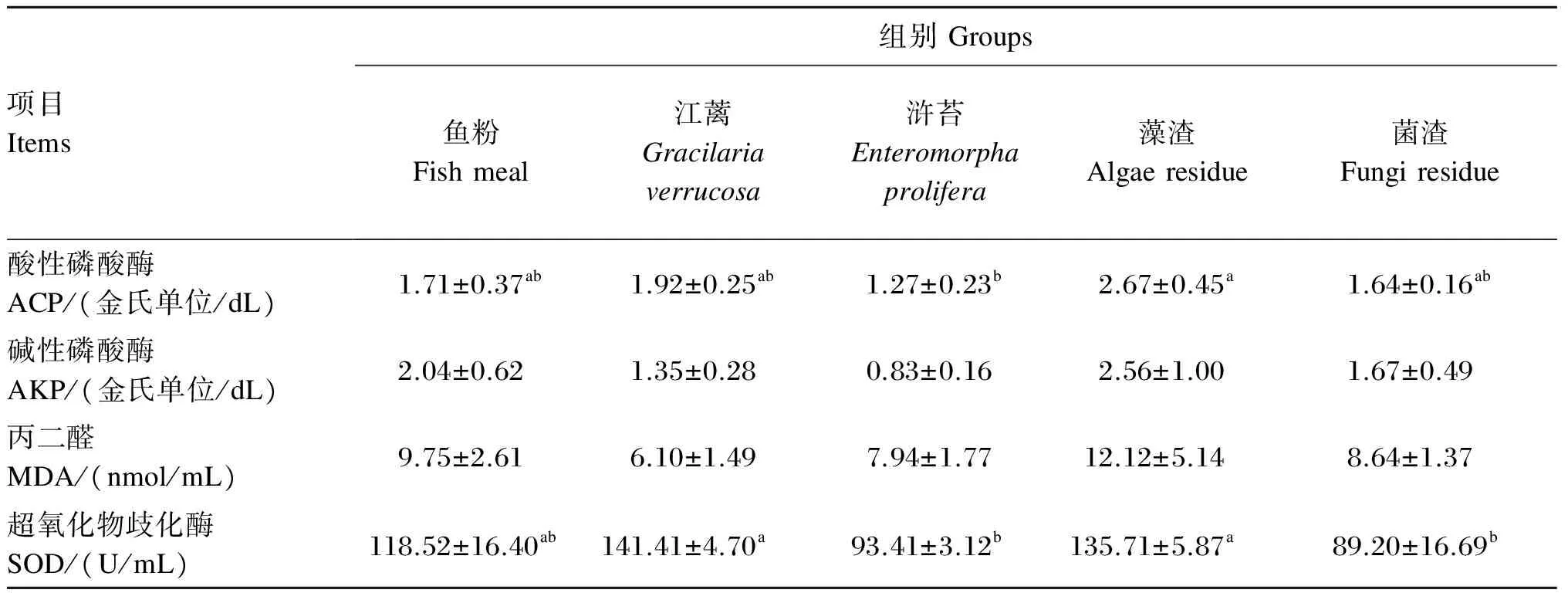
表7 江蓠、浒苔、藻渣和菌渣替代鱼粉对大菱鲆幼鱼血清非特异性免疫和抗氧化指标的影响
2.3 江蓠、浒苔、藻渣和菌渣替代鱼粉对大菱鲆幼鱼体组成和肌肉氨基酸组成的影响
江蓠、浒苔、藻渣和菌渣替代鱼粉对大菱鲆幼鱼体组成的影响如表8所示。江蓠、浒苔、藻渣和菌渣组鱼体的水分和粗灰分含量与鱼粉组相比均无显著差异(P>0.05);鱼粉组鱼体粗蛋白质含量显著高于江蓠、浒苔组(P<0.05),与藻渣和菌渣组无显著差异(P>0.05);藻渣和菌渣组鱼体粗脂肪含量显著高于鱼粉组(P<0.05),与江蓠和浒苔组无显著差异(P>0.05)。
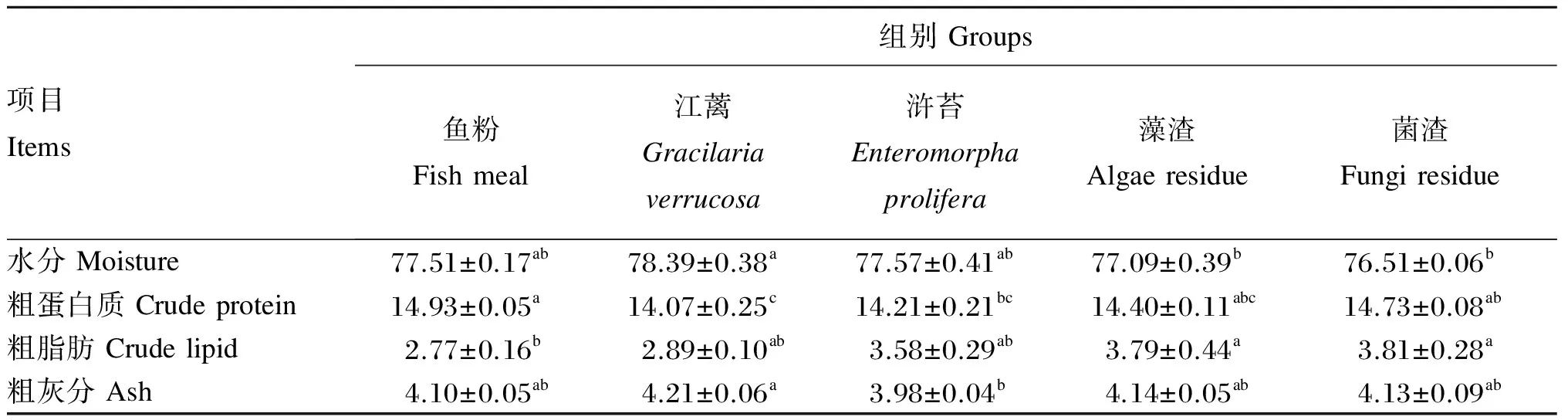
表8 江蓠、浒苔、藻渣和菌渣替代鱼粉对大菱鲆幼鱼体组成的影响
江蓠、浒苔、藻渣和菌渣替代鱼粉对大菱鲆幼鱼肌肉氨基酸组成的影响如表9所示。鱼粉组肌肉赖氨酸含量显著高于浒苔、藻渣组(P<0.05),与江蓠和菌渣组无显著差异(P>0.05);鱼粉组肌肉组氨酸和牛磺酸含量显著高于江蓠、浒苔、藻渣和菌渣组(P<0.05);江蓠组肌肉丙氨酸含量显著高于鱼粉和菌渣组(P<0.05),与浒苔和藻渣组无显著差异(P>0.05);各组肌肉其他氨基酸含量均无显著差异(P>0.05)。鱼粉组肌肉总必需氨基酸含量显著高于浒苔和藻渣组(P<0.05),与江蓠和菌渣组无显著差异(P>0.05);各组肌肉总非必需氨基酸含量无显著差异(P>0.05);鱼粉组总氨基酸含量显著高于藻渣组(P<0.05),与江蓠、浒苔和菌渣组无显著差异(P>0.05)。
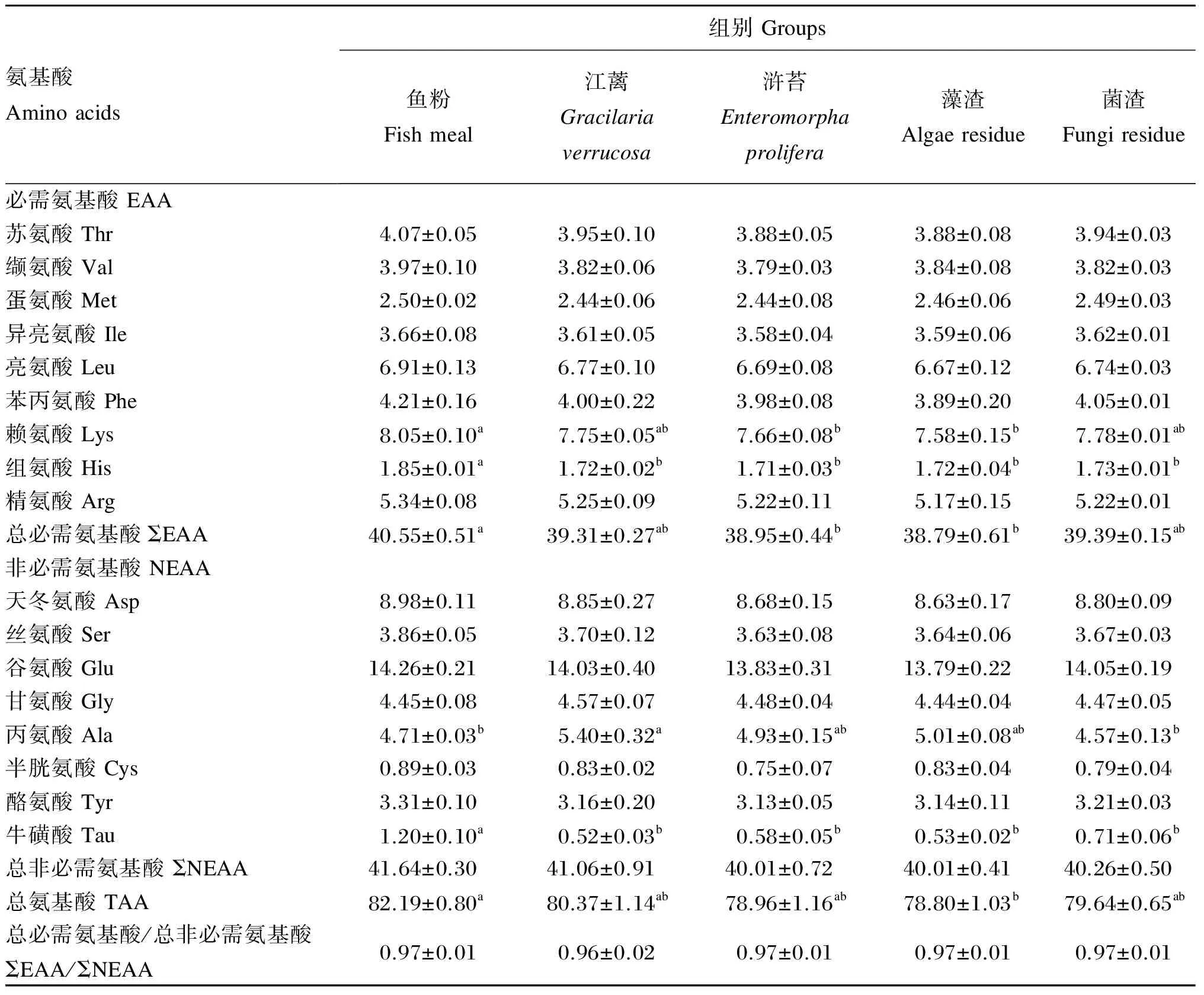
表9 江蓠、浒苔、藻渣和菌渣替代鱼粉对大菱鲆幼鱼肌肉氨基酸组成的影响(干物质基础)
2.4 江蓠、浒苔、藻渣和菌渣替代鱼粉对大菱鲆幼鱼肠道组织结构的影响
各组大菱鲆幼鱼的中肠横切面结构如图1所示。
江蓠、浒苔、藻渣和菌渣替代鱼粉对大菱鲆幼鱼肠道组织结构的影响如表10所示。藻渣组前肠皱襞高度显著高于鱼粉组(P<0.05),显著低于菌渣组(P<0.05),与江蓠、浒苔组无显著差异(P>0.05);藻渣组前肠上皮细胞高度显著高于鱼粉、江蓠和浒苔组(P<0.05),与菌渣组无显著差异(P>0.05);各组前肠微绒毛高度无显著差异(P>0.05)。菌渣组中肠皱襞高度显著高于鱼粉组(P<0.05),与江蓠、浒苔和藻渣组无显著差异(P>0.05);各组中肠上皮细胞高度和微绒毛高度均无显著差异(P>0.05)。
3 讨 论
3.1 江蓠、浒苔、藻渣和菌渣替代鱼粉对大菱鲆幼鱼生长性能的影响
关于海藻在水产饲料中的应用已有许多报道,主要包括红藻门的江蓠、石花菜、紫菜,绿藻门的石莼、浒苔,褐藻门的海带等多种常见海藻[9-14]。相当一部分研究表明,饲料中添加少量的海藻(2.5%~10.0%)有利于鱼类的生长,可以提高生长性能,增加摄食率和饲料效率,提高生理活性、抗病力和应激能力[9-12]。
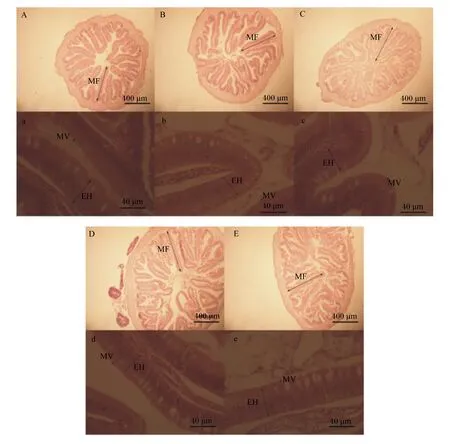
MF:黏膜皱襞;EH:肠上皮细胞;MV:微绒毛。A、a:鱼粉组;B、b:江蓠组;C、c:浒苔组;D、d:藻渣组;E、e:菌渣组。A、B、C、D和E的比例尺为400 μm,a、b、c、d和e的比例尺为40 μm。
MF: mucosal fold; EH: enterocyte; MV: microvillu. A and a: fish meal group; B and b:Gracilariaverrucosagroup; C and c:Enteromorphaproliferagroup; D and d: algae residue group ; E and e: fungi residue. The scale bar of A, B, C, D and E was 400 μm and the scale bar of a, b, c, d and e was 40 μm.
图1各组大菱鲆幼鱼中肠横切面结构
Fig.1 The midgut transection structure of juvenile turbot in each group
本试验中,将江蓠、浒苔和藻渣与植物蛋白质配合替代饲料中部分鱼粉后,江蓠组饲料中各主要必需氨基酸含量与鱼粉组相比没有显著降低(表3),而浒苔组饲料中赖氨酸、蛋氨酸和组氨酸含量大大低于鱼粉组,这可能是江蓠组大菱鲆幼鱼生长速度、饲料效率、蛋白质沉积率优于浒苔组的主要原因。本研究发现藻渣组大菱鲆幼鱼的生长效果优于浒苔组而与鱼粉组无显著差异,虽然藻渣组饲料的必需氨基酸含量与鱼粉组饲料相比有不同程度地降低,其可能的原因是藻渣含有褐藻糖胶、岩藻多糖、褐藻淀粉以及褐藻胶等多糖[15],这些多糖促进了大菱鲆幼鱼的生长,这与林建斌等[16]报道的饲料中添加0.6%的海带多糖可以提高珍珠龙胆石斑鱼的生长,降低饲料系数的结果一致。对于其他鱼种也有类似发现,杨晴等[17]报道饲料中有效浓度为0.10%的褐藻糖胶可以提高黄颡鱼的增重率和特定生长率,并能提高胃和肠道脂肪酶的活性。
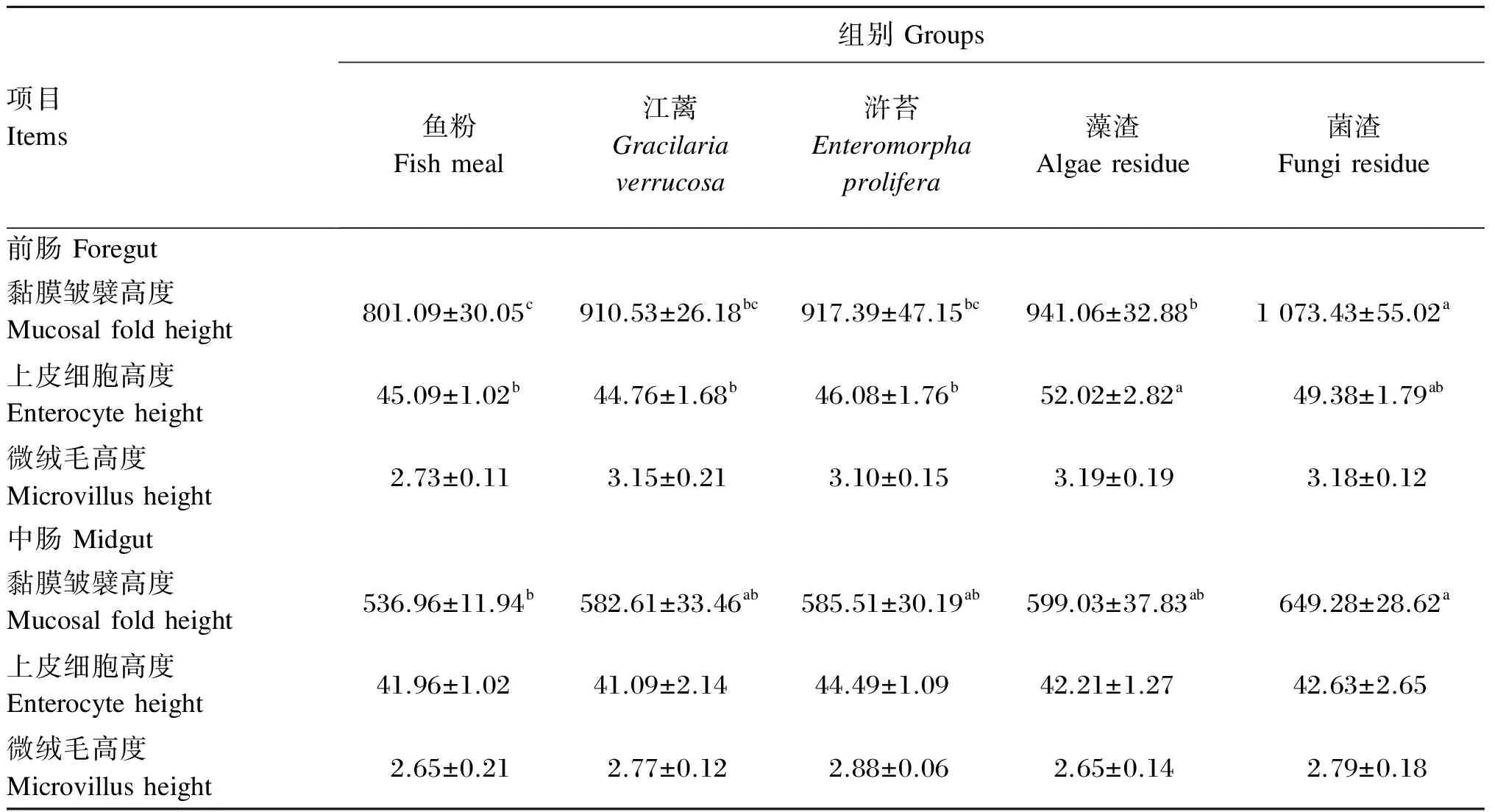
表10 江蓠、浒苔、藻渣和菌渣替代鱼粉对大菱鲆幼鱼肠道组织结构的影响
本研究发现,浒苔和藻渣组的摄食率显著高于鱼粉和江蓠组。浒苔和藻渣提高大菱鲆幼鱼摄食率可能与其中具有诱食作用的物质有关,有研究表明海藻中普遍存在一些具有诱食作用的物质,如二甲基-β-丙酸噻亭(DMPT或DMSP)[18-19],可以增加摄食率。但本试验中江蓠组大菱鲆幼鱼的摄食率并没有提高,反而出现下降,这可能是因为江蓠中含有的诱食成分较少,因本研究未对相关诱食成分进行测定,具体原因还有待进一步研究。虽然江蓠组大菱鲆幼鱼的摄食率低,但其生长效果较好,这除了与饲料中必需氨基酸含量高有关外,饲料中的牛磺酸对鱼体生长可能也起到促进作用。有研究表明,牛磺酸可以调节消化酶的活性,增强鱼体对饲料中营养物质的消化吸收,提高饲料效率[20]。江蓠组饲料中牛磺酸的含量高于浒苔和藻渣组,这可能也是江蓠组大菱鲆幼鱼虽然摄食率低但并没有影响其生长性能的原因之一
在以往的利用植物蛋白质替代鱼粉对大菱鲆影响的研究中,饲料中植物蛋白质替代鱼粉的比例较低。有研究表明,大豆浓缩蛋白只能替代饲料中17%的鱼粉而不影响大菱鲆生长[21];玉米蛋白粉可替代大菱鲆饲料中21%的鱼粉而不影响其生长性能和饲料效率[22];复合植物蛋白质替代大菱鲆饲料中20.7%的鱼粉会显著降低其生长性能[23];以酶解动物软骨蛋白粉与植物蛋白质复合后可替代大菱鲆幼鱼饲料中24%鱼粉而不影响其生长与摄食[24]。对于其他鱼类的研究也有类似的结果。王国霞等[25]发现豆粕可以替代饲料中16%的鱼粉而不影响花鲈生长;吴莉芳等[26]发现豆粕替代饲料中13.4%的鱼粉时没有影响黄金鲈的生长;崔存河等[27]发现菜籽粕可以替代饲料中13.5%的鱼粉而不影响虹鳟的生长性能和饲料利用。在本试验中,江蓠、浒苔、藻渣、菌渣与植物蛋白质配合后替代了饲料中35%的鱼粉,而江蓠、藻渣和菌渣组大菱鲆幼鱼并未因植物蛋白质大量替代鱼粉而显著降低生长性能。这说明,与传统陆生植物蛋白相比,江蓠、藻渣作为一种海洋性植物蛋白质来源,对因植物蛋白质大量替代鱼粉而造成的大菱鲆生长性能的下降有一定改善作用;菌渣是富含真菌蛋白的一类蛋白质来源,其中含有的一些物质对大菱鲆的生长也起到了一定的促进作用。
3.2 江蓠、浒苔、藻渣和菌渣替代鱼粉对大菱鲆幼鱼脂肪代谢和非特异性免疫能力的影响
血清中HDL-C和LDL-C含量反映了血清中高密度脂蛋白和低密度脂蛋白含量的多少。高密度脂蛋白含量增加可以使血液中更多的胆固醇被转运到肝脏,低密度脂蛋白含量增加可以使更多的胆固醇和甘油三酯转运到血液[28],二者的综合作用影响了血清中TC和TG的含量。在本试验中,血清TG含量的变化趋势和LDL-C一致,与鱼粉组相比,江蓠和浒苔组血清中LDL-C和TG含量都有所降低,同时浒苔和藻渣组血清中HDL-C含量均显著升高,说明饲料中添加江蓠、浒苔和藻渣对增加血清中的高密度脂蛋白含量、降低血清中的低密度脂蛋白含量有一定作用,有利于大菱鲆的血脂健康。
MDA是自由基氧化的终产物,具有细胞毒性,在生物体内可引起蛋白质、核酸等生命大分子的交联聚合,在生物体外影响线粒体呼吸链复合物及线粒体内关键酶活性[12]。血清中MDA含量可反映机体内脂质的过氧化程度,间接反映组织细胞的氧化损伤程度[12,29]。SOD对机体氧化与抗氧化平衡起着重要作用,能清除超氧阴离子自由基,保护细胞免受损伤。本试验中,江蓠、浒苔、藻渣和菌渣组血清MDA含量和SOD活性与鱼粉组均无显著差异,说明江蓠、浒苔、藻渣和菌渣替代鱼粉对大菱鲆机体的抗氧化能力没有造成不良影响。
血清中GPT、GOT活性的变化是判断肝脏损伤程度的主要依据之一。GPT和GOT主要存在于肝脏中,正常情况下血清中这2种酶的活性很低,当肝脏组织受损时,细胞膜通透性发生改变,GPT和GOT会进入血液,导致血清中GPT和GOT活性上升,因此血清中GPT和GOT的活性可反映肝脏的损伤情况[30]。在本试验中,各组大菱鲆幼鱼血清中GPT与GOT活性均无显著差异,说明江蓠、浒苔、藻渣和菌渣替代鱼粉对大菱鲆幼鱼的肝脏没有造成不良影响。饲料氨基酸主要通过转氨基作用和脱氨基作用在体内代谢转化,鱼类则主要通过联合脱氨基作用满足机体需要,GPT和GOT是鱼类氨基酸代谢中的2个关键酶,它们在肝脏中活性的高低反映了氨基酸代谢程度的强弱和肝脏功能的正常与否[30]。在本试验中,江蓠和菌渣组的肝脏GPT活性显著低于鱼粉组,说明江蓠和藻渣组大菱鲆氨基酸代谢较慢,这可能与鱼体对氨基酸的吸收有关,由于饲料中氨基酸组成略有差异,不同氨基酸吸收速度不同,这可能影响了鱼体对氨基酸的代谢,从而影响了肝脏GPT活性。
水生动物的特异性免疫机制不具备抗体生成的二次反应,非特异性免疫在水生动物的免疫防御中起着至关重要的作用[30]。鱼类血清中ACP、AKP等是评价鱼类非特异性免疫的重要指标,在提高鱼体的抗病力以及抗应激力中发挥促进作用[30-32]。ACP是溶酶体的标志酶,主要参与磷酸酯的代谢调节、信号传导以及能量转化,而AKP可以直接参与生物体磷酸基团的转移和代谢[30]。在本研究中,江蓠、浒苔、藻渣、菌渣组大菱鲆幼鱼血清中ACP、AKP活性与鱼粉组无显著差异,说明以江蓠、浒苔、藻渣和菌渣替代鱼粉对大菱鲆幼鱼的非特异性免疫力无不良影响,并且藻渣组血清中ACP和AKP的活性还高于鱼粉组,这可能是藻渣中的多糖和寡糖起到了一定的增强非特异性免疫力的作用。有研究表明,褐藻多糖硫酸酯可以提高小鼠的免疫力和抗病毒能力[33-34];褐藻酸寡糖可以提高大菱鲆的非特异性免疫力[32]。
3.3 江蓠、浒苔、藻渣和菌渣替代鱼粉对大菱鲆幼鱼体组成和肌肉氨基酸组成的影响
本研究中,各组鱼体的体组成存在一定的差异,这可能与饲料中牛磺酸含量有关。周铭文等[35]发现,牛磺酸可以增加斜带石斑鱼全鱼蛋白质沉积,降低全鱼脂肪沉积;张圆琴等[36]发现,在高植物蛋白质饲料中添加牛磺酸可降低大菱鲆鱼体的脂肪沉积;齐国山[37]发现,牛磺酸增加了初始体重为6.3和48.0 g大菱鲆的蛋白质沉积,降低了脂肪沉积,但对初始体重为165.9 g的大菱鲆同时增加了蛋白质和脂肪沉积。牛磺酸对鱼体蛋白质沉积的调节是通过调节与蛋白质合成代谢相关的激素的分泌来进行的[35],有研究发现,在鲤鱼饲料中添加牛磺酸能够促进甲状腺激素分泌[38],而甲状腺激素能够促进组织分化、生长和发育,加速蛋白质与各种酶的生成。牛磺酸对脂肪沉积的调节则是通过络合胆汁酸,从而刺激胆汁酸分泌来实现的,胆汁酸的分泌可以增加脂肪酸的氧化和能量消耗[39]。在本试验中,鱼粉组饲料和鱼体肌肉中的牛磺酸含量较高,血清中总胆汁酸含量也较高,这与相关研究结果一致,可能是鱼粉组鱼体粗蛋白质含量较高、粗脂肪含量较低的原因之一。
本研究对鱼体肌肉氨基酸组成进行分析后发现,江蓠、浒苔、藻渣和菌渣替代鱼粉对大菱鲆营养的肌肉氨基酸组成存在影响,这主要是受饲料氨基酸组成的影响,有研究表明机体组织的必需氨基酸组成与饲料氨基酸组成存在明显的相关关系[40]。
3.4 江蓠、浒苔、藻渣和菌渣替代鱼粉对大菱鲆幼鱼肠道组织结构的影响
黏膜皱襞高度、上皮细胞高度、微绒毛高度是评价肠道组织形态变化的重要指标,肠道的皱襞高度和微绒毛高度扩大了肠道面积,决定了肠道吸收营养物质的能力。在本研究中,江蓠、浒苔、藻渣和菌渣替代鱼粉对大菱鲆幼鱼的肠道发育均起到一定的促进作用,尤其是藻渣和菌渣,这2组大菱鲆幼鱼的肠道发育非常好,这可能与藻渣和菌渣中的一些糖类物质有关。藻渣中含有海带多糖,以及一些寡糖如甘露糖,菌渣是土曲霉菌发酵生产衣康酸的下脚料,其中含有一些构成真菌细胞的聚糖如几丁质,而壳聚糖是部分或完全脱乙酸基的几丁质的衍生物,被壳聚糖酶水解可得到壳寡糖。有研究发现,饲料中添加海带粗多糖可以提高斜带石斑鱼肠道绒毛长度[41];潘金露[42]发现,饲料中添加壳寡糖可以提高大菱鲆肠道皱襞高度和微绒毛高度;田娟等[43]发现,壳寡糖可以促进吉富罗非鱼肠道发育;于朝磊等[44]报道,在饲料中添加甘露寡糖可以提高半滑舌蹋的肠道皱襞高度,促进肠道微绒毛发育。本研究中藻渣和菌渣中含有的多糖和寡糖可能是促进大菱鲆幼鱼肠道发育的原因之一。
4 结 论
① 将10%的江蓠、藻渣和菌渣与植物蛋白质配合替代饲料中35%的鱼粉对大菱鲆幼鱼的生长性能无不良影响,以10%的浒苔与植物蛋白质配合替代饲料中35%的鱼粉则降低了大菱鲆幼鱼的生长性能。
② 藻渣和菌渣可改善大菱鲆幼鱼的肠道组织结构。
[1] 孔庆辉.2014年鱼粉市场回顾及后期供需展望[J].海洋与渔业,2015(3):89-93
[2] SHIMENO S,MIMA T,IMANAGA T,et al.Inclusion of combination of defatted soybean meal,meat meal,and corn gluten meal to yellowtail diets[J].Nippon Suisan Gakkaishi,1993,59(11):1889-1895.
[3] SANZ A,MORALES A E,DE LA HIGUERA M,et al.Sunflower meal compared with soybean meals as partial substitutes for fish meal in rainbow trout (Oncorhynchusmykiss) diets:protein and energy utilization[J].Aquaculture,1994,128(3/4):287-300.
[4] EUSEBIO P S,COLOSO R M.Nutritional evaluation of various plant protein sources in diets for Asian sea bassLatescalcarifer[J].Journal of Applied Ichthyology,2010,16(2):56-60.
[5] MILLAMENA O M.Replacement of fish meal by animal by-product meals in a practical diet for grow-out culture of grouperEpinepheluscoioides[J].Aquaculture,2002,204(1/2):75-84.
[6] 赵晓丹.食物抗营养因子[M].北京:中国农业大学出版社,2015.
[7] 常巧玲,孙建义.海藻饲料资源及其在水产养殖中的应用研究[J].饲料工业,2006,27(2):62-64.
[8] 李人光,姜永新,姜瑞勇,等.大型海藻作为饲料的综合利用技术[J].科学养鱼,2009(1):64-65.
[9] NORAMBUENA F,HERMON K,SKRZYPCZYK V,et al.Algae in fish feed:performances and fatty acid metabolism in juvenile Atlantic salmon[J].PLoS One,2015,10(4):e0124042.
[10] WASSEF E A,EL-SAYED A F M,KANDEEL K M,et al.Evaluation of PterocladiaRhodophytaand UlvaChlorophytameals as additives to gilthead seabreamSparusauratadiets[J].Egyptian Journal of Aquatic Research,2005,31:321-332.
[11] 李雅婷,陈明,曾帅霖,等.饲料中添加龙须菜对眼斑拟石首鱼生长、脂肪酸组成、免疫及肠道的影响[J].南方水产科学,2016,12(1):85-93.
[12] 周胜强,游翠红,王树启,等.饲料中添加浒苔对黄斑蓝子鱼生长性能与生理生化指标的影响[J].中国水产科学,2013,20(6):1257-1265.
[13] 苏延明,蔡学新,孙俭,等.用几种饲料原料饲喂中间球海胆稚胆的效果[J].大连海洋大学学报,2008,23(3):242-246.
[14] 杨维维,刘文斌,沈美芳,等.海带粉对克氏原螯虾生长、非特异性免疫和肝胰脏抗氧化能力的影响[J].大连海洋大学学报,2014,29(1):40-44.
[15] 许凤清,吴皓.海带多糖的研究进展[J].中国中医药信息杂志,2005,12(6):106-108.
[16] 林建斌,梁萍,朱庆国,等.海带多糖对珍珠龙胆石斑鱼生长性能和免疫力的影响[J].福建农业学报,2017,32(1):17-21.
[17] 杨晴,杨锐,周歧存,等.褐藻糖胶对黄颡鱼幼鱼生长性能和消化酶活性的影响[J].动物营养学报,2014,26(7):1880-1887.
[18] 邹仕庚,陶正国,许毅.DMPT在水产动物营养中的应用[J].广东饲料,2005,14(3):34-35.
[19] 王亮.部分大型海藻DMSP含量的检测[D].硕士学位论文.苏州:苏州大学,2012.
[20] 高春生,范光丽,王艳玲.牛磺酸对黄河鲤鱼生长性能和消化酶活性的影响[J].中国农学通报,2007,23(6):645-647.
[21] DAY O J,GONZLEZ H G P.Soybean protein concentrate as a protein source for turbotScophthalmusmaximusL.[J].Aquaculture Nutrition,2015,6(4):221-228.
[22] REGOST C,ARZEL J,KAUSHIK S J.Partial or total replacement of fish meal by corn gluten meal in diet for turbot (Psettamaxima)[J].Aquaculture,1999,180(1/2):99-117.
[23] 陈超,陈京华.牛磺酸、晶体氨基酸对大菱鲆摄食、生长和饲料利用率的影响[J].中国农学通报,2012,28(23):108-112.
[24] 刘运正,何艮,麦康森,等.新型复合动植物蛋白源部分替代鱼粉对大菱鲆幼鱼生长和肉质的影响[J].中国海洋大学学报(自然科学版),2016,46(1):33-39.
[25] 王国霞,付晶晶,黄燕华,等.5种植物蛋白源替代鱼粉对花鲈生长性能和消化酶活性的影响[J].湖北农业科学,2014,53(4):866-870.
[26] 吴莉芳,王俊岭,邢秀苹,等.不同大豆蛋白源对黄金鲈生长、饲料利用及肌肉营养成分的影响[J].西北农林科技大学学报(自然科学版),2017,45(1):1-6.
[27] 崔存河,杨成辉,卜宪勇,等.菜粕替代鱼粉对虹鳟(Oncorhynchusmykiss)生长、体组成和血液学指标的影响[J].饲料工业,2016,37(22):21-25.
[28] 纪利芹,蒋克勇,韩龙江,等.连续降温对大菱鲆成鱼代谢机能的影响[J].海洋科学,2014,38(5):46-53.
[29] JAIN S K,LIM G.Pyridoxine and pyridoxamine inhibits superoxide radicals and prevents lipid peroxidation,protein glycosylation,and (Na++K+)-ATPase activity reduction in high glucose-treated human erythrocytes[J].Free Radical Biology & Medicine,2001,30(3):232-237.
[30] 严俊丽,陈四清,常青,等.南极磷虾粉替代鱼粉对圆斑星鲽幼鱼生长性能、血清和肝脏生化指标及血清非特异性免疫指标的影响[J].动物营养学报,2016,28(11):3503-3510.
[31] 宋晓玲,杨绪彤,思瀚文,等.双歧杆菌细胞壁肽聚糖的分离及其对二种海产动物免疫活性的影响[J].水产学报,2005,29(3):350-355.
[32] 霍圃宇,潘金露,韩雨哲,等.褐藻酸寡糖对大菱鲆幼鱼生长性能、血液学指标及非特异性免疫影响[J].广东海洋大学学报,2015,35(4):10-16.
[33] 刘宪丽,刘东颖,汪艳秋,等.褐藻多糖硫酸酯免疫调节和抗肿瘤活性研究[J].中国微生态学杂志,2010,22(12):1074-1076.
[34] 刘晓东,王为栋,张福波,等.褐藻多糖硫酸脂对小鼠免疫及抗日本乙型脑炎病毒的影响[J].黑龙江畜牧兽医,2016(19):32-35.
[35] 周铭文,王和伟,叶继丹.斜带石斑鱼生长性能、体成分和组织游离氨基酸含量对饲料中牛磺酸含量的响应[J].动物营养学报,2015,27(3):785-794.
[36] 张圆琴,张越,卫育良,等.大菱鲆鱼体脂肪累积调节方法研究[J].上海海洋大学学报,2016,25(5):700-709.
[37] 齐国山.饲料中牛磺酸、蛋氨酸、胱氨酸、丝氨酸和半胱胺对大菱鲆生长性能及牛磺酸合成代谢的影响[D].博士学位论文.青岛:中国海洋大学,2012.
[38] 邱小琮,赵红雪.牛磺酸对鲤生长及血清T3、T4含量的影响[J].淡水渔业,2006,36(1):22-24.
[39] KIM S M,MATSUNARI H,NOMURA K,et al.Effect of dietary taurine and lipid contents on conjugated bile acid composition and growth performance serum biochemical indices related to lipoprotein metabolism of juvenile Japanese flounderParalichthysolivaceus[J].Fisheries Science,2008,74(4):875-881.
[40] 崔燕燕,张南南,马倩倩,等.四种植物蛋白对中华绒螯蟹幼蟹生长性能、氨基酸沉积率和抗氧化酶活性的影响[J].水生生物学报,2017,41(1):146-154.
[41] 李文武.海带粗多糖对斜带石斑鱼血清指标和肠道功能的影响[D].硕士学位论文.福州:福建农林大学,2015.
[42] 潘金露.饲料中壳寡糖和褐藻酸寡糖对大菱鲆(Scophthalmusmaximus)消化及肠道菌群的影响[D].硕士学位论文.大连:大连海洋大学,2016.
[43] 田娟,孙立威,文华,等.壳寡糖对吉富罗非鱼幼鱼生长性能、前肠组织结构及肠道主要菌群的影响[J].中国水产科学,2013,20(3):561-568.
[44] 于朝磊,常青,吕云云.甘露寡糖对半滑舌鳎(CynoglossussemilaevisGünther)稚鱼生长、肠道发育和非特异性免疫水平的影响[J].渔业科学进展,2014,35(6):53-59.
*Corresponding author, professor, E-mail: liangmq@ysfri.ac.cn
猜你喜欢
黑龙江水产(2022年6期)2022-12-03
当代水产(2022年8期)2022-09-20
当代水产(2022年6期)2022-06-29
中华环境(2021年10期)2021-11-22
当代水产(2019年6期)2019-07-25
当代水产(2019年1期)2019-05-16
中国水产(2019年3期)2019-03-25
现代食品(2018年10期)2018-02-14
临床医药文献杂志(电子版)(2017年98期)2017-03-06
化工管理(2017年29期)2017-03-03
