
大气增温对滇西北高原典型湿地湖滨带优势植物凋落物质量衰减的影响
2018-01-04 05:33张晓宁刘振亚李丽萍肖德荣
生态学报 2017年23期
张晓宁,刘振亚,李丽萍,王 行,张 贇,孙 梅,肖德荣
西南林业大学国家高原湿地研究中心, 昆明 650224
大气增温对滇西北高原典型湿地湖滨带优势植物凋落物质量衰减的影响
张晓宁,刘振亚,李丽萍,王 行,张 贇,孙 梅,肖德荣*
西南林业大学国家高原湿地研究中心, 昆明 650224
气候变暖对湿地生态系统碳循环的影响受到国内外的广泛关注。研究大气增温对植物凋落物分解的影响是掌握气候变暖对湿地生态系统碳循环影响过程及其作用机制的基础。通过开顶式生长室(Open-top Chambers,OTCs)模拟大气增温(2.0±0.5)℃和(3.5±0.5)℃,以滇西北高原典型湿地纳帕海湖滨带的优势植物茭草(Zizaniacaduciflora)、水葱(Scirpustabernaemontani)、黑三棱(Sparganiumstoloniferum)和杉叶藻(Hippurisvulgaris)为研究对象,研究其凋落物在大气界面,水界面,土-水界面的质量衰减对大气增温的响应。结果表明,(1)大气增温促进了4种植物凋落物在不同分解界面的质量衰减,但其质量衰减率在不同分解界面对增温的响应存在差异。其中,增温对水界面植物凋落物质量衰减的促进作用最大,并随着增温的幅度的增加而增加。其次是土-水界面,增温对大气界面植物凋落物质量衰减的促进作用最小。(2)植物凋落物初始C/N值与其凋落物质量衰减率呈负相关,大气增温对低初始C/N值物种凋落物质量衰减的促进更为显著。(3)相对于大气增温,植物凋落物初始C/N值对其凋落物质量衰减的影响高于大气增温对其质量衰减的影响。研究表明,气候变暖将促进湿地植物凋落物的分解,进而对湿地生态系统物质循环及其生态功能产生影响,其影响程度与湿地植物物种有关,其作用机制有待于进一步深入研究。
大气增温;湖滨带植物;凋落物分解;滇西北高原
植物凋落物分解是生态系统碳循环的关键环节,其过程影响着生态系统碳的输入和输出[1-2],进而对生态系统碳源/汇功能产生影响。当前,以温度升高为主要特征的气候变暖对自然生态系统碳循环的影响受到广泛关注[3]。因此,研究增温对植物凋落物分解的影响成为掌握气候变暖对自然生态系统碳循环影响的基础[4]。
温度是影响植物凋落物分解最为重要的因子之一[5]。有研究表明,温度增加可使凋落物分解速率增加4%—7%[6],导致其土壤净碳通量增加200%[7]。但是,不同植物物种凋落物分解对增温响应存在差异,在相同增温条件下,阔叶林和针叶林分解量分别增加了1.87%和2.63%[8]。也有研究表明,由于土壤湿度变化产生的补偿机制对小范围温度升高导致的凋落物分解速率增加产生了抑制,增温对植物凋落物分解速率没有影响[9- 10]。另外,有研究发现增温处理明显降低植物凋落物的分解率[11]。可见,温度与植物凋落物分解速率之间的关系存在较大的不确定性,尚需深入研究。
滇西北高原湿地地处青藏高原东南缘横断山区,属我国封闭与半封闭的独特湿地类型[12]。由于该区域常年冷湿的环境条件,大量植物凋落物不易分解而成为湿地生态系统碳汇的基础,对区域碳循环和大气温室气体的平衡起着重要的调节作用[13-14]。前期研究表明,经过1 a的分解,该区域典型湿地纳帕海湖滨带优势植物凋落物残留量较高[13],其湿地土壤碳储量已达32.92×104t[14]。滇西北高原属气候变化敏感区,与全球气候变暖趋势一致,该区域在过去50年年均温度自1960年的5.75℃升高至2011年的7.07℃,温度升高率达22.96%,显著高于云南省的其他地区[15]。气候变暖对该区域湿地生态系统碳循环有何影响,其影响程度如何已受到广泛关注[3],但目前尚缺乏有关研究。
本研究选取滇西北高原典型湿地纳帕海湖滨带优势植物茭草(Zizaniacaduciflora)、水葱(Scirpustabernaemontani)、黑三棱(Sparganiumstoloniferum)和杉叶藻(Hippurisvulgaris)凋落物为研究对象,基于政府间气候变化专门委员会(IPCC)对未来大气增温的预测,通过开顶式原位增温系统(Open-top Chambers,OTCs)模拟大气增温,研究大气增温处理下(1)植物凋落物在不同分解界面的质量衰减特征及其种间差异,(2)植物凋落物初始C/N值对植物凋落物质量衰减的影响,(3)大气温度、植物初始C/N值对凋落物衰减的差异影响特征。通过研究,拟掌握气候变暖对滇西北高原湿地植物凋落物分解的影响及其作用机制,为应对气候变化、加强该区域湿地生态系统碳汇功能的保护提供一定的理论依据。
1 材料与方法
1.1 区域概况
滇西北高原纳帕海湿地位于青藏高原东南缘横断山区、香格里拉市境内(99°37′ —99°43′ E,27°49 ′—27°55′ N),平均海拔3260 m,面积共计3100 hm2,为我国低纬度高海拔地区、在空间具有典型的“面山-湖滨-湖盆”生态结构特征的封闭与半封闭的季节性湖泊湿地类型[13,16]。纳帕海拥有丰富的生物多样性与特有性,分布有黑颈鹤(Grusnigricollis)、黑鹳(Ciconianigra)等国家I级保护鸟类,中甸叶须鱼(Ptychobarbuschungtienensis)等地方特有鱼类,具有极高的生物多样性保护价值。1984年,经云南省政府批准建立纳帕海省级自然保护区;2005年,被指定为国际重要湿地,成为国内外关注生物多样性保护的热点区域[12]。
纳帕海所在区域地处北温带气候带,属高原寒温性湿润气候,干湿季分明,其中,6—9 月份为雨季、10月—次年5月为旱季,年均降雨619.9 mm。每年雨季,纳帕海湖水上涨、水深可达4—5 m;到旱季,纳帕海湖水通过西北角落水洞潜流汇入金沙江,形成大面积的浅水沼泽和沼泽化草甸,平均水深不足20 cm。纳帕海全年均温5.4℃,最热月7月、月均温13.2℃,最冷月1月、月均温-3.7℃。
湖滨带植被是纳帕海湿地生态结构的重要组成部分,其优势植物是驱动着整个湿地生态系统物质循环与能量流动的基础[17]。前期研究表明,纳帕海湖滨带优势植物茭草、水葱、黑三棱、杉叶藻等挺水植物群落是该湿地生态系统碳收支平衡的基础[18]。
1.2 实验设计
1.2.1 开顶式生长室(OTCs)的设计及运行
2014年4月,在滇西北高原纳帕海湿地原位构建9个底面直径2.4 m的圆形研究单元,每研究单元通过取土形成35—40 cm的淹水环境,每3个研究单位为1种处理。基于IPCC对未来大气增温的预测,在其中2组处理的地面建立以阳光板为材料,高为2.4 m的开顶式生长室(OTCs),通过OTCs顶部开口的不同、分别设置大气增温(2.0±0.5)℃(EW-I)与(3.5±0.5)℃(EW-II)两种处理,每种处理3个重复,对照研究单元无OTC装置。所有研究单元通过深20 cm、宽20 cm水沟相通,保持各研究单元淹水环境一致。另外,根据实验区淹水条件的设计需要,可通过从附近的湿地引水以维持必要的淹水生境。
运用TP- 2200HumidityandTemperatureLoggers(固定在研究单元距水面1 m处)实时采集不同实验处理下的大气温度数据。温度数据监测频率设置为1次/h,每天24组数据。在实验结束时导出温度数据,并对所采集的温度数据进行整理分析(图1)。

图1 不同处理下大气温度的变化特征Fig.1 Mean daily and monthly temperatures under different experimental warming treatments during Dec. 2014 to Dec. 2015 其中, CK:对照大气温度,ambient temperature; EW-I:大气增温(2.0±0.5)℃ 处理,experimental warming (2.0±0.5) ℃; EW-II:大气增温(3.5±0.5)℃ 处理,experimental warming (3.5±0.5) ℃
2014年12月—2015年12月期间,CK、EW-I、EW-II处理下年平均温度分别为8.21、10.43、12.05℃,大气增温分别为2.22、3.84℃,符合大气增温实验设计要求。另外,在实验期间,CK、EW-I、EW-II处理下记录最高温分别为37.69、46.88、47.50 ℃,最低温分别为-22.69、-19.13、-16.75 ℃。
1.2.2 植物凋落物的制备与放置
本研究选取纳帕海湖滨带优势植物茭草、水葱、黑三棱和杉叶藻凋落物为研究对象,依据前期现地的观测,挺水植物凋落物可存于大气、水体,以及土-水界面中。因此,本研究将4种物种凋落物分别置于大气、水体、土-水3种分解界面予以研究。
2014 年 10月,在纳帕海湿地植物生长末期,对茭草、水葱、黑三棱、杉叶藻4种植物的地上部分进行收割,装袋后带回实验室。将收割获得的植物放置实验室,经1周的室内自然风干、并仔细剔除植物体的泥土等杂物后,放置烘箱(65 ℃)48 h,然后剪成10 cm 长的小段,并按物种将其混合均匀后制成凋落物。在室内称取茭草、水葱、黑三棱和杉叶藻4种物种凋落物分别装入编号的尼龙网袋(孔径1 mm,宽10 cm、长15 cm),每袋放入5.00 g凋落物。每物种、每分解界面放置凋落物分解袋45袋,4种植物、3种分解界面共计540袋。
2014年12月,将装有植物凋落物的分解袋放置于CK、EW-I和EW-II处理中,其中,大气界面悬于空中1.2 m处,水界面通过在凋落物袋中放入乒乓球以使凋落物袋悬浮于水中,土-水界面的凋落物袋则是通过PVC管将其固定在土-水界面。
1.2.3 植物凋落物有关参数的测定
2014年11月,在实验室分别测定4种植物凋落物初始的全碳(TC)和全氮(TN),其中,TC采用浓H2SO4-K2CrO4外加热法测定, TN采用浓H2SO4-H2O2加热后扩散法测定[19],测定结果如表1。

表1 4种湖滨带优势植物凋落物的初始TC、TN与C/N
2015年4月、6月、8月、10月和12月,分别将放置野外的植物凋落物分解袋取回实验室,每月、每处理、每物种、每界面分别取回凋落物袋共计108袋。将取回的凋落物放置实验室晾干一周,仔细剔除泥土等杂质,放置于65 ℃的烘箱48 h,用电子天平称重,分别计算各植物凋落物分解的质量衰减率、残留率、分解常数等指标。
质量衰减率用质量损失法计算:衰减率E%=(Mo-Mt)/Mo×100%;残留率W%=100%-E%;其中,Mt为凋落物分解后剩余质量(g),Mo为凋落物初始质量(g),t为凋落物分解时间。
利用Olson[20]指数衰减模型计算凋落物分解率常数k,即Mt=Moe-kt,其中,Mt为凋落物分解后剩余质量(g),Mo为凋落物初始质量(g),t为分解时间。
1.3 数据分析
利用SPSS 16.0统计软件,采用单因素方差分析(One-Way ANVOA)对不同大气增温处理、分解界面的植物凋落物质量衰减率、分解常数k进行差异性检验,显著性水平设定为P=0.05;用Canoco 4.5 软件的冗余分析,对凋落物分解常数k与凋落物初始C/N值和有关温度值进行相关性分析。
2 结果
2.1 凋落物在大气界面的质量衰减
2014年12月—2015年4月间,4种植物凋落物在大气界面的质量衰减量均较小,自2015年5月始,其质量减少的比例逐渐增加,其中,5—10月间的质量衰减量占年总衰减量80%以上。经过1年的分解,在CK、EW-I和EW-II处理下,茭草凋落物的年衰减率分别为(22.29±0.36)%、(33.56±0.15)%、(32.35±3.77)%;水葱年衰减率为(27.39±0.92)%、(29.43±0.19)%、(28.65±1.75)%;黑三棱年衰减率为(38.92±2.11)%、(47.63±3.52)%、(37.87±4.16)%;杉叶藻年衰减率为(58.39±0.01)%、(58.81±1.41)%、(56.52±5.31)%。其中,大气增温显著增加了茭草、黑三棱的质量年衰减率,而对水葱和杉叶藻凋落物衰减率的影响未达到显著水平。另外,不同植物凋落物的质量年均衰减率不同,其中, 在CK处理下,4种植物年衰减率由大至小依次为杉叶藻、黑三棱、水葱、茭草(F=393.079,P<0.001); 在EW-I处理下,黑三棱和杉叶藻的年衰减率显著高于茭草和水葱(F=200.936,P<0.001);而在EW-II处理下,杉叶藻的质量衰减率显著高于其他3种物种(F=39.304,P<0.001)(图2)。

图2 4种植物凋落物在大气界面不同增温处理下的质量残留率 (平均值±标准差) Fig.2 The remaining litter mass (%) (mean±S.E.) of the four plants under different experimental warming treatments at atmosphere interface
相对于CK,EW-I、EW-II增温显著增加了茭草凋落物分解常数(k值),而对水葱、黑三棱和杉叶藻凋落物分解常数的影响未达到显著水平(表2)。在相同大气增温处理下、不同植物凋落物分解常数k也存在差异,其中,在CK处理下,4种植物的分解常数k由高至低依次为杉叶藻、黑三棱、水葱、茭草(F=191.233,P<0.001);在EW-I、EW-II处理条件下,杉叶藻和黑三棱凋落物分解常数k显著高于茭草和水葱的分解常数(FEW-I=108.712,P<0.001;FEW-II=32.626,P<0.001)。
2.2 凋落物在水界面的质量衰减
相对于CK,大气增温对4种植物凋落物在水界面的质量衰减具有显著的促进作用,但对不同植物的影响存在一定差异。通过1年的分解,在CK、EW-I和EW-II处理下,茭草凋落物的质量衰减率分别为(57.76±1.75)%、(62.18±4.31)%、(67.86±0.01)%,水葱的质量衰减率分别为(59.81±0.88)%、(67.66±4.32)%、(72.95±4.98)%,黑三棱的质量衰减率分别为(54.05±1.58)%、(68.21±0.21)%、(71.82±1.29)%,杉叶藻的质量衰减率分别为(67.53±3.58)%、(78.17±0.49)%、(79.56±4.05)%,相同物种不同处理间均存在显著差异(FZC=12.301,P<0.01;FST=14.828,P<0.01;FSS=290.264,P<0.001;FHV=17.659,P<0.001)。另外,在相同大气温度处理下,杉叶藻凋落物衰减率高于其他3种植物(FCK=28.889,P<0.001;FEW-I=17.981,P<0.001;FEW-II=8.099,P<0.01)(图3)。
相对于CK,EW-I、EW-II处理显著增加了4种植物凋落物的分解常数k值。另外,在CK处理下,4种植物凋落物的分解常数k值差异不显著(F=2.585,P>0.05),在EW-I、EW-II增温处理下,杉叶藻的k值均高于其他3种植物(FEW-I=20.064,P<0.001;FEW-II=4.498,P<0.05)(表2)。
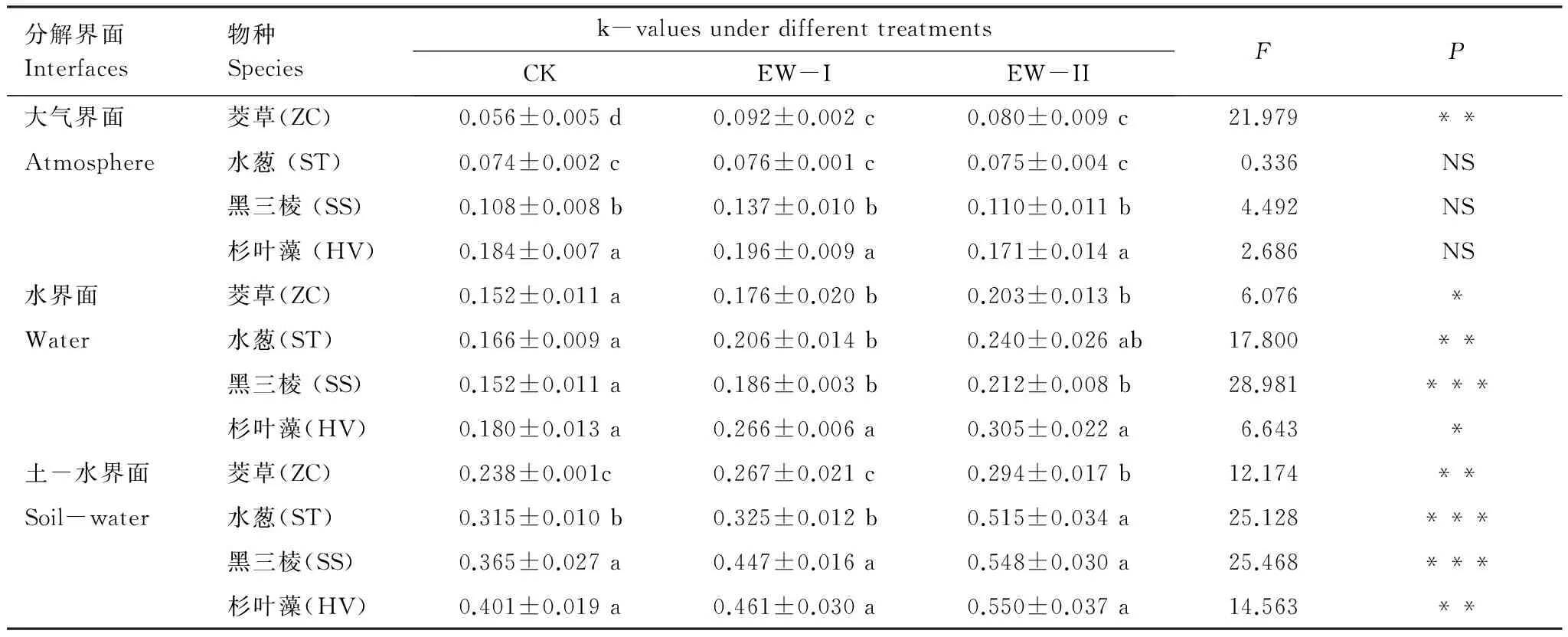
表2 不同增温处理下4种植物凋落物在不同分解界面的分解常数k值
茭草: ZC,Zizaniacaduciflora;水葱: ST,Scirpustabernaemontani;黑三棱: SS,Sparganiumstoloniferum;杉叶藻: HV.Hippurisvulgaris; NS: 无显著差异, no significant difference;P>0.05; *P< 0.05, **P<0.01, ***P<0.001; 相同分解界面、同列不同字母表示差异显著 (P<0.05)

图3 增温处理下4种植物凋落物在水界面的质量残留率(平均值±标准差)Fig.3 The remaining litter mass (%) (mean±S.E) of the four plants at the water interface under different experimental warming treatments
2.3 凋落物在土-水界面的质量衰减
大气增温对4种植物凋落物在土-水界面的质量衰减均有显著的促进作用。通过1年的分解,茭草凋落物在EW-II处理下的质量衰减率为(79.33±2.62)%,显著高于CK和EW-I处理下的质量衰减率(74.68±0.84)%、(76.28±0.47%)%(F=8.627,P<0.01);水葱凋落物在EW-II处理下的质量衰减率为(88.65±0.01)%,显著高于CK和EW-I处理下的质量衰减率(76.44±0.16)%、(76.94±2.01%)(F=48.167,P<0.001);黑三棱在EW-I和EW-II处理下的质量衰减率分别为(89.32±1.63)%和(93.52±0.01)%,显著高于CK处理下的质量衰减率(85.28%±1.70)(F=25.884,P<0.001);杉叶藻在EW-I和EW-II处理下的质量衰减率分别为(91.59±2.33)%和(95.00±0.98)%,显著高于CK处理下的质量衰减率(87.56±0.63)%(F=28.835,P<0.001)。另外,在CK处理下,4种植物凋落物质量年衰减率由大至小依次是杉叶藻、黑三棱、水葱、茭草(F=161.592,P<0.001);在EW-I处理下,黑三棱、杉叶藻的年衰减率显著高于茭草、水葱(F=86.245,P<0.001);在EW-II处理下,茭草的年衰减率显著低于其他3种植物(F=31.093,P<0.001)(图4)。
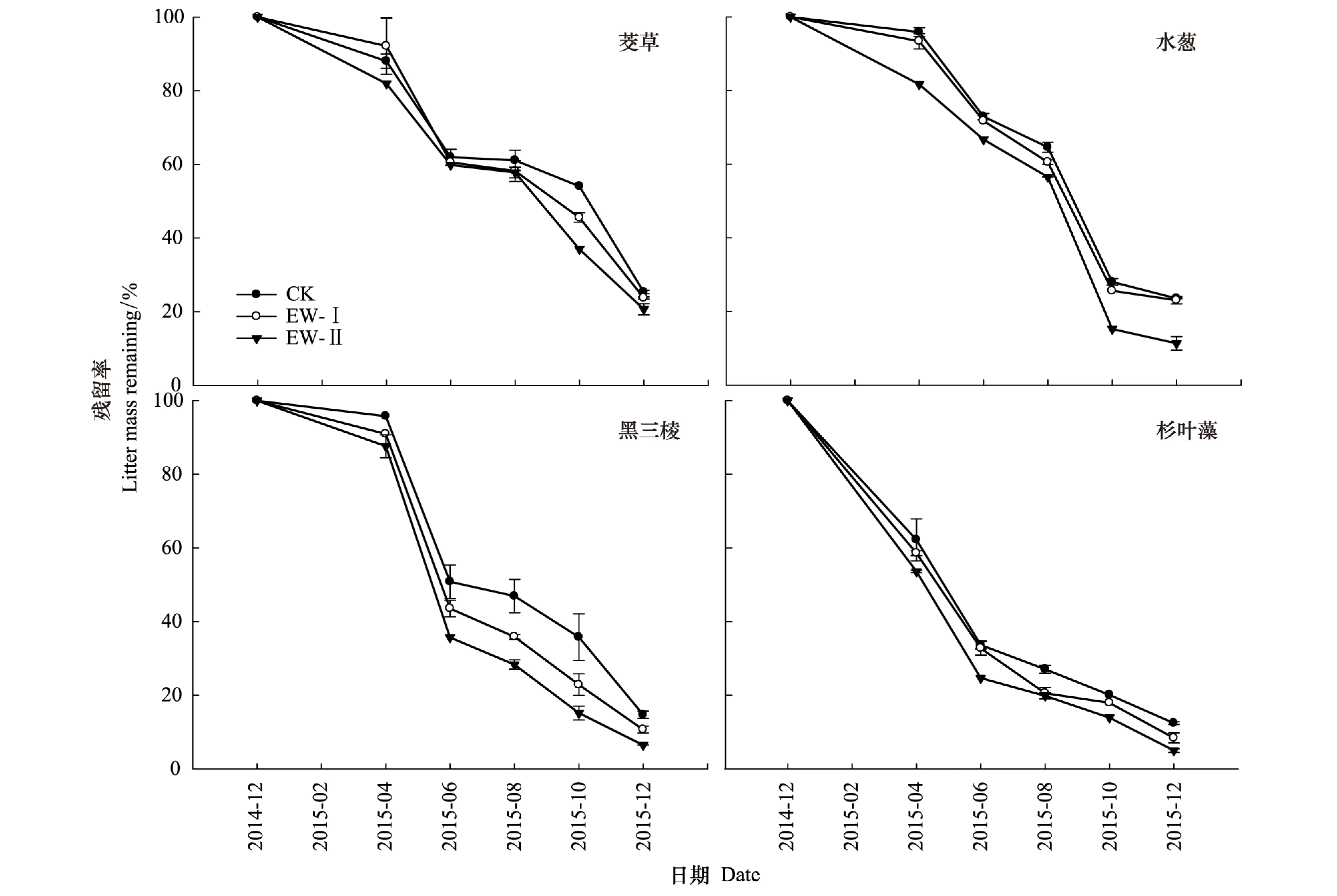
图4 增温处理下4种植物凋落物在土-水界面的质量残留率 (平均值±标准差) Fig. 4 The remaining litter mass (%) (mean±S.E) of the four plants under different experimental warming treatments in the below-water interface
EW-I、EW-II处理均显著增加了4种植物凋落物的分解常数k值,其k值随增温幅度的增加而增加。另外,在CK、EW-I处理下,杉叶藻和黑三棱凋落物分解常数k显著高于其他2种植物的分解常数(FCK=27.662,P<0.001;FEW-I=43.093,P<0.001);在EW-II处理下,茭草凋落物的分解常数k显著低于其他3种植物的分解常数(FEW-II=26.185,P<0.001)(表2)。
2.4 凋落物分解常数(k值)与温度、初始C/N值的相关性
在大气界面,植物凋落物初始C/N值与分解常数k值呈显著负相关,相关系数达0.88,温度对凋落物分解常数k值的影响不显著。在水界面和土-水界面,温度和凋落物初始C/N值与凋落物分解常数k值均存在相关性,其中,凋落物初始C/N值与凋落物分解常数k值呈负相关,温度值与分解常数k值呈正相关,但在水界面温度值对k的影响较大,而凋落物初始C/N值与凋落物分解常数的相关系数较小;而在土-水界面,温度与凋落物初始C/N值与凋落物分解常数k值相关系数均较高(表3)。
凋落物分解常数k值与温度值(最高温、最低温和年均温)、凋落物初始C/N值的冗余分析表明(图5),不同增温处理间的凋落物的分解速率沿X轴方向变化,解释了59.67%的变量,其中,EW-I和EW-II处理下的样品分别聚类在一起,与CK之间距离较远,表明初步增温导致凋落物分解加快,但随着增温幅度的增加对凋落物分解的影响逐渐减弱。不同物种的分解速率差异主要沿Y轴方向变化,解释了25.00%的变量,其中,茭草的分解速率明显不同于其他3个物种(水葱、黑三棱与杉叶藻),而其他3个物种之间的区分度不明显。统计分析进一步表明,凋落物初始C/N值是影响凋落物分解的主要因素(r=-0.72,P<0.001),呈负相关关系,即凋落物分解的速率随凋落物初始C/N值的增加而不断减小。温度是影响凋落物分解的次要因素(r=0.33,P<0.001),与分解速率之间成正相关,且主要受最低温度和平均温度的影响。
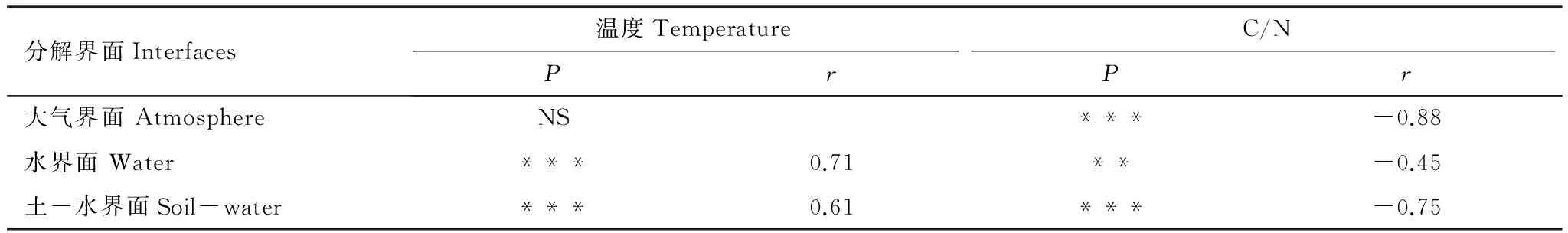
表3 温度值和C/N值对凋落物分解常数k的影响
NS=no significant difference,P>0.05; *P<0.05, **P<0.01, ***P<0.001
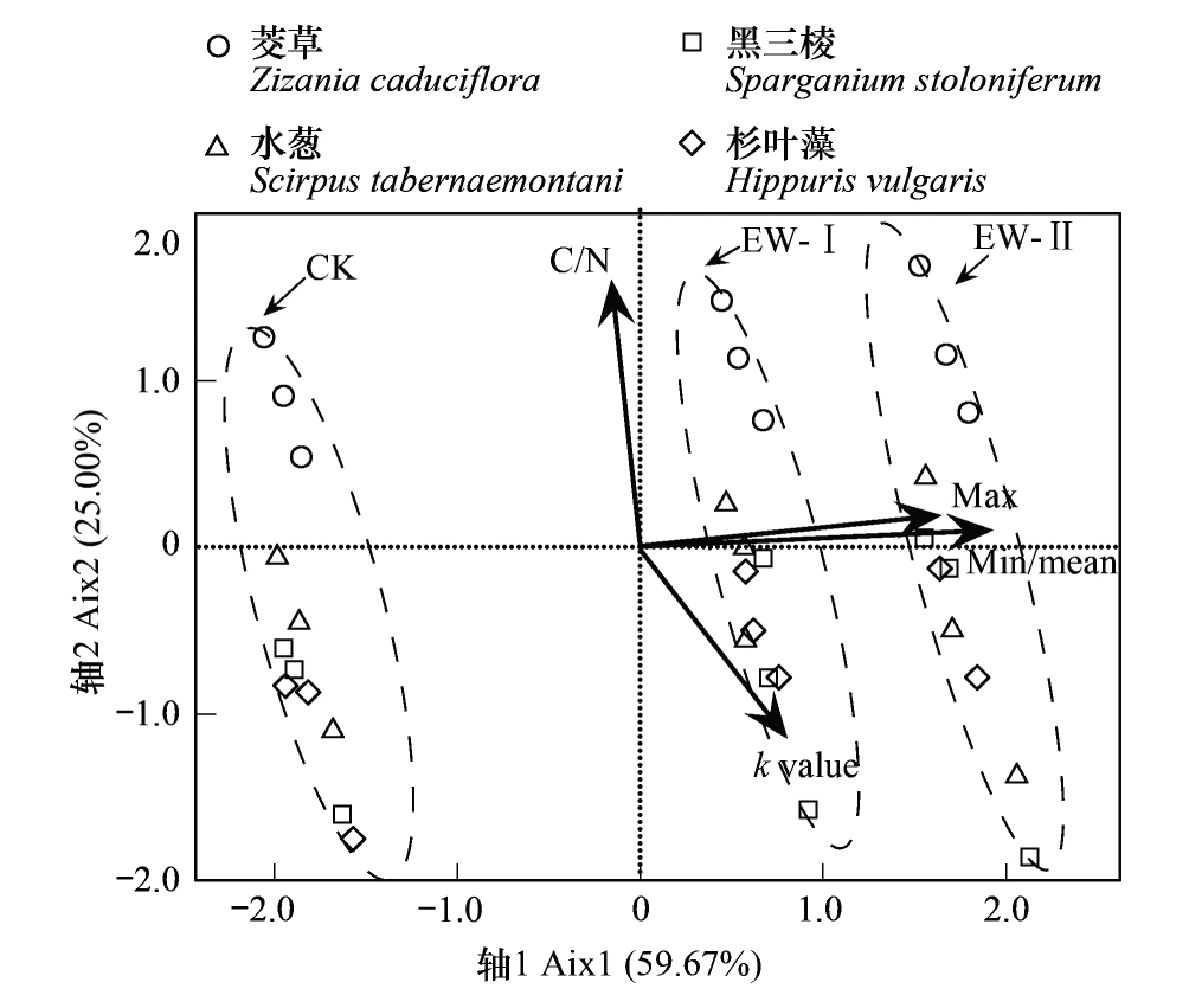
图5 植物凋落物初始C/N值和温度值对凋落物分解常数k的影响Fig.5 The impact of C/N of the plants and temperature to the k-values, Zizania caduciflora=ZC, Scirpus tabernaemontani=ST, Sparganium stoloniferum=SS, Hippuris vulgaris=HV
3 讨论和结论
植物凋落物的分解速率通常受分解者(微生物、无脊椎动物)活动的影响[4,21],而温度等环境因子则通过对微生物、无脊椎动物等分解者的活动以影响植物凋落物分解速率[4,22]。增温可有效增强微生物的酶促化学反应,从而加速微生物对凋落物中碳水化合物的分解[23],提高其对凋落物碳、氮利用率,从而加快植物凋落物的整体分解速率[24]。本研究中,大气增温对4种湿地湖滨带优势植物凋落物在不同分解界面的质量衰减均有一定的促进作用,其中,增温对大气界面植物凋落物质量衰减率的促进作用最小,其次为土-水界面,增温对水界面植物凋落物质量衰减率的促进作用最大。这与不同分解界面水、热条件,及其作用下的分解者活动的综合作用有关。在大气界面,由于香格里拉地区雨季较短,植物凋落物分解的水分条件较差,增温对大气中凋落物微生物的增殖与活动促进作用有限,同时大气界面缺乏土壤无脊椎动物活动对凋落物物理破碎过程,导致了增温对大气界面植物凋落物分解速率的促进作用最小。在水界面,植物凋落物分解的水环境条件最好,水体温度随着大气温度的增加而增加且水温总体稳定,水温升高加快了微生物的繁殖、增强了细菌和真菌等微生物的活性,提高了微生物对凋落物碳、氮等营养物质的利用效率,进而促进了植物凋落物的分解[25-27];另外,虽然水界面缺乏土壤无脊椎动物对凋落物的物理破碎过程,但是,由于纳帕海湿地水体在冬季结冰,春季快速增温将加速冰的融化过程而造成植物凋落物的物理破碎度增加,这有利于植物凋落物的分解。在土-水分解界面,由于该界面温度对大气增温的响应具有明显的滞后性,同时,土壤无脊椎动物种群的分布数量及活动受到水环境条件的限制,因此,大气增温对土-水界面植物凋落物分解率的影响相对于水界面小,但其分解速率随着增温幅度增加而增大的趋势显著。
植物凋落物自身特征在其分解中起关键作用[28],C/N值是影响微生物活性并调节凋落物营养动态平衡的重要指标[29],通常认为植物初始C/N值低的植物凋落物分解较快[30]。在本研究中,在不同增温处理下,4种湿地湖滨带优势植物凋落物的分解常数k值均与其初始C/N值呈显著负相关,即C/N值越高其凋落物分解越慢。其中,茭草凋落物初始C/N值显著高于水葱、黑三棱、杉叶藻,在增温与非增温处理下,其凋落物分解常数明显低于水葱、黑三棱与杉叶藻3种凋落物的分解常数,这与茭草凋落物中含较高比例的难分解的结构性化合物纤维素、半纤维素和木质素等有关。过去研究表明,凋落物中的N是促进早期分解的关键因子[31],在分解初期与分解速率正相关[25,32],但是,凋落物初始C/N值随着凋落物的分解而变化,进而改变了凋落物质量和分解元素的需求,初始C/N值对于凋落物分解的影响逐渐减弱,N在凋落物分解过程中的作用发生改变,在凋落物分解晚期与分解速率呈负相关[33-34]。本研究中,4种凋落物C/N值随着凋落物的分解而改变,但在大气变暖作用下,湿地湖滨带植物凋落物分解过程中C/N值动态变化如何影响其分解速率,不同物种间是否存在趋同或趋异响应,这有待深入研究。
通常情况下,增温与植物自身质量均对其凋落物分解存在一定影响,但其贡献率存在差异。本研究中,由于大气界面植物凋落物分解缺乏必要的水分条件,凋落物分解的温度与湿度协同促进作用在较大程度上被抑制,尤其增温处理进一步减少了大气界面的湿度,凋落物分解速率主要受物种自身质量的影响,增温对其分解速率的促进作用最小。在水界面,增温增加了微生物的活性[25],同时,冰融造成凋落物的物理破碎,这极大地促进了凋落物的分解。因此,增温通过改变微生物活性和凋落物物理性状而促进其分解,进而导致凋落物自身质量对分解率的作用减弱。相对其他两种分解界面,土-水界面不受水分限制与冰融作用的影响,凋落物的分解同时受到外界温度作用于微生物活性和植物自身质量的综合影响,因此,其分解常数与温度、植物初始C/N值的相关性均较高。另外,水葱、黑三棱与杉叶藻凋落物分解对温度的响应存在明显的趋同性,而茭草凋落物分解对增温的响应与这3种物种存在明显的趋异性,这种对增温的趋同与趋异响应机制需要深入探讨。此外,本研究也表明不同增温幅度对植物凋落物年分解率的影响存在差异,其促进作用随增温幅度的增加而明显降低,这可能与凋落物分解过程中微生物活性的温度适应范围有关。
滇西北高原湿地生态系统具有极为重要的碳汇功能,植物凋落物的分解与存留是其碳汇功能的基础。本研究进一步表明,滇西北高原湿地湖滨带优势植物凋落物分解同时受植物物种自身特征、分解界面以及大气温度的综合影响,其中,大气温度的增加是促进植物凋落物分解的重要外界因素,但其影响程度随着湿地植物物种的差异及其生境的不同而不同。因此,可以预见,未来气候变暖将促进该区域湿地生态系统植物凋落物的分解,进而对湿地生态系统物质循环及其碳汇功能产生影响。但是,由于湿地生态系统植物物种组成的多样性,以及湿地生境的复杂性,这决定了未来气候变暖对湿地生态系统碳汇功能影响的复杂性与不确定性,其相关作用机制有待于进一步研究。
[1] Hobbie S E. Plant species effects on nutrient cycling: revisiting litter feedbacks. Trends in Ecology & Evolution, 2015, 30(6): 357- 363.
[2] Fornara D A, Tilman D. Plant functional composition influences rates of soil carbon and nitrogen accumulation. Journal of Ecology, 2008, 96(2): 314- 322.
[3] Fussmann K E, Schwarzmüller F, Brose U, Jousset A, Rall B C I. Ecological stability in response to warming. Nature Climate Change, 2014, 4(3): 206- 210.
[4] 戴国华, 吕宪国, 杨青, 蒋海东, 武海涛, 朱美容. 三江平原湿地主要优势植物枯落物分解过程与能量动态研究. 湿地科学, 2008, 6(1): 29- 33.
[5] Bothwell L D, Selmants P C, Giardina C P, Litton C M. Leaf litter decomposition rates increase with rising mean annual temperature in Hawaiian tropical montane wet forests. PeerJ, 2014, 2(13): e685.
[6] Moore B, Braswell B H, Jr. Planetary metabolism: understanding the carbon cycle. Ambio, 1994, 23(1): 4- 12.
[7] Fierer N, Craine J M, McLauchlan K, Schmel J P. Litter quality and the temperature sensitivity of decomposition. Ecology, 2005, 86(2): 320- 326.
[8] Wang C Y, Han G M, Jia Y, Feng X G, Tian X J. Insight into the temperature sensitivity of forest litter decomposition and soil Enzymes in subtropical forest in China. Journal of Plant Ecology, 2011, 5(3): 279- 286.
[9] Cheng X L, Luo Y Q, Su B, Zhou X H, Niu S L, Sherry R, Weng E S, Zhang Q F. Experimental warming and clipping altered litter carbon and nitrogen dynamics in a tallgrass prairie. Agriculture, Ecosystems & Environment, 2010, 138(3- 4): 206- 213.
[10] Clarkson B R, Moore T R, Fitzgerald N B, Thornburrow D, Watts C H, Miller S. Water table regime regulates litter decomposition in restiad peatlands, New Zealand. Ecosystem, 2014, 17(2): 317- 326.
[11] Verburg P S J, Van Loon W K P, Lükewille A. The CLIMEX soil-heating experiment: soil response after 2 years of treatment. Biology and Fertility of Soils, 1999, 28(3): 271- 276.
[12] 肖德荣, 田昆, 张利权. 滇西北高原纳帕海湿地植物多样性与土壤肥力的关系. 生态学报, 2008, 28(7): 3116- 3124.
[13] 郭绪虎, 肖德荣, 田昆, 余红忠. 滇西北高原纳帕海湿地湖滨带优势植物生物量及其凋落物分解. 生态学报, 2013, 33(5): 1425- 1432.
[14] 李宁云, 袁华, 田昆, 彭涛. 滇西北纳帕海湿地景观格局变化及其对土壤碳库的影响. 生态学报, 2011, 31(24): 7388- 7396.
[15] Fan Z X, Bräuning A, Thomas A, Li J B, Cao K F. Spatial and temporal temperature trends on the Yunnan Plateau (Southwest China) during 1961- 2004. International Journal of Climatology, 2011, 31(14): 2078- 2090.
[16] 胡金明, 李杰, 袁寒, 董云霞. 纳帕海湿地季节性景观格局动态变化及其驱动. 地理研究. 2010, 29(5): 899- 908.
[17] 黄靖宇, 宋长春, 张金波, 郭跃东, 廖玉静. 凋落物输入对三江平原弃耕农田土壤基础呼吸和活性碳组分的影响. 生态学报, 2008, 28(7): 3417- 3424.
[18] 董瑜, 田昆, 郭绪虎, 肖德荣, 郭雪莲. 不同区域气候条件影响下的纳帕海湿地植物叶绿素荧光特性. 生态环境学报, 2013, 22(4): 588- 594.
[19] 鲍士旦. 土壤农化分析. 3版. 北京: 中国农业出版社, 2000: 12- 12.
[20] Olson J S. Energy storage and the balance of producers and decomposers in ecological systems. Ecology, 1963, 44(2): 322- 331.
[21] Gulis V, Ferreira V, Graca M A S. Stimulation of leaf litter decomposition and associated fungi and invertebrates by moderate eutrophication: implications for stream assessment. Freshwater Biology, 2006, 51(9): 1655- 1669.
[22] 宋新章, 江洪, 张慧玲, 余树全, 周国模, 马元丹, Chang S X. 全球环境变化对森林凋落物分解的影响. 生态学报, 2008, 28(9): 4414- 4423.
[23] Suseela V, Tharayil N, Xing B S, Dukes J S. Warming alters potential enzyme activity but precipitation regulates chemical transformations in grass litter exposed to simulated climatic changes. Soil Biology and Biochemistry, 2014, 75(1): 102- 112.
[24] 孙志高, 刘景双. 湿地枯落物分解及其对全球变化的响应. 生态学报, 2007, 27(4): 1606- 1618.
[25] Li J W, Ziegler S, Lane C S, Billings S A. Warming-enhanced preferential microbial mineralization of humified boreal forest soil organic matter: Interpretation of soil profiles along a climate transect using laboratory incubations. Journal of Geophysical Research Atmospheres, 2012, 117(G2): G02008.
[26] Wieder W R, Bonan G B, Allison S D. Global soil carbon projections are improved by modelling microbial processes. Nature Climate Change, 2013, 3(10): 909- 912.
[27] Schimel J, Balser T C, Wallenstein M. Microbial stress- response physiology and its implications for Ecosystem function. Ecology, 2007, 88(6): 1386- 1394.
[28] Ward S E, Orwin K H, Ostle N J, Briones M J I, Thomson B C, Griffiths R T, Oakley S, Quirk H, Bardgett R D. Vegetation exerts a greater control on litter decomposition than climate warming in peatlands. Ecology, 2015, 96(1): 113- 123.
[29] Güsewell, Gessner M O. N: P ratios influence litter decomposition and colonization by fungi and bacteria in microcosms. Functional Ecology, 2009, 23(1): 211- 219.
[30] 刘瑞鹏, 毛子军, 李兴欢, 孙涛, 李娜, 吕海亮, 刘传照. 模拟增温和不同凋落物基质质量对凋落物分解速率的影响. 生态学报, 2013, 33(18): 5661- 5667.
[31] Hobbie S E, Eddy W C, Buyarski C R, Adair E C, Ogdahl M L, Weisenhorn P. Response of decomposing litter and its microbial community to multiple forms of nitrogen enrichment. Ecological Monographs, 2012, 82(3): 389- 405.
[32] 葛晓改, 曾立雄, 肖文发, 黄志霖, 周本智. 三峡库区森林凋落叶化学计量学性状变化及与分解速率的关系. 生态学报, 2015, 35(3): 779- 787.
[33] Li L J, Zeng D H, Yu Z Y, Fan Z P, Yang D, Liu Y X. Impact of litter quality and soil nutrient availability on leaf decomposition rate in a semi-arid grassland of Northeast China. Journal of Arid Environments, 2011, 75(9): 787- 792.
[34] Aponte C, García L V, Maraón T. Tree species effect on litter decomposition and nutrient release in mediterranean oak forests changes over time. Ecosystems, 2012, 15(7): 1204- 1218.
EffectofexperimentalwarmingonthedecompositionoflitterfromdominantlakesideplantsinatypicalwetlandofNorthwesternYunnanPlateau,China
ZHANG Xiaoning, LIU Zhenya, LI Liping, WANG Hang, ZHANG Yun, SUN Mei, XIAO Derong*
NationalPlateauWetlandsResearchCenter,SouthwestForestryUniversity,Kunming650224,China
The impacts of climate change on the carbon cycle of wetland ecosystems have
considerable attention worldwide. However, the effects of global warming on the decomposition of litter from wetland plants, which is essential for anticipating the effects of climate change on the carbon source/sink function of wetlands, remains poorly understood. In the present study, experimentally examined the influence of atmospheric warming on plant biomass reduction at atmosphere, water, and soil-water interfaces, using open-top chambers that generated different warming conditions ((2.0±0.5) ℃ and (3.5±0.5) ℃) and four dominant lakeside plants (Zizaniacaduciflora,Scirpustabernaemontani,Sparganiumstoloniferum, andHippurisvulgaris) from the Napahai wetland, which is located on the Northwestern Yunnan plateau. The results revealed that (1) experimental warming promoted the mass decay of all four plant species at all interfaces, and the decay rate at the water interface was highest, followed by that at the soil-water interfaces and atmosphere, respectively. (2) The decomposition rates were correlated with the initial C/N ratio of plant litters, regardless of the experimental warming conditions, and the decay of biomass from plants with lower C/N ratios was more sensitive to warming than those with higher ratios. (3) Overall, litter decomposition was affected more by the initial C/N ratio than by the warming treatments. However, the present study still suggests that global warming could promote the decomposition of plant matter in wetland ecosystems in Northwestern Yunnan, with considerable consequences for the carbon cycle and ecological functioning of wetland ecosystems. Accordingly, studies should continue to examine the mechanisms underlying the effects of atmospheric warming on litter decomposition.
air warming; dominant lakeside plants; litter decomposition; Northwestern Yunnan Plateau
国家自然科学基金(31370497, 31500409);云南省高原湿地创新团队 (2012HC007)
2016- 10- 02; < class="emphasis_bold">网络出版日期
日期:2017- 08- 14
*通讯作者Corresponding author.E-mail: xiaoderong1@163.com
10.5846/stxb201610021988
张晓宁,刘振亚,李丽萍,王行,张贇,孙梅,肖德荣.大气增温对滇西北高原典型湿地湖滨带优势植物凋落物质量衰减的影响.生态学报,2017,37(23):7811- 7820.
Zhang X N, Liu Z Y, Li L P, Wang H, Zhang Y, Sun M, Xiao D R.Effect of experimental warming on the decomposition of litter from dominant lakeside plants in a typical wetland of Northwestern Yunnan Plateau, China.Acta Ecologica Sinica,2017,37(23):7811- 7820.
猜你喜欢
家庭影院技术(2021年8期)2021-11-02
数学年刊A辑(中文版)(2021年1期)2021-06-09
当代陕西(2020年13期)2020-08-24
制造技术与机床(2017年5期)2018-01-19
潍坊学院学报(2016年2期)2016-12-01
山西大同大学学报(自然科学版)(2016年4期)2016-11-27
河北书画研究(2016年2期)2016-08-24
新农业(2016年18期)2016-08-16
新高考·高一物理(2016年3期)2016-05-18
新闻传播(2015年11期)2015-07-18
