
大气增温对滇西北高原典型湿地湖滨带优势植物的光和CO2利用能力的影响
2018-01-04 05:33:41刘振亚张晓宁李丽萍肖德荣
生态学报 2017年23期
刘振亚,张晓宁,李丽萍,王 行,张 贇,孙 梅,肖德荣
西南林业大学国家高原湿地研究中心, 昆明 650224
大气增温对滇西北高原典型湿地湖滨带优势植物的光和CO2利用能力的影响
刘振亚,张晓宁,李丽萍,王 行,张 贇,孙 梅,肖德荣*
西南林业大学国家高原湿地研究中心, 昆明 650224
大气增温对湿地植物光合作用的影响及其作用机制是近年来生态学界关注的热点。采用开顶式生长室(Open-top chambers,OTCs)模拟大气增温((2.0 ± 0.5)℃,(3.5 ± 0.5) ℃),研究增温对滇西北高原典型湿地纳帕海湖滨带2种优势植物(茭草Zizaniacaduciflora,黑三棱Sparganiumstoloniferum)的光和CO2利用以及光合碳同化速率的影响。结果表明,(1)增温对不同植物的光和CO2利用能力以及碳同化速率的影响存在种间差异。增温显著降低了茭草的光饱和点(LSP)、光补偿点(LCP)、光能利用幅(LSP-LCP)、CO2饱和点(CSP)、CO2利用幅(CSP-CCP)以及最大净光合速率(Pmax),却显著增加了其CO2补偿点(CCP);相反,增温显著提高了黑三棱的LSP、(LSP-LCP)以及最大净光合速率(Pnmax),而显著降低了其LCP,但对其CO2利用参数无显著影响。(2)2种植物的光和CO2利用能力对增温的响应存在季节性差异。不同温度处理下,茭草在6、8月的LSP和(LSP-LCP)均显著高于10月的对应值,其Pnmax也随生长期的增加而降低;黑三棱的光响应参数在不同生长季间无显著差异,其Pnmax在8月最高,而在6月和10月相对较低。(3)温度因子与茭草的光能利用参数均呈负相关,而与黑三棱的光能利用参数均呈正相关。本研究的温度因子中,日间积温对Pnmax和(LSP-LCP),年均温和最低温对(CSP-CCP)值的影响最为显著。本研究进一步表明,气候变暖对滇西北高原湿地湖滨带优势植物光合作用存在影响,但不同物种间存在差异响应,这种差异响应可能导致湿地生态系统植被结构的改变,其有关生态过程有待于进一步研究。
滇西北高原;大气增温;湖滨带优势植物;光合作用
光合作用是植物生长和物质积累的基础,而植物对其进行光合作用所需要的光和CO2的利用是衡量植物光合效率的关键[1]。在植物光合作用过程中,光饱和点与补偿点,CO2饱和点与补偿点是植物对光能及CO2利用效率的集中体现[2]。
温度显著影响植物对光和CO2的利用效率,进而通过对植物光合作用的影响而影响其物质积累、生长、发育及种群扩散,导致整个生态系统结构与功能改变[3]。温度对植物光及CO2利用参数的影响存在差异。有研究表明,增温增加植物的光饱和点并降低其光补偿点,使得植物对光能的利用幅增大,促进植物的净光合速率[4];另有研究却表明,增温显著降低植物的光饱和点并显著增加其光补偿点,导致其光能利用幅度缩小[5]。也有研究认为,增温对植物的CO2补偿点和饱和点存在一定影响,但差异不显著[6],这与物种自身的差异密切相关[7]。另外,植物对增温的响应和适应存在一定的温度幅度[8]。一般情况下,在植物最适温度范围内,植物光合能力随环境温度的升高而增加,而当环境温度超过植物的最适生长范围时,增温会对植物的光合能力产生抑制作用,使得植物的光合速率降低[9-11]。也有少量研究认为,温度升高对植物光合作用参数的影响不显著[12- 13]。另外,植物的光和CO2响应,以及光合速率等与生长季密切相关[14]。研究表明,植物成熟叶往往比幼叶和过熟叶具有更高的光合碳同化能力、水分利用效率和光饱和点[14- 16],幼叶和过熟叶对环境变化较为敏感,而成熟叶的敏感度较低[14]。可见,增温对植物的光和CO2的利用能力,以及在此作用下的植物光合速率的影响尚存在众多争议。随着气候变暖对湿地生态系统影响的持续关注,湿地植物对气候变暖的响应成为了国内外研究的热点[17],但在全球范围内,其相关研究目前尚存在着许多不确定性。
滇西北高原湿地地处青藏高原东南缘,是我国“川滇生态屏障”的重要组成部分,对气候变化高度敏感[18]。由于滇西北高原湿地为断裂陷落形成的封闭与半封闭的独特湿地类型,具有典型的“面山-湖滨带-湖盆”结构特征,湖滨带是该类型湿地生态结构的基础,发挥着缓冲带、生物多样性保育和护岸等重要生态功能,也是响应气候变化的关键区域[19- 20]。湖滨带植物是维持湿地生态系统结构和功能的载体,研究其对气候变化的响应是掌握气候变化对该类型湿地影响的基础。自1960—2011年,滇西北高原地区大气年均温度增加了1.3℃,大气温度升高率达22.9%,其气候变暖的总体趋势与全球变化趋势一致,但显著高于云南其他地区[21]。随着滇西北高原地区大气温度的持续增加,必将对该区域湿地生态系统结构与功能产生深远的影响,进而对该区域湿地及其生物多样性的保护带来极大的挑战,但目前尚缺乏相关研究。
本研究选取滇西北高原典型湿地纳帕海2种湖滨带优势植物为研究对象,通过开顶式(Open-top chambers,OTCs)植物原位增温系统,研究了其(1)光能利用参数(光饱和点与补偿点),(2)CO2利用参数(CO2饱和点与补偿点),以及(3)最大净光合速率对大气增温的差异。研究拟掌握滇西北高原湿地植物光合生长对不同大气增温处理下的趋同与趋异响应特征,为科学评估气候变暖对滇西北高原湿地生态系统的影响奠定基础,为应对气候变化、加强该区域的湿地保护提供一定的理论依据。
1 材料与方法
1.1 研究区域
纳帕海湿地地处滇西北高原横断山脉中段香格里拉市境内(27°49′—27°55′N,99°37′—99°43′E),海拔3260 m,面积3100 hm2,为我国封闭与半封闭的独特湿地类型。纳帕海湿地属北温带高原寒温性湿润气候,受南北向排列的山地和大气环流的影响,全年盛行南风和南偏西风,干湿季分明,11月至次年5月为干季,降水少,6月至10月为湿季,降水充足。该区域太阳辐射较强,年日照数平均达2180.3 h,年均温为5.4 ℃,最热月7月均温为13.2 ℃,最冷月1月均温为-3.7 ℃。年平均降水量619.9 mm。≥10 ℃年积温为1392.8 ℃。
纳帕海湿地于1984年建立云南省纳帕海省级自然保护区,2005年被列入《国际重要湿地名录》。纳帕海分布有全球15种鹤类中唯一生活在高原的国家Ⅰ级保护鸟类黑颈鹤(Grusnigricollis)、国家I级保护动物黑鹳(Ciconianigra),以及滇西北高原特有鱼类中甸叶须鱼(Ptychobarbuschungtienensis)等,其生物多样性与特有性保护受到国内外的广泛关注。另外,纳帕海湿地也是众多植物物种分布的交汇区,植物种类丰富多样,其湖滨带分布有3个沉水植物群落、2个浮叶植物群落、6个挺水植物群落,其中,其湖滨带优势植物主要分布有茭草(Zizaniacaduciflora),黑三棱(Sparganiumstoloniferum)等[22]。
1.2 材料与方法
1.2.1 开顶式生长室(OTCs)的设计及运行
2014年4月,在滇西北高原纳帕海湿地建立9个原位研究单元,每个研究单元为直径2.4 m的圆形水池,各单元间相距3 m并通过PVC管连通,以保持各研究单元的淹水条件一致。基于IPCC对未来气候变暖的预测[23],将研究单元分为3组,每组3个重复,其中1组为对照组(CK),另外2组通过在其地面构建以阳光板为材料,底直径为2.4 m、高为2.4 m的开顶式生长室(Open-top chambers,OTCs),并依据开顶直径大小以实现研究单元大气分别增温(2.0±0.5)℃(experimental warming I=EW-I)和(3.5±0.5)℃(experimental warming II=EW-II))(图1)。

图1 滇西北高原纳帕海湿地开顶式生长室Fig.1 Open-top chambers (OTCs) in napahai wetland located at the northwest of Yunnan plateau
利用TP- 2200温度实时记录器(固定在研究单元距水面1 m处)对CK、EW-I和EW-II处理的大气温度进行实时记录,每小时记录1次、每天共记录24次,在实验结束时对温度数据进行导出处理。本研究时间段为2015年4月—2015年10月,相比于CK,EW-I与EW-II处理下的大气温度分别平均增温2.18 ℃和3.62 ℃,符合实验设计的需求(图2)。

图2 2015年4—10月不同处理下的大气日平均温度和月平均温度Fig.2 Mean daily and mean monthly air temperatures in different treatments from April to October, 2015CK=环境温度,EW-I=(2.0 ±0.5)℃增温,EW-II=(3.5±0.5)℃增温
1.2.2 供试物种的选取与移植
根据前期对纳帕海湖滨带优势植物研究的基础,本研究选取茭草(Zizaniacaduciflora)和黑三棱(Sparganiumstoloniferum)2种湖滨带优势植物为研究对象。于2015年4月,在纳帕海湿地分别挖取这2种植物足够的克隆单株,选取其长势基本相似的植株为实验对象,分别测定每克隆单株的高度与基径(表1)。通过单株盆栽、将植株分别移植到直径为35 cm、高为40 cm、内附湖滨带原位土壤(30 cm高)的塑料盆中进行培养。将移植了植株的塑料盆分别置入不同研究单元,每种植物、每研究单元放置5盆,即每个大气温度处理共计15盆。各研究单元淹水深度以其原生水淹环境为基准(20—30 cm)。自植株盆栽后,加强对植株的日常管理,以保证移植植株的成活。
1.3 光合生理指标的测定
分别于2015年6月、8月和10月,选择晴朗的天气,用美国产Li- 6400XT便携式光合作用测定仪,测定不同大气温度处理下2种植物的最大净光合速率(Pnmax),光饱和点(LSP)、光补偿点(LCP),CO2饱和点(CSP)和CO2补偿点(CCP)等生理参数,每研究单元每物种测定1—2株,每个温度处理下的每种植物测定总重复数不少于5次。
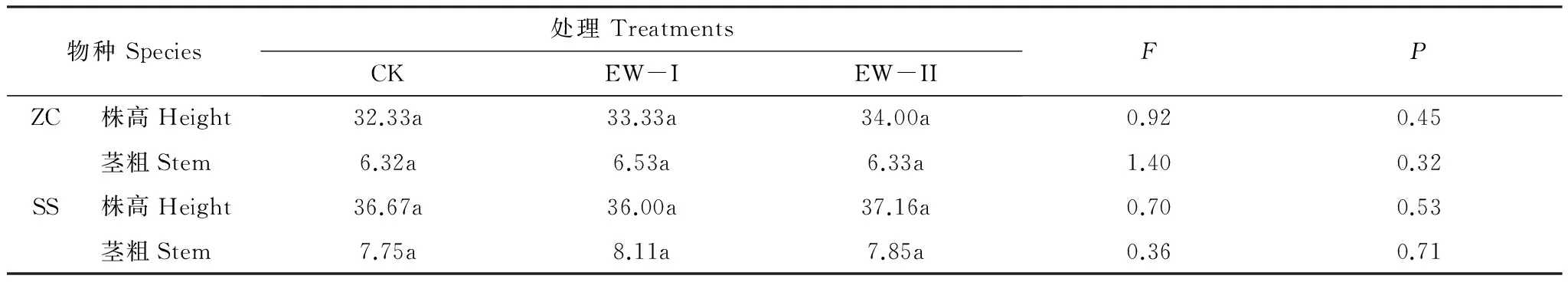
表1 不同温度处理下两种湖滨带优势植物的株高和基茎比较
其中,ZC:茭草,Zizaniacaduciflora; SS:黑三棱,Sparganiumstoloniferum;表中每一行的相同字母表示不同温度处理间差异不显著(P>0.05)
叶片净光合速率(Pn)、光饱和点(LSP)、光补偿点(LCP)的测定方法为[24]:使用开放气路,空气流速为0.5 L/min,叶片温度25 ℃,叶室中相对湿度70%—75%,CO2浓度400 μmol/mol。诱导光强约为2000 μmol m-2s-1,诱导时间30 min。测定时光强由强到弱依次设定光量子通量密度(PFD)为2200、2000、1900、1800、1700、1600、1500、1400、1300、1200、1100、1000、900、800、700、600、500、400、300、200、100、50、30、10、0 μmol m-2s-1。然后将野外测定的净光合速率数据通过软件Photosynthesis拟合光合-光响应曲线,并计算出植物的最大净光合速率(Pnmax)、光饱和点(LSP)、光补偿点(LCP)。
二氧化碳饱和点(CSP)与补偿点(CCP)的测定均在植物的光饱和点的条件下完成,使用开放气路,空气流速为0.5 L/min,叶片温度25 ℃,采用LI- 6400液化CO2钢瓶提供不同的CO2浓度,测定CO2浓度分别为400、300、200、100、50、400、600、800、1000、1200、1500、1800、2000、2200、2400、2600 μmmol/mol。将实测的净光合速率数据通过软件Photosynthesis拟合CO2响应曲线,并计算出CO2补偿点(CCP)、CO2饱和点(CSP)。
1.4 统计分析
采用SPSS统计分析软件包(SPSS 16.0,SPSS Inc., Chicago,IL, USA),通过单因素方差分析(One-Way ANOVA)、并采用LSD法进行多重比较,对不同大气温度处理下的植物最大净光合速率(Pnmax)、光饱和点(LSP)、光补偿点(LCP)、(LSP-LCP)值、CO2饱和点(CSP)、CO2补偿点(CCP)、(CSP-CCP)值进行差异性检验,统计显著性水平为P<0.05,极显著水平为P<0.01。根据植物生长期间的大气温度值,分别计算出年均温(Mean Year Temperature)、最高温(Max Temperature)、最低温(Minimum Temperature)、日间积温(Diurnal Cumulative Temperature)和夜间积温(Nocturnal Cumulative Temperature)等参数,将这些温度值与植物最大净光合速率(Pnmax)、(LSP-LCP)值、(CSP-CCP)值进行冗余分析(RDA),检测植物光合作用参数与温度变量之间的关系。
2 结果
2.1 光响应参数
相对于对照(CK),增温(EW-I、EW-II)降低了茭草的光饱和点(LSP)、光补偿点(LCP),以及光能利用幅(LSP-LCP)。在6月,其LSP、LCP、(LSP-LCP)值分别降低13.8%、28.9%,4.5%、19.8%,13.9%、29.1%,但不同处理间差异不显著(P>0.05);在8月,其LSP、LCP、(LSP-LCP)值分别降低42.3%、58.4%,3.6%、6.3%,42.5%、58.8%,3个处理间的LSP、(LSP-LCP)存在显著差异(P<0.05),但EW-I、EW-II间无显著差异(P>0.05);在10月,其LSP、LCP、(LSP-LCP)值分别降低41.2%、68.5%,16.7%、45.5%,41.8%、69.1%,3个处理间的LSP、LCP、(LSP-LCP)值均存在显著差异(P<0.05),但EW-I、EW-II间差异不显著(P>0.05)(图3)。另外,茭草的LSP、LCP和(LSP-LCP)值在不同生长季间也存在差异(图4)。CK、EW-I和EW-II处理下,茭草在10月的LSP和(LSP-LCP)均显著高于6月和8月(P<0.05),但在6月和8月间均不存在显著差异(P>0.05),而其LCP在3个生长季间均不存在显著差异(图4)。
相对于CK,增温增加了黑三棱的LSP和(LSP-LCP),但降低了LCP(图3)。在6月,其LSP、(LSP-LCP)分别增加了140%、185%,149.8%、235%,并达到显著水平(P<0.05),但EW-I、EW-II处理间不存在显著差异(P>0.05);在8月,其LSP、(LSP-LCP)分别增加了10.3%、21.3%,10.6%、22.3%,CK与EW-II处理间差异显著(P<0.05),而EW-I、EW-II间差异不显著(P>0.05);在10月,其LSP、(LSP-LCP)分别增加了18.9%、32.1%,20.9%和34.8%,并达到显著水平(P<0.05),但EW-I、EW-II处理间差异不显著(P>0.05)。在6月,其LCP分别降低了11%和36.8%,在8月,其LCP分别降低了4.9%和42.3%,3个处理间存在显著差异(P<0.05),但CK和EW-I间不存在显著差异(P>0.05)。在10月,其LCP分别降低了36%和40.5%,并达到显著差异水平(P<0.05),但EW-I、EW-II处理间差异不显著(P>0.05)。同样,黑三棱的LSP、LCP和(LSP-LCP)在不同生长季间也存在差异,其LSP和(LSP-LCP)值在8月最高,在6月和10月相对较低,其LCP值在8月最低,而在6月和10月相对较高。在CK处理下,其6月的LSP值显著低于8月和10月的对应值,而在EW-I、EW-II处理下,其LSP、LCP和(LSP-LCP)值在生长季间差异不显著(P>0.05)(图4)。
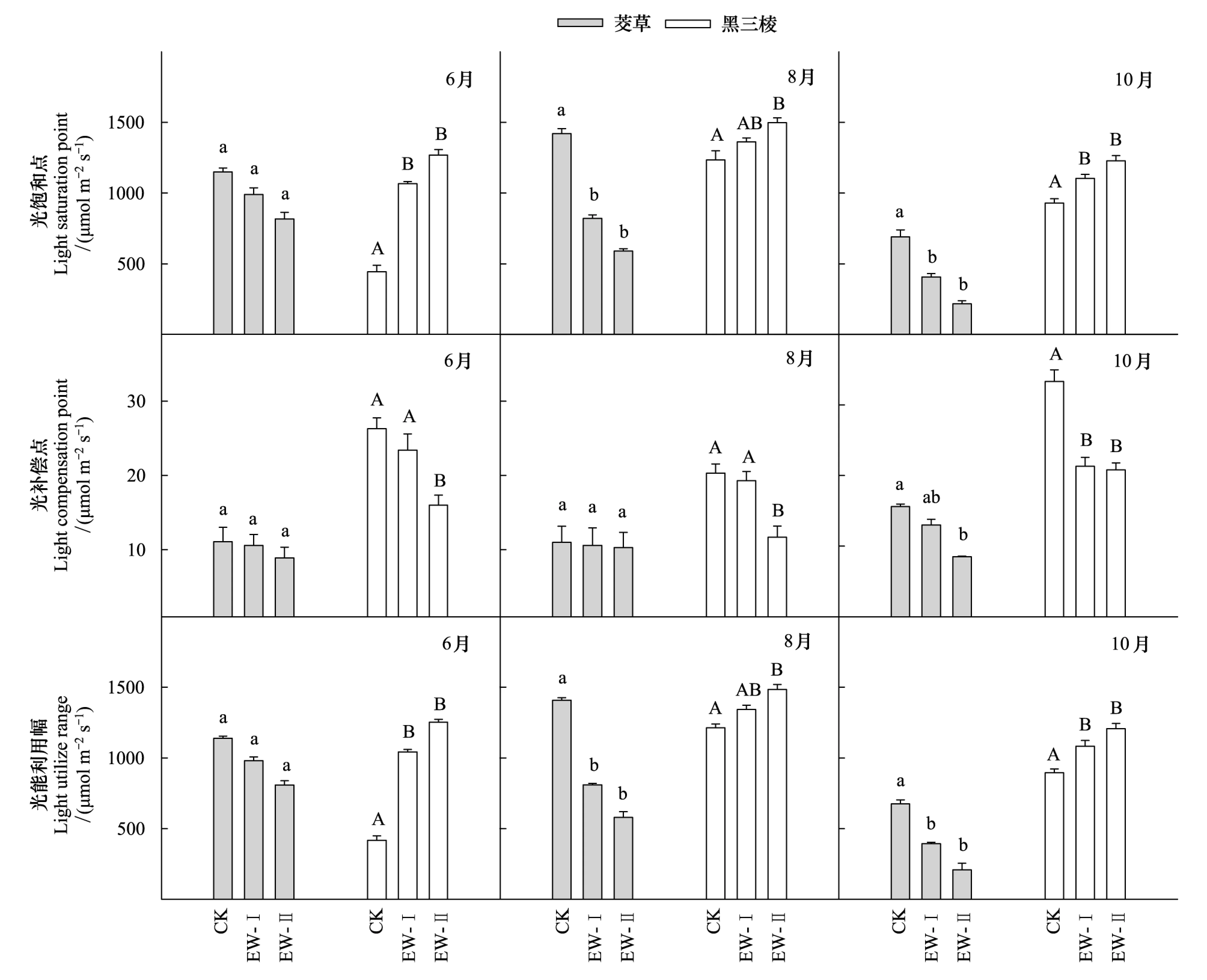
图3 不同生长季的茭草和黑三棱的光响应参数在三个不同温度处理间的差异Fig.3 Differences in light response parameters of Zizania caduciflora and Sparganium stoloniferum among three temperatural treatments in different growing stages图中每个物种对应的光响应参数柱上的不同字母代表差异显著(P<0.05),相同字母代表差异不显著(P>0.05);光响应参数包括光饱和点(light saturation point,LSP)、光补偿点(light compensation point,LCP)、光能利用幅(LSP-LCP)

图4 不同温度处理下茭草和黑三棱的光响应参数在3个生长季间的差异Fig.4 Differences in light response parameters of Zizania caduciflora and Sparganium stoloniferum among three growing stages in different temperatural treatments图中每个物种对应的光响应参数柱上的不同字母代表差异显著(P<0.05),相同字母代表差异不显著(P>0.05);光响应参数包括光饱和点(light saturation point, LSP)、光补偿点(light compensation point, LCP)、光能利用幅(LSP-LCP)
2.2 CO2响应参数
相对于CK,EW-I、EW-II降低了茭草的CO2饱和点(CSP)、CO2利用幅(CSP-CCP),但提高了CO2补偿点(CCP)(图5)。在6月,其CSP、(CSP-CCP)值分别降低1.3%、13.5%,1.3%、5%,3个处理间差异显著,但CK与EW-I间不存在显著差异;在8月,其CSP、(CSP-CCP)值分别降低7.6%、8.9%,7.9%、9.1%,3个处理间差异显著,但EW-I与EW-II间差异不显著;在10月,其CSP、(CSP-CCP)值分别降低1.6%和5.4%,1.7%和5.9%,3个处理间差异显著,但CK与EW-I间不存在显著差异。在6月和10月,3个处理间存在显著差异,但CK和EW-I间不存在显著差异。在8月,3个处理间存在显著差异,但EW-I、EW-II间不存在显著差异。在6月,其CCP分别增加了4.3%、18.6%;在8月,其CCP分别增加了11.3%、12.2%;在10月,其CCP分别增加了7.6%、45.9%,其中,在6月和10月、3个处理间存在显著差异,但CK和EW-I间差异不显著;在8月,不同增温处理下其CCP存在显著差异,但EW-I、EW-II间不存在显著差异。茭草在6月的CSP和(CSP-CCP)值显著低于其在8月和10月的对应值,而3个参数值在3个生长季间均不存在显著差异(图6)。
相对于CK,EW-I、EW-II增加了黑三棱的CSP和(CSP-CCP)值,却降低其CCP。在6月,其CSP、(CSP-CCP)值分别增加了2.5%、2.6%,2.9%、3%,在8月,分别增加了1.4%、5.3%,1.5%,5.6%,在10月,分别增加了0.6%、6.9%,0.7%、7.1%,但不同处理间均为达到显著差异。在6月,其CCP分别下降了20.9%、22.3%,3个处理间差异显著,但EW-I、EW-II间差异不显著;在8月,其CCP分别下降了7.5%、17.7%;在10月,其CCP分别下降了0.9%、9.7%,3个处理间不存在显著差异。黑三棱的CSP、CCP以及(CSP-CCP)在不同生长季间存在一定差异,其中,6月的CSP、CCP、(CSP-CCP)相对较高,在8月的值最低,但均未达到显著水平(图6)。
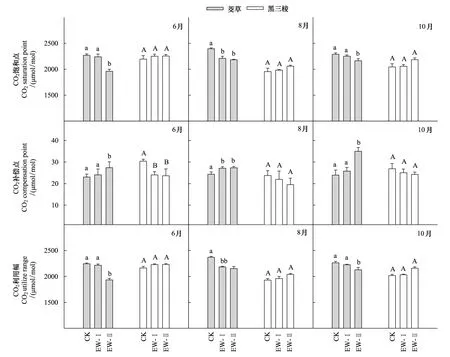
图5 不同生长季的茭草和黑三棱的CO2响应参数在三个不同温度处理间的差异Fig.5 Differences in CO2 response parameters of Zizania caduciflora and Sparganium stoloniferum among three temperatural treatments in different growing stages 图中每个物种对应的CO2响应参数柱上的不同字母代表差异显著(P<0.05),相同字母代表差异不显著(P>0.05);CO2响应参数包括CO2饱和点(CO2 saturation point, CSP)、CO2补偿点(CO2 compensation point, CCP)、CO2利用幅(CSP-CCP)
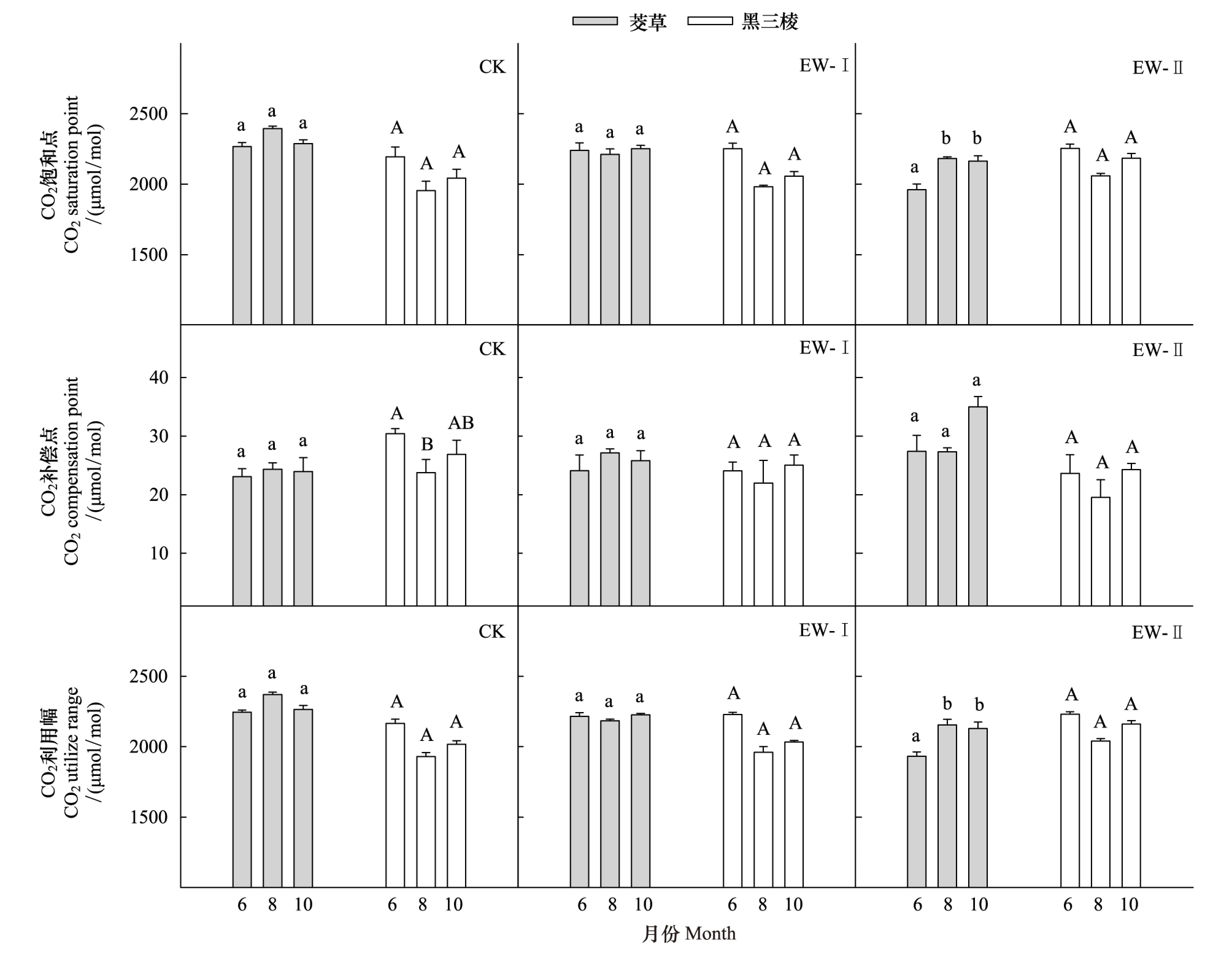
图6 3组温度处理下茭草和黑三棱的CO2响应参数在不同生长季间的差异Fig.6 Differences in CO2 response parameters of Zizania caduciflora and Sparganium stoloniferum among three growing stages in different temperatural treatments图中每个物种对应的CO2响应参数柱上的不同字母代表差异显著(P<0.05),相同字母代表差异不显著(P>0.05); CO2响应参数包括CO2饱和点(CO2 saturation point, CSP)、CO2补偿点(CO2 compensation point, CCP)、CO2利用幅(CSP-CCP)
2.3 净光合速率
相对于CK,EW-I、EW-II处理降低了茭草的最大净光合速率(Pnmax)。在6月,其Pnmax分别下降了6.5%和32.4%,3个处理间差异显著,但CK和EW-I间不存在显著差异;在8月,其Pnmax分别下降了37.6%和52.4%,不同处理间差异显著;在10月,其Pnmax分别下降了68.4%和80.9%,不同处理间存在显著差异。在相同处理下,茭草的Pnmax随着生长期的增加而呈降低趋势,在3个生长季均达到差异水平(图7)。
相对于CK, EW-I、EW-II处理增加了黑三棱的Pnmax。在6月,其Pnmax分别增加了100%和156%,3个处理下均存在显著差异;在8月,其Pnmax分别增加了12.4%和40.4%,CK与EW-II处理间差异显著;在10月,其Pnmax分别增加了2.2%和22.1%,EW-II分别于CK、EW-I存在显著差异。相同处理下,黑三棱在8月的Pnmax高于6月和8月的对应值,其中,EW-I、EW-II处理下、其在6月和10月的Pnmax值不存在显著差异(图7)。

图7 茭草和黑三棱的最大净光合速率分别在不同温度处理和不同生长季间的差异Fig.7 Differences in the maximum net photosynthetic rate (Pnmax) of Zizania caduciflora and Sparganium stoloniferum among three different temperatural treatments and three different growing stages图中每个物种对应的CO2响应参数柱上的不同字母代表差异显著(P<0.05),相同字母代表差异不显著(P>0.05)
2.4 温度与植物光合参数的相关性
增温对湿地湖滨带优势植物光合作用的影响存在种间差异(图8)。2物种的Pnmax、(LSP-LCP)、(CSP-CCP)值与年均温、最高温、最低温、日间积温、夜间积温的冗余分析(RDA)表明:各温度变量与茭草的Pnmax、(LSP-LCP)、(CSP-CCP)光合作用参数均为负相关,相关系数达0.926。其中,日间积温(Diurnal)对茭草Pnmax和(LSP-LCP)值的影响最为显著,而年均温(Mean)和最低温(Min)对其(CSP-CCP)的影响最显著。与茭草不同,黑三棱的Pnmax、(LSP-LCP)和(CSP-CCP)与各温度变量间均存在正相关(图8),相关系数达0.956。同样,日间积温对黑三棱Pnmax和(LSP-LCP)的影响最显著,年均温和最低温对其(CSP-CCP)的影响最显著。
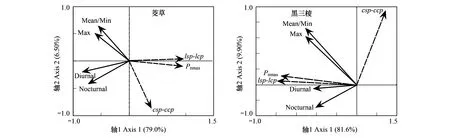
图8 冗余分析(RDA)示各温度参数与植物光合生理参数的相关关系Fig.8 The correlations between temperature factors and plant photosynthetic physiology parameters revealed by the RDA analysisMean:年均温,mean annual temperature;Max:最高温,max temperature;Min:最低温,min temperature;Diurnal:日间积温,diurnal temperature;Noctural:夜间积温,nocturnal temperature;光合生理参数包括最大净光合速率(Pnmax)、光能利用幅(LSP-LCP)、CO2利用幅(CSP-CCP)
3 讨论
3.1 增温对湿地湖滨带植物的光与CO2利用潜力的影响
植物的光饱和点(LSP)和光补偿点(LCP)是植物对光照条件要求的反映,与温度条件密切相关[25]。LSP提高和LCP降低分别增强了植物对强光和弱光的利用能力,能增加植物的光能利用幅(LSP-LCP),进而提高植物的光能捕获范围和光合能力,并增加同化量[26]。本研究中,增温显著降低了茭草的LSP、LCP和(LSP-LCP),表明在温度增加情况下其对光能的利用潜力减弱,这与赵娟等[4]、张利阳[5]等的研究结果一致。与茭草不同,增温显著增加了黑三棱的LSP、(LSP-LCP)值,但降低了LCP值,表明了增温处理下黑三棱利用光能的潜力增强。本研究进一步表明,增温对植物光响应参数的影响存在种间差异,这种种间差异可能与增温处理下植物体暗反应所产生的三碳化合物数量,以及植物捕获光量子的能力有关[27]。
温度也是影响植物对CO2利用能力的关键因子,其CO2饱和点(CSP)、补偿点(CCP),以及利用幅(CSP-CCP)等参数则反映了植物对大气CO2的捕捉、固定与利用能力[28]。本研究中,增温显著降低了茭草的CSP和(CSP-CCP),但显著增加其CCP,表明增温显著降低了茭草的CO2利用能力,随着未来大气CO2浓度的增加,茭草的生长将受到抑制,这可能与环境温度超过茭草对大气CO2利用的最适范围,热胁迫致使其活化的Rubisco酶减少以限制叶片光合潜力的发挥有关。与茭草不同,增温对黑三棱的CSP、CCP及(CSP-CCP)的影响均未达到显著水平,表明在未来的气候变暖下、黑三棱对大气CO2利用能力变化不大,这与徐振峰等[6]的研究结果一致,但与景立权等[6]的研究结果不同。
温度对植物光合速率的影响存在正效应[8, 29]、负效应[9]或无影响[30],与物种种类密切相关[31]。本研究中,大气增温使得茭草的Pnmax值呈现不同程度的降低,体现了增温对其光合作用的负效应,与祁秋艳等[9]和石福孙等[29]的研究结果一致,表明未来大气增温对茭草生长存在抑制作用。而黑三棱的Pnmax值则随温度的升高而上升,体现了增温对其光合作用的正效应,表明气候变暖有利于黑三棱的光合生长,这与Okimoto等[8]的研究结果一致。茭草与黑三棱光合速率对增温的差异响应也表明未来气候变化对湿地植物光合生长的影响存在复杂性和不确定性[26]。
3.2 植物的光和CO2利用潜力的季节性差异
植物的光合作用与物种和所处的生长季密切相关[14, 32]。本研究中,在相同大气温度处理下,茭草的LSP、(LSP-LCP)和Pnmax随着植物生长期的增加而呈现显著下降趋势,这种季节性差异可能是因为随着茭草叶龄增长,其对强光的利用能力减弱、光利用幅度降低等有关[32]。黑三棱Pnmax则随着生长期的增长而表现出上升趋势,这与众多有关陆生木本植物的研究结果一致[16]。
通常情况下,成熟叶光合速率显著高于幼叶,这主要是由于较高的电子流输送到羧化部位进行羧化作用,以及常绿叶片较低的光呼吸速率,或是因为成熟叶拥有较多的光系统(PSII)电子受体,或是成熟叶具有较高的比叶重和较低的叶绿素含量[14]。本研究中,茭草在10月(过熟叶)的光利用能力及光合速率较低,这可能与茭草过熟叶片的羧化受体和光合活化叶绿素含量下降有关,体现茭草叶片进行光合作用的时间相对较短,碳收支效率相对较低。相反,黑三棱叶片的光合作用时间较长,其叶片直至10月仍有较高的光合速率,表明黑三棱的碳利用效率较高。大气增温处理下,茭草与黑三棱光合作用的季节性差异及其形成机制有待深入研究。
3.3 温度对湿地湖滨带植物光合作用的影响
本研究中,温度因子与茭草的光和CO2利用参数以及净光合速率存在负相关,与黑三棱的光利用参数和净光合速率存在正相关。其中,日间积温是影响两种植物Pnmax和(LSP-LCP)的主要因素,这可能是由于植物的光合作用主要发生在白天,且日间积温是植物生长积温的决定因素有关[33]。最低温对两种植物(CSP-CCP)的影响最显著,这可能是因为最低温度通常发生在夜间,直接影响植物的酶活性而影响叶片在夜间的暗呼吸、碳消耗、CO2积累等。
植物的生长存在一定的温度适应范围。温度低于植物最适生长温度时,增温通过增加植物的呼吸动力来消耗叶片中的碳水化合物以增加库需(CSP-CCP增加),从而增加光合作用;当温度高于植物最适生长温度时,增温则可能抑制植物的暗呼吸消耗,表现为(CSP-CCP)下降。本研究表明,黑三棱的最适生长温度高于茭草的最适生长温度。基于此,随着未来的全球气候变暖,茭草的光和CO2利用能力逐渐降低,将导致其光合速率下降和有机物积累不足,其种群原有的优势地位将逐渐减弱。相反,黑三棱的最适生长温度较高,其光和CO2利用能力将随着气候变暖而呈增加趋势,其优势种的地位可能会得以维持。温度对湿地植物种间关系的影响及其作用机制有待深入研究。
4 结论
温度升高显著影响滇西北高原湿地湖滨带优势植物的光和CO2利用,以及碳同化能力,但存在种间差异。其中,增温降低了茭草的光和CO2的利用能力,以及净光合速率,但增加了黑三棱的光饱和点、光能利用幅和净光合速率。同时,相同温度处理下、两种植物的光和CO2利用能力表现出季节性差异,体现了湿地植物光合生长对不同生长季的适应策略。另外,温度因子与茭草的光能利用参数呈负相关,与黑三棱的光能利用参数均呈正相关,而日间积温对两种植物的Pnmax和(LSP-LCP),年均温和最低温对(CSP-CCP)值的影响最为显著。
[1] Brodribb T J, Holbrook N M. Forced depression of leaf hydraulic conductanceinsitu: effects on the leaf gas exchange of forest trees. Functional Ecology, 2007, 21(4): 705- 712.
[2] Sterck F J, Duursma R A, Pearcy R W, Valladares F, Cieslak M, Weemstra M. Plasticity influencing the light compensation point offsets the specialization for light niches across shrub species in a tropical forest understorey. Journal of Ecology, 2013, 101(4): 971- 980.
[3] Wright I J, Reich P B, Westoby M, Ackerly D D, Baruch Z, Bongers F, Cavender-Bares J, Chapin T, Cornelissen J H C, Diemer M, Flexas J, Garnier E, Groom P K, Gulias J, Hikosaka K, Lamont B B, Lee T, Lee W, Lusk C, Midgley J J, Navas M L, Niinemets Ü, Oleksyn J, Osada N, Poorter H, Poot P, Prior L, Pyankov V I, Roumet C, Thomas S C, Tjoelker M G, Veneklaas E J, Villar R. The worldwide leaf economics spectrum. Nature, 2004, 428(6985): 821- 827.
[4] 赵娟, 宋媛, 毛子军. 蒙古栎幼苗光合作用以及叶绿素荧光对温度和降水交互作用的响应. 北京林业大学学报, 2013, 35(1): 64- 71.
[5] 张利阳, 温国胜, 张汝民, 王电杰, 张俊. 毛竹光合生理对气候变化的短期响应模拟. 浙江农林大学学报, 2011, 28(4): 555- 561.
[6] 徐振锋, 胡庭兴, 张力, 张远彬, 鲜骏仁, 王开运. 青藏高原东缘林线交错带糙皮桦幼苗光合特性对模拟增温的短期响应. 植物生态学报, 2010, 34(3): 263- 270.
[7] 景立权, 赖上坤, 王云霞, 杨连新, 王余龙. 大气CO2浓度和温度互作对水稻生长发育的影响. 生态学报, 2016, 36(14): 4254- 4265.
[8] Okimoto Y, Nose A, Katsuta Y, Tateda Y, Agarie S, Ikeda K. Gas exchange analysis for estimating net CO2fixation capacity of mangrove (Rhizophorastylosa) forest in the mouth of river Fukido, Ishigaki Island, Japan. Plant Production Science, 2007, 10(3): 303- 313.
[9] 祁秋艳, 杨淑慧, 仲启铖, 张超, 王开运. 崇明东滩芦苇光合特征对模拟增温的响应. 华东师范大学学报: 自然科学版, 2012, (6): 29- 38.
[10] Liang J Y, Xia J Y, Liu L L, Wan S Q. Global patterns of the responses of leaf-level photosynthesis and respiration in terrestrial plants to experimental warming. Journal of Plant Ecology, 2013, 6(6): 437- 447.
[11] Niu S L, Li Z X, Xia J Y, Han Y, Wu M Y, Wang S Q. Climatic warming changes plant photosynthesis and its temperature dependence in a temperate steppe of northern China. Environmental and Experimental Botany, 2008, 63(1- 3): 91- 101.
[12] Llorens L, Peuelas J, Beier C, Emmett B, Estiarte M, Tietema A. Effects of an experimental increase of temperature and drought on the photosynthetic performance of two ericaceous shrub species along a North-South European gradient. Ecosystems, 2004, 7(6): 613- 624.
[13] Rosa R K, Oberbauer S F, Starr G, Parker La Puma, Pop E, Ahlquist L, Baldwin T. Plant phenological responses to a long-term experimental extension of growing season and soil warming in the tussock tundra of Alaska. Global Change Biology, 2015, 21(12): 4520- 4532.
[14] Chondrogiannis C, Grammatikopoulos G. Photosynthesis in developing leaf of juveniles and adults of three Mediterranean species with different growth forms. Photosynthesis Research, 2016, 130(1- 3): 427- 444.
[15] Hanba Y T, Miyazawa S I, Kogami H, Terashima I. Effects of leaf age on internal CO2transfer conductance and photosynthesis in tree species having different types of shoot phenology. Functional Plant Biology, 2001, 28(11): 1075- 1084.
[16] Andrianasolo F N, Casadebaig P, Langlade N, Debaeke P, Maury P. Effects of plant growth stage and leaf aging on the response of transpiration and photosynthesis to water deficit in sunflower. Functional Plant Biology, 2016, 43(8): 797- 805.
[17] Cronk J K, Fennessy S M. Wetland Plants: Biology and Ecology. Boca Raton, FL: CRC Press, 2001: 632.
[18] Erwin K L,Gardner R C. Wetlands and global climate change: the role of wetland restoration in a changing world. Wetlands Ecology and Management, 2009, 17(1): 71- 84.
[19] Richardson D M, Holmes P M, Esler K J, Galatowitsch S M, Stromberg J C, Kirkman S P, Pyšek P, Hobbs R J. Riparian vegetation: degradation, alien plant invasions, and restoration prospects. Diversity and Distributions, 2007, 13(1): 126- 139.
[20] 肖德荣, 田昆, 张利权. 滇西北高原纳帕海湿地植物多样性与土壤肥力的关系. 生态学报, 2008, 28(7): 3116- 3124.
[21] Fan Z X, Bräuning A, Thomas A, Li J B, Cao K F. Spatial and temporal temperature trends on the Yunnan Plateau (Southwest China) during 1961- 2004. International Journal of Climatology, 2011, 31(14): 2078- 2090.
[22] 肖德荣, 田昆, 袁华, 杨宇明, 李宁云, 徐守国. 高原湿地纳帕海水生植物群落分布格局及变化. 生态学报, 2006, 26(11): 3624- 3630.
[23] IPCC. Technical summary//Stocker TF, Qin D, Plattner G K, Tignor M, Allen SK, Boschung J, Nauels A, Xia Y, Bex V, Midgley P M. Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge, United Kingdom and New York, NY, USA: Cambridge University Press, 2013.
[24] 管铭, 金则新, 李月灵, 王强. 千岛湖次生林优势种植物光合生理生态特性. 生态学报, 2015, 35(7): 2057- 2066.
[25] Zhao C Z, Liu Q L. Growth and photosynthetic responses of two coniferous species to experimental warming and nitrogen fertilization. Canadian Journal of Forest Research, 2009, 39(1): 1- 11.
[26] Wang K Y, Kellomäki S, Laitinen K. Effects of needle age, long-term temperature and CO2treatments on the photosynthesis of Scots pine. Tree Physiology, 1995, 15(4): 211- 218.
[27] 徐兴利, 金则新, 何维明, 王兴龙, 车秀霞. 不同增温处理对夏蜡梅光合特性和叶绿素荧光参数的影响. 生态学报, 2012, 32(20): 6343- 6353.
[28] 黄滔, 刘炜, 唐红, 黄程前, 黄文韬, 唐军. 4个观赏竹种的光合特性及其影响因子分析. 植物资源与环境学报, 2016, 25(1): 24- 33.
[29] 石福孙, 吴宁, 吴彦, 王乾. 模拟增温对川西北高寒草甸两种典型植物生长和光合特征的影响. 应用与环境生物学报, 2009, 15(6): 750- 755.
[30] Lambrecht S C, Loik M E, Inouye D W, Harte J. Reproductive and physiological responses to simulated climate warming for four subalpine species. New Phytologist, 2007, 173(1): 121- 134.
[31] 张小全, 徐德应. 杉木中龄林不同部位和叶龄针叶光合特性的日变化和季节变化. 林业科学, 2000, 36(3): 19- 26.
[32] 陈凯利, 李建明, 贺会强, 胡晓辉, 姚勇哲, 孙三杰. 水分对番茄不同叶龄叶片光合作用的影响. 生态学报, 2013, 33(16): 4919- 4929.
[33] Yvon-Durocher G, Caffrey J M, Cescatti A, Dossena M, Giorgio P D, Gasol J M, Montoya J M, Pumpanen J, Staehr P A, Trimmer M, Woodward G, Allen A P. Reconciling the temperature dependence of respiration across timescales and ecosystem types. Nature, 2012, 487(7408): 472- 476.
InfluenceofsimulatedwarmingonlightandCO2utilizationcapacitiesoflakesidedominantplantsinatypicalplateauwetlandinnorthwesternYunnan
LIU Zhenya, ZHANG Xiaoning, LI Liping, WANG Hang, ZHANG Yun, SUN Mei, XIAO Derong*
NationalPlateauWetlandsResearchCenter,SouthwestForestryUniversity,Kunming650224,China
The influence of air warming on the photosynthesis of wetland plants and the underlying mechanisms has recently been one of the important topics in ecological research. Using the open-top chambers (OTC), we studied the influence of experimental air warming on the light and CO2utilization capacities and photosynthetic rate of two dominant species (ZizaniacaducifloraandSparganiumstoloniferum) on the lakeside of Napahai plateau wetland in northwestern Yunnan. The results showed that (1) the effects of warming on light and CO2utilization capacities were species-specific. ForZ.caduciflora, air warming significantly decreased the light saturation point, light compensation point, light utilization range, CO2saturation point, CO2utilization range, and maximum net photosynthetic rate, but it increased the CO2compensation point. In contrast, inS.stoloniferum, air warming significantly increased the light saturation point, light utilization range, and maximum net photosynthetic rate, but it decreased the light compensation point and weakly influenced the CO2utilization parameters. (2) The effect of air warming on the light and CO2utilization capacities of the two plant species showed seasonal variation. InZ.caduciflora, under the three temperature treatments, the light saturation point and light utilization range in October were significantly lower than those in June and August, and a seasonal decrease was observed in its maximum net photosynthetic rate. Contrarily, inS.stoloniferum, both the light and CO2utilization parameters were not influenced by air warming, but its maximum net photosynthetic rate was higher in August than that in June and October. (3) All temperature parameters were negatively correlated with photosynthetic parameters inZ.caduciflora, whereas they were positively correlated inS.stoloniferum. The diurnal temperature was the major factor influencing the maximum net photosynthetic rate and light utilization range, while the mean and minimum temperatures mainly affected the CO2utilization range. This study indicates that the lakeside dominant plants in northwestern Yunnan plateau show different adaptive strategies to air warming. In the context of continuous climate warming in northwestern Yunnan, the dominate status ofZ.caducifloramight get weakened, while that ofS.stoloniferummight get strengthened, and these changes, in turn, can induce alterations in vegetative components, structures, and functions of the whole ecosystem.
Northwestern Yunnan Plateau; air warming; lakeside dominant plants; photosynthesis
国家自然科学基金(31370497, 31500409);云南省高原湿地创新团队 (2012HC007)
2016- 10- 02; < class="emphasis_bold">网络出版日期
日期:2017- 08- 14
*通讯作者Corresponding author.E-mail: xiaoderong1@163.com
10.5846/stxb201610021989
刘振亚,张晓宁,李丽萍,王行,张贇,孙梅,肖德荣.大气增温对滇西北高原典型湿地湖滨带优势植物的光和CO2利用能力的影响.生态学报,2017,37(23):7821- 7832.
Liu Z Y, Zhang X N, Li L P, Wang H, Zhang Y, Sun M, Xiao D R.Influence of simulated warming on light and CO2utilization capacities of lakeside dominant plants in a typical plateau wetland in northwestern Yunnan.Acta Ecologica Sinica,2017,37(23):7821- 7832.
猜你喜欢
中等数学(2022年2期)2022-06-05 07:10:50
小学生学习指导(低年级)(2020年6期)2020-07-25 02:31:36
中学生数理化·高一版(2020年6期)2020-07-25 01:47:24
江苏农业科学(2020年12期)2020-07-20 03:26:38
西安科技大学学报(社会科学版)(2019年1期)2019-09-10 07:22:44
小学生学习指导(低年级)(2018年9期)2018-09-26 05:59:44
疯狂英语·新读写(2018年2期)2018-09-07 09:32:10
中学生数理化·八年级物理人教版(2017年3期)2017-11-09 03:05:33
新课程·中旬(2014年10期)2014-12-26 04:47:30
上海金属(2014年5期)2014-12-20 07:58:39
