
乒乓球和羽毛球运动员大脑可塑性差异比较
2017-12-06 06:30:17谭晓缨
中国体育科技 2017年6期
戴 雯,张 剑,谭晓缨
乒乓球和羽毛球运动员大脑可塑性差异比较
戴 雯,张 剑,谭晓缨
研究目的:长期系统化的运动训练会导致大脑运动皮层可塑性改变。在大脑层面上研究和分析项目间的特性,不仅能帮助运动员和教练更精确地了解专项技术特点,更能为训练效果的评价提供客观生理指标。研究方法:研究选用乒乓球和羽毛球项目中常用主动肌第1骨间背侧肌和桡侧腕伸肌,通过单脉冲经颅磁刺激刺激大脑初级运动皮层,产生运动诱发电位(MEP),以此评价大脑运动皮质兴奋性水平。结果:乒乓球运动员第1骨间背侧肌的MEP振幅和斜率显著高于对照组,羽毛球运动员桡侧腕伸肌的MEP振幅和斜率显著高于乒乓球组和对照组。结论:长期、系统的运动训练“有选择性”地增加主动肌肉对应区域的运动皮层兴奋性,能提高皮质内神经募集水平,帮助运动员更好地完成动作输出。
运动技能;经颅磁刺激;大脑可塑性;运动皮质兴奋性;运动诱发电位
1 前言
运动员专项技能的形成,离不开长期、高强度的专项训练。专项训练的结果之一就是技术的熟练性,其实质是大脑对肌肉控制能力的提高。大脑的这种变化通常称之为大脑运动皮层的可塑性。运动训练对大脑运动皮层可塑性的影响,主要体现在大脑运动中枢(初级运动皮层)激活的增加[3]。研究认为,这是由于长期重复的技能学习改变了神经突触的数量,加强了相应脑区之间的连接[10]。与之相对应,运动中枢激活的加强,又进一步提升了运动员对相关动作指令的加工和提取,帮助运动员更迅速和稳定地完成专项动作[33]。因此,结合运动专项研究大脑可塑性的变化,不仅能帮助运动员和教练更精确地分析项目特征,也为今后对运动员选材和专项训练效果的客观评价提供了理论基础。
经颅磁刺激(Transcranial Magnetic Stimulation,TMS)通过磁头线圈释放电流,刺激初级运动皮层(Primary motor cortex,M1),产生下行冲动电位,电位经皮质脊髓束到达脊髓运动神经元池,在对应肌肉中发生反应,形成运动诱发电位(Motor Evoked Potential,MEP)[2]。MEP振幅随TMS的刺激输出强度的增加而增大,这一变化不仅可以反映运动皮层-脊髓神经传导通路的兴奋性,还可以通过限定一定范围的刺激输出强度间接反映大脑运动皮层区神经元的激活情况[28]。该方法已成为大脑可塑性研究的常见电生理技术[14],国内、外研究通过这一技术探寻技能学习中运动皮质兴奋性和肌肉收缩的特性[9,34,35]。然而,当前该领域的研究主要聚焦于特殊技能群体(如运动员、音乐家、舞蹈演员等)皮质或脑区间的差异[24]。甚少有人分析不同专项运动员同一脑区对不同肌肉和关节控制的异同,并利用大脑可塑性的差异反过来印证专项间的差异。
结合目前TMS技术对上肢的高敏感性[36],研究选取了羽毛球和乒乓球进行比较。羽毛球和乒乓球虽同属隔网对抗性球类项目,对上肢的要求主要体现在灵活性和爆发力上,但两者的动作技术和球路变化各有其特点,动作的精确程度主要与上肢和手部小肌肉群特定部位的精细控制能力有关。单就上肢而言,羽毛球运动员在进行发球和跳杀扣球时主要运用手臂力量,由大臂带动小臂,通过肩-肘关节的力量完成快速抽击,主动肌包括桡侧腕伸肌、斜方肌及肱三头肌外侧头等[20,30]。乒乓球运动员则主要依靠手部力量,在同样达到快和准的情况下,还需要控制腕关节的旋转来实现球路的多变,主动肌主要涉及肘-腕关节,如第1骨间背侧肌、旋前圆肌及尺侧腕屈肌等[6]。由此可见,二者涉及的主要关节和主动肌群差异较大。研究拟以乒乓球和羽毛球为例,通过单脉冲TMS干预后的MEP变化趋势对比两组专项的运动员和普通人群的M1在桡侧腕伸肌(Extensor Carpi Radialis,ECR)及第1骨间背侧肌(First Dorsal Interosseous,FDI)上的大脑可塑性差异。
基于此,研究提出两个假设:1)长期高强度专项训练使运动员表现出更显著的皮层可塑性,主要表现在大脑对特定肌群运动控制能力的提高;2)运动员的大脑可塑性在运动皮层中呈现出与专项相关的“区域化”特点。
2 研究对象与方法
2.1 被试
实验选取被试共45人,每组均为15人。个人信息如表1所示,所有被试均为右利手、矫正视力正常、无任何精神病史。运动员组均是国家二级以上运动员,具有参加省级以上级别大赛的经验,其中,乒乓球组的所有被试均采用横拍打法。两组运动员的具体训练时长均由运动队统一安排,训练背景大致相同。对照组被试为普通大学生,没有进行过专业、系统的乒乓球、羽毛球或其他体育项目的训练。
研究通过了伦理委员会的审核,并签署知情同意书。被试在实验过程中有权随时中止实验。实验结束后给予相应报酬。

表1 被试基本信息Table 1 Subject Information
2.2 测试方法
研究选用两侧直径为9 cm的8字形线圈,连接至TMS刺激器(型号Magstim200),对左侧M1进行TMS刺激。刺激器通过Y形电缆与线圈相连,线圈的手柄几乎垂直于中央沟,并与大脑中央矢状线呈30°~45°,通过大脑的电流为“后-前方向”[28,29]。线圈放置在用最轻微的阈上刺激诱发出目标肌肉最大MEP振幅的区域,用专用笔在该区域进行标记并作为运动热点,通常情况下,FDI与ECR可共用同一个运动热点。
桡侧腕伸肌(ECR)和第1骨间背侧肌(FDI)的肌电数据使用直径9mm的Ag-AgCl表面电极记录。实验时将探测电极放置在右侧目标肌肉的肌腹处,参考电极放置在目标肌肉的远心端处,接地电极放置在手腕处。数据通过神经放电信号采集器(型号CED Micor1401)采集,采集到的信号使用电生理信号采集调节放大器(型号Model 2024F)进行过滤(带通滤波20~25 000 Hz),并放大1 000倍;过滤放大后的信号以1 kHz的频率被数字化记录,通过Signal 6.0软件进行线下分析。
2.3 实验流程
所有实验均于早上8:00-11:00在屏蔽信号的实验室中进行,整个过程安静且无人打扰。被试被安排坐在有舒服靠背的皮质座椅上,以使他们的右手前臂可以自然垂放,右手轻松置于座椅扶手上。
正式实验前首先确定每位被试FDI肌肉的静息阈值(Rest Motor Threshold,RMT)和运动阈值(Active Motor Threshold,AMT),并根据阈值确定运动热点。RMT指目标肌肉在静息状态下,连续10 次刺激中有至少5 次诱发出大于50 μV MEP的最小磁刺激强度;AMT指目标肌肉进行10%最大肌力收缩的前提下,连续10 次刺激中有至少5 次诱发出100~200 μV MEP的最小磁刺激强度[2]。
实验采用3×9×2(运动技能×刺激强度×肌肉部位)的混合设计。以运动技能因素作为组间因素(即乒乓球组、羽毛球组和对照组);以刺激强度和肌肉部位作为组内因素,其中:1)刺激强度根据事先定义好的基本刺激强度(SI1mV)进行调整,分别施予每位被试SI1mV强度的50%、70%、80%、90%、100%、110%、120%、130%及150%(共9种强度),并以此为基础建立每位被试的皮质脊髓输入-输出曲线(I-O曲线)[28]。SI1mV定义为施予FDI的刺激使运动诱发电位最接近1 mV时的强度,ECR测试同样选用该刺激强度(即不同肌肉条件下所对应的强度不变)。每种强度连续施予15次、每次间隔6~8 s。设置诱发潜伏期在15~35 ms间的第1次显著肌电波动为有效MEP,记录其波峰与波谷间的峰间值作为每次诱发的MEP振幅[26],取前10次有效MEP的振幅计算该强度的平均MEP振幅。强度顺序随机化,两次强度之间提供至少10 min的休息时间;2)肌肉部位分为FDI及ECR,同一肌肉的所有强度测试在1天内完成,两块肌肉的测试时间间隔至少48 h,每块肌肉的测试均重复进行1)中的操作。
2.4 数据分析与统计
数据以平均值±标准差呈现。研究将TMS刺激的前100 ms作为背景肌电基线(b.EMG)对整个诱发过程进行矫正、平滑和整合。图1以某位乒乓球运动员(男,19岁,训练年限11年)的FDI为例,显示了处理后的9个刺激强度所诱发出的运动电位。研究以运动技能作为组间因素,刺激强度作为组内因素,对不同肌肉条件分别进行重复测量方差分析,以检验MEP在不同肌肉条件下随强度变化的情况,同时使用单因素方差分析分别对每个强度下的3组被试的运动诱发电位进行检验,以探究不同专项运动员和对照组在哪些强度中存在显著差异。为简化I-O曲线数据,将根据已知数据所得出的线性回归直线的非标准化斜率定义为曲线坡度,并使用单因素方差对不同运动技能间的I-O曲线斜率进行分析。最后用皮尔逊积差相关来探讨斜率与训练总时长的关系,并进行线性拟合。数据的检验统计通过SPSS 17.0完成,置信空间设为P<0.05。
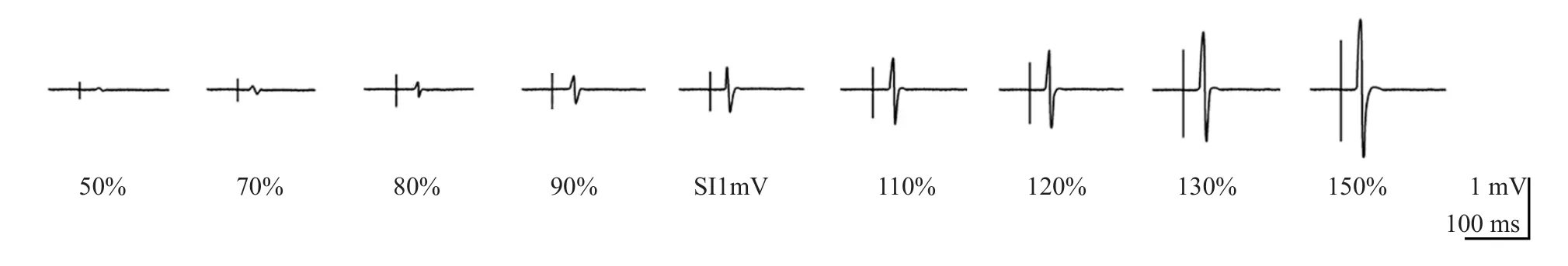
图1 不同强度的FDI运动诱发电位图示 (乒乓球被试,男)Figure 1. Example Recordings of FDI’s MEP in Di ff erent Stimulus Intensity (Subject of Table Tennis,Male)
3 研究结果
在所有实验中均没有被试提出不舒适感,也未见任何副作用。3组被试FDI的RMT、AMT以及诱发SI1mV所需的刺激输出强度间均无显著差异(RMT:F(2,44)=0.325,P=0.724;AMT:F(2,44)=0.057,P=0.944;SI11mV:F(2,44)=0.568,P=0.571)。不同组别间FDI的SI1mV差异同样不显著(FDI:F(2,44)=0.065,P=0.937)。乒乓球组和羽毛球组被试的训练年限之间也没有显著差异(=0.232,df=28,P=0.819)。
3.1 FDI皮层兴奋性变化情况
随着刺激强度的增减,被试的MEP振幅也有相应的增减。图2表示3组被试FDI肌肉的I-O曲线趋势。3组被试(乒乓球、羽毛球和对照组)的MEP振幅主效应(F(8,35)=494.014,P<0.001,η2=0.991)和组间效应(F(2,42)=5.203,P=0.010,η2=0.199)均显著。事后检验表明,乒乓球和对照组之间的MEP振幅存在显著差异(P=0.007),但乒乓球—羽毛球,羽毛球—对照组之间均没有显著差异(P>0.05)。
对9种刺激强度分别进行比较发现,3组被试在较高或较低的70%(F(2,44)=4.950,P=0.012)、120%(F(2,44)=13.314,P<0.001)、130%(F(2,44)=5.324,P=0.009)和150%(F(2,44)=9.667,P<0.001)强度时均存在显著差异。在最低强度50%时没有显著差异,可能是由于数值太小所导致。
此外,研究还计算了各组I-O曲线的斜率来评价兴奋性的增长幅度。图2表示3组被试FDI肌肉的I-O曲线斜率。3组被试间存在显著差异(F(2,42)=13.180,P<0.01)。事后检验表明乒乓球组和羽毛球组的斜率均显著大于对照组(乒乓球组-对照:P<0.001;羽毛球组-对照:P=0.027),但两组运动员之间差异不显著(P=0.064)。
3.2 ECR皮层兴奋性变化情况
与FDI增减趋势一样,ECR的MEP振幅也会随着刺激强度的增减而增减。图3表示3组被试ECR肌肉的I-O曲线趋势和斜率。3组被试(乒乓球、羽毛球和对照组)的MEP振幅主效应(F(8,35)=83.356,P<0.001,η2=0.950)和组间效应(F(2,42)=16.211,P<0.001,η2=0.436)均显著。事后检验表明,羽毛球与另外两组之间的MEP振幅均存在显著差异(P<0.001),但乒乓球和对照组之间的差异不显著(P>0.05)。
同样,对3组被试ECR肌肉的各强度MEP振幅分别进行比较后发现,与FDI的增长趋势不同,羽毛球的ECR要远远大于乒乓球和对照组。3组被试在70%(F(2,42)=4.974,P=0.012)、80%(F(2,42)=14.373,P<0.001)、90%(F(2,42)=10.958,P<0.001)、100%(F(2,42)=7.703,P=0.001)、110%(F(2,42)=7.520,P=0.002)、120%(F(2,42)=10.414,P<0.001)、130%(F(2,42)=15.949,P<0.001)、150%(F(2,23)=13.072,P<0.001)强度时均存在显著差异。
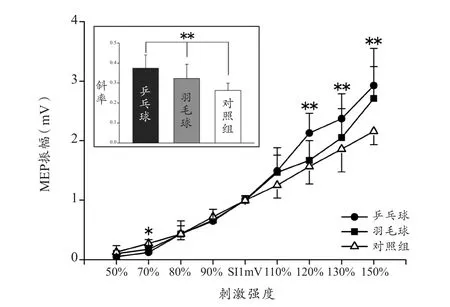
图2 3组被试FDI肌肉的I-O曲线趋势和斜率Figure 2. Trend and Slope of Input–Output Curve in FDI among Three Groups
此外,研究同样也计算了各组被试ECR上的I-O曲线的斜率来评价兴奋性的增长幅度。图3表示3组被试ECR肌肉的I-O曲线斜率。三组被试间存在显著差异(F(2,42)=12.558,P<0.01)。事后检验表明羽毛球组的斜率显著大于乒乓球组和对照组(羽毛球组-乒乓球组:P=0.001;羽毛球组-对照组:P<0.001),但乒乓球和对照组之间差异不显著(P>0.05)。
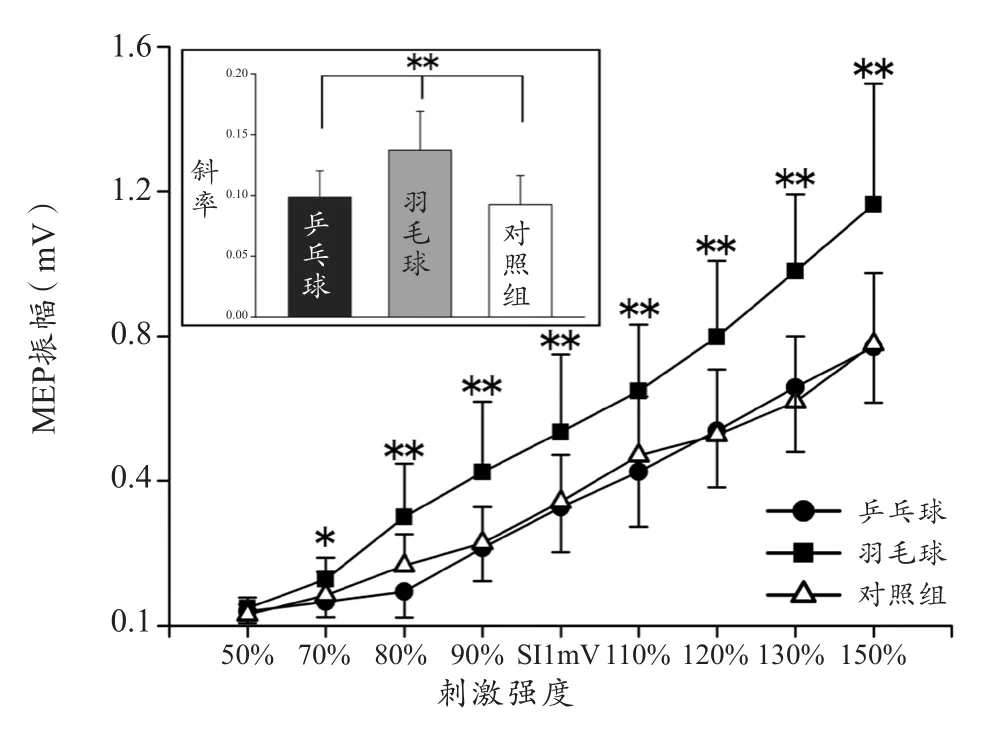
图3 3组被试ECR肌肉的I-O曲线趋势和斜率Figure 3. Trend and Slope of Input–Output Curve in ECR among Three Groups
4 讨论
MEP可以评估运动皮质脊髓束募集到的锥体神经元,通过椎体神经元对肌肉的激活程度来评价大脑运动皮层的可塑性[23]。I-O曲线是用来测试皮质脊髓投射强度的方法[5],斜率的变化依赖于运动皮层和脊髓中受到激活的运动神经元和中间神经元的分布情况[28],斜率越大说明受激活的神经元分布越广。本研究在AMT、RMT以及SI1mV中均未发现刺激输出强度的显著差异。这表明了在该TMS范式可测量范围之内,3组被试皮质脊髓神经元的最大激活能力以及运动皮层-脊髓的传输能力并没有因为运动员的长期训练存在差异。
4.1 运动员和对照组运动皮层可塑性的差异
本研究发现,经过长期专业训练的运动员M1区域的MEP振幅显著大于对照组。这与前人“单一阶段的运动技能学习会促进运动诱发电位的提高”的观点相一致[7,27]。随着刺激的增加,运动员I-O曲线的递增幅度逐步加大。研究中运动员的I-O曲线斜率显著高于对照组,说明运动员的运动皮层兴奋性更大,皮质内的募集水平更高,同时预示了长期训练所造成的皮层可塑性变化主要体现在M1功能上的重组。研究认为,MEP振幅的增加可能与大脑运动中枢(M1)中现有突触的连接加强有关。突触连接的加强在一定程度上可以加速大脑对运动信息的编码以及对相关技能的提取,帮助运动员在复杂多变的环境和条件下更加稳定和快速地做出所需要的反应。
相关的动物实验表明,运动训练会使已经发育完全的运动皮层形成新的突触连接,这些连接不会直接影响技能的获取,但会在运动技能学习中起巩固作用[4,18]。运动员在学习或专精于某项技能时,恰恰需要通过反复大量的练习来进行巩固和变通。运动员和新手之间最大的区别在于,尽管人类擅长通过模仿,短时间掌握某个技能动作,但只有经过长期反复的练习才能在复杂多变的环境和条件下,从大量技能动作中更快速、准确地选取最合适的动作,做出适当的反应。研究表明,在学习的后期阶段,“运动-感觉”网络的连通更加稳固[14,21]。这一阶段中, M1作为大脑的运动中枢,主要控制运动的稳定性和灵敏性[8],将多样化的肌肉、关节等运动参数整合在一起,并通过神经网络集成连贯动作完成输出[12,32]。也就是说,对于已经熟练掌握了多个单一、基础动作的个体而言,M1实际上是一个融合器,灵活调节和重组所学习到的各个运动组块,形成新的运动图式以达到最优化地技能输出[17]。因此,研究认为,M1在专家和新手中所起的作用存在一定程度的差异,而这种差异则可能是导致运动员的MEP振幅更大的主要原因。
4.2 运动皮层可塑性对特定肌群运动技能控制的影响
达成熟练的运动技能需要重组M1中受到激活的皮质脊髓神经元,提高这些神经元的神经募集水平[27]。神经募集水平指1次神经冲动能引起的相应肌纤维收缩量,募集水平高意味着M1发出的神经冲动更高效,从而更精确地控制运动技能。本研究利用SI1mV限定了刺激输出强度,仍然发现,乒乓球运动员FDI的MEP振幅在距离SI1mV刺激强度较远的120%~150%(高强度)和70%(低强度)中均显著高于其他两组;而羽毛球运动员ECR的MEP振幅在70%~150%刺激强度中始终显著高于其他两组。这不仅表明了运动员在特定肌群中体现出了更显著的运动皮层可塑性,还表明了运动员具有更敏感的神经募集水平——无论是高强度还是低强度的外界刺激都能更有效地激活皮质脊髓神经元。这种变化可以帮助运动员更好地完成运动技能。然而,研究在50%强度时未发现显著差异,分析原因可能是由于部分被试所接受的50%刺激强度已低于静息阈限,这也说明皮质脊髓神经元的激活与阈上刺激有关,排除了运动员自身潜在的阈下变化。
由此可见,熟练的运动技能使运动员的M1产生可塑性变化,对外界刺激更加敏感;反之,M1的高效率编码也会促进运动技能的输出,帮助运动员更快速也更稳定地在众多运动组块中对应外界条件选取正确的反应。这种变化在技能相关肌群中尤其显著。“M1-运动输出-M1”在运动技能学习中是一个良性循环,相辅相成。因此,研究认为,探究M1的活性和兴奋性也可以在一定程度上评价某一运动技能的熟练程度。
4.3 项目特点对运动皮层可塑性的影响
尽管许多神经影像学研究都发现了特定的运动锻炼会激活更多与运动任务区域相关的神经[1,11,31],但很少有专门就不同专项的特定运动区进行深入研究。研究结果表明,羽毛球运动员的ECR兴奋性显著高于乒乓球运动员和对照组,与之相对,乒乓球运动员的FDI则显著高于对照组。在运动员的训练过程中,对主动肌肉的侧重点不同是导致这一结果的主要原因:乒乓球和羽毛球虽然都是隔网球类运动,其上肢的运动模式却是大相径庭。单就上肢而言,乒乓球在讲究击球速度的“快”和“准”外,更注重球路变化。乒乓球运动中,前臂增加球的速度,针对前臂主要进行力量训练,而手腕加大球的旋转以达到球路多变,针对手腕以敏捷性训练为主。羽毛球则主要靠击球力量和出球速度取胜,上肢训练也以力量训练和爆发力训练为主,侧重上臂整体力量传递的连续性,通过大臂和小臂的配合,达到力量的最大输出。从这一点来看,我们发现,乒乓球训练的关键点在于腕关节连接的小臂和手部动作的改变,而羽毛球训练的关键点在于肘关节连接的大臂和小臂的配合。总的来说,乒乓球训练侧重肘关节力量到腕关节灵活性的转变,FDI受腕关节支配,是其运动模式中的主动肌肉[15];羽毛球训练则侧重肩关节到肘关节的整体瞬时发力,ECR伸展并带动肘部弯曲完成动作[22]。二者技术特性上的差异也是导致大脑可塑性不同的主要原因。此外,在FDI的实验中,与对照组相比,羽毛球组表现出了和乒乓球组一样的高兴奋性,尽管它的兴奋趋势和斜率都低于乒乓球组。研究者认为,这可能是因为二者均为持拍运动,在握拍和控拍时都运用到了FDI肌肉。因此,研究认为,两种球类运动对上肢的要求不同以及运动模式之间的差异是导致两种项目的运动员M1中同一区域所对应的FDI和ECR皮质脊髓兴奋性并非全部显著优于常人的主要原因,也从侧面印证了FDI和ECR在这两项运动中的地位。
5 结论
5.1 长期专项训练促进大脑皮层兴奋性
运动员的大脑运动皮层兴奋性更加显著。长期、系统的运动训练可以增加大脑皮质脊髓束的兴奋性,提高皮质内募集水平。这种变化会随着外界刺激的增加而逐步提升。大脑皮质的兴奋易化和募集效应意味着运动皮层的重组和突触连接的加强,体现了大脑可塑性变化,从而进一步促进运动技能的学习与控制。
5.2 乒乓球和羽毛球运动员的大脑可塑性变化具有“专项相关区域化”的特点
长期专项训练在促进运动皮层兴奋的同时,还根据不同的动作和技术特征表现出了与之对应的大脑可塑性变化。不同专项运动员的可塑性变化主要产生在专项相关的主动肌群上,体现出了与专项有关的特定肌群“区域化”改变的特点。
5.3 建议与展望
在前人对大脑运动皮层经外界干预而造成的不同兴奋性变化的基础上,本研究进一步细化了长期运动以及运动特性对大脑初级运动皮层的可塑性影响,从大脑运动皮层的兴奋性与长期高强度的系统运动训练之间的双向影响分析了M1在不同运动技能掌握阶段所起的作用。与前人相比,本研究主要在以下几个方面有了新的进展和认识:1)从不同的专项出发,针对性地分析脑部特定运动区(M1)对应的不同肢体位置上(FDI和ECR)运动诱发电位兴奋性的差异,从电生理角度剖析不同专项间技术特征、参与肌群的差异与长期、系统化训练后大脑可塑性差异之间关系;2)选取专业乒乓球和羽毛球运动员,对探究高水平运动员的大脑运动皮层可塑性变化具有一定的实际意义,提出了一种可利用的新的评价运动技能水平的角度。检测大脑运动皮层神经元的激活程度以及特定激活部位或可发展成为从大脑系统层面评价训练效果和运动水平的新指标。
本研究主要立足于运动员大脑初级运动皮层的结构性变化,未来的研究可以加入不同的心理或训练任务,或展开纵向跟踪干预等,从多个方面进一步验证本研究所提出的关于运动员和新手间的大脑可塑性差异,或深入区分长期训练和高强度训练对大脑运动皮层兴奋性的影响[19]。此外,未来的研究还可以考虑将经颅磁刺激和其他电生理或神经成像技术结合运用,更加系统探究运动员、音乐家或舞蹈演员等专门群体[9,16]经过长期运动技能学习后产生的大脑可塑性改变。
[1] 吴殷,张剑,曾雨雯,等. 不同类型运动项目对运动员大脑结构可塑性变化研究[J]. 体育科学,2015,35(04):52-57.
[2] 张剑,庄洁,陈佩杰. 经颅磁刺激及其在运动科学中的应用展望[J]. 中国运动医学杂志,2009,3(28):353-356.
[3] BAECK J S,KIM Y T,SEO J H,et al. Brain activation patterns of motor imagery re fl ect plastic changes associated with intensive shooting training[J]. Behav Brain Res,2012,234:26-32.
[4] BARNES S J,FINNERTY G T. Sensory experience and cortical rewiring[J]. Neuroscientist,2010,16:186-198.
[5] BIZZI E,TRESCH M C,SALTIEL P,et al. New perspectives on spinal motor systems[J]. Nat Rev Neurosci,2000,1:101-108.
[6] CHIEN-LU TSAI. The Surface EMG Activity of The Upper Limb Muscles in Table Tennis Forehand Drives [C]. Austria:International Symposium on Biomechanics in Sports,2006.
[7] DAYAN E,COHEN L G. Neuroplasticity subserving motor skill learning[J]. Neuron,2011,72:443-454.
[8] DE BEAUMONT L,THEORET H,MONGEON D,et al. Brain function decline in healthy retired athletes who sustained their last sports concussion in early adulthood[J]. Brain,2009,132:695-708.
[9] ELBERT T,PANTEV C,WIENBRUCH C,et al. Increased cortical representation of the fi ngers of the left hand in string players[J].Sci,1995,270:305-307.
[10] ERICSSON K A,LEHMANN A C. Expert and exceptional performance:evidence of maximal adaptation to task constraints[J].Annu Rev Psychol,1996,47:273-305.
[11] GEYER S,MATELLI M,LUPPINO G,et al. Functional neuroanatomy of the primate isocortical motor system[J]. Anat Embryol (Berl),2000,202:443-474.
[12] GRAZIANO M S and AFLALO T N. Mapping behavioral repertoire onto the cortex[J]. Neuron,2007,56:239-251.
[13] HARDWICK R M,ROTTSCHY C,MIALL R C,et al. A quantitative meta-analysis and review of motor learning in the human brain[J]. Neuroimage,2013,67:283-297.
[14] HIROKI N,MICHIKO Y,AKITO M,et al. Characteristics of the athletes’ brain:Evidence from neurophysiology and neuroimaging[J]. Brain Res Rev,2010,62:197-211.
[15] IINO Y,KOJIMA T. Kinetics of the upper limb during table tennis topspin forehands in advanced and intermediate players[J].Sports Biomech,2011,10(4):361-377.
[16] KARGO W J,NITZ D A. Early skill learning is expressed through selection and tuning of cortically represented muscle synergies[J]. J Neurosci,2003,23:11255-11269.
[17] KELLY A M,GARAVAN H. Human functional neuroimaging of brain changes associated with practice[J]. Cereb Cortex,2005,15:1089-1102.
[18] KLEIM J A,HOGG T M,VANDENBERG P M,et al. Cortical synaptogenesis and motor map reorganization occur during late,but not early,phase of motor skill learning[J]. J Neurosci,2004,24:628-633.
[19] LANDI S M,BAGUEAR F and DELLA-MAGGIORE V. One week of motor adaptation induces structural changes in primary motor cortex that predict long-term memory one year later[J]. J Neurosci,2011,31:11808-11813.
[20] LEES A. Science and the major racket sports:a review[J]. J Sports Sci,2003,21:707-732.
[21] LEHERICY S,BENALI H,VAN DE MOORTELE P F,et al.Distinct basal ganglia territories are engaged in early and advanced motor sequence learning[J]. Proc Natl Acad Sci U S A,2005,102:12566-12571.
[22] MARAVITA A,IRIKI A. Tools for the body (schema)[J].Trends Cogn Sci,2004,8:79-86.
[23] NI Z,LIANG N,TAKAHASHI M,et al. Motor strategies and excitability changes of human hand motor area are dependent on different voluntary drives[J]. Eur J Neurosci,2006,23:3399-3406.
[24] O’SHEA J,JOHANSEN-BERG H,TRIEF D,et al. Functionally speci fi c reorganization in human premotor cortex[J]. Neuron,2007,54:479-490.
[25] PEARCE A J,THICKBROOM G W,BYRNES M L,et al. Functional reorganization of the corticomotor projection to the hand in skilled racquet players[J]. Exp. Brain Res,2000,130:238–243.
[26] ROBERT C,ROTHWELL J C. Cortical Connectivity[M]. Berlin:Springer-Verlag,2012:49-69.
[27] ROSENKRANZ K,KACAR A,ROTHWELL J C. Differential modulation of motor cortical plasticity and excitability in early and late phases of human motor learning[J]. J Neurosci,2007,27:12058-12066.
[28] ROSENKRANZ K,WILLIAMON A,ROTHWELL J C. Motorcortical excitability and synaptic plasticity is enhanced in professional musicians[J]. J Neurosci,2007,27:5200-5206.
[29] ROSSI S,HALLETT M,ROSSINI P M,et al. Safety,ethical considerations,and application guidelines for the use of transcranial magnetic stimulation in clinical practice and research[J].Clini neurophysiol,2009,120(12):2008-2039.
[30] SAKURAI S,OHTSUKI T. Muscle activity and accuracy of performance of the smash stroke in badminton with reference to skill and practice[J]. J Sports Sci,2000,18:901-914.
[31] SANES J N,DONOGHUE J P,THANGARAJ V,et al. Shared neural substrates controlling hand movements in human motor cortex[J]. Science,1995,268:1775-1777.
[32] SCHIEBER M H. Constraints on somatotopic organization in the primary motor cortex[J]. J Neurophysiol,2001,86:2125-2143.
[33] SCOTT S H. Optimal feedback control and the neural basis of volition motor control[J]. Nat Revs Neurosci. 2004,5(1427):534-546.
[34] TRUCCOLO W,FRIEHS G M,DONOGHUE J P,et al. Primary motor cortex tuning to intended movement kinematics in humans with tetraplegia[J]. J Neurosci,2008,28:1163-1178.
[35] WEN D,YAN-LING P,ZHEN N,et al. Maintenance of balance between motor cortical excitation and inhibition after long-term training[J]. Neurosci,2016,336:114-122.
[36] YOSHIDA K,SUGIYAMA S. Di ff erences between EMGs of forearm skeletal muscles for fl ick strokes against backspin and no-spin services in table tennis[J]. Int J Table Tennis Scis,2010.
Comparison of Brain Plasticity between Table Tennis and Badminton Athletes
DAI Wen,ZHANG Jian,TAN Xiao-ying
Objective:Long-term and systematic exercise training can lead to plastic changes in motor cortex. Research and analysis on characteristics of di ff erent sports in terms of brain level not only helps athletes and coaches to understand the characteristics of sports more accurately,but also provides objective and physiological indicators for selection and training. Methods:This study chooses fi rst dorsal interosseous and extensor carpi radialis which mainly exercised in table tennis and badminton games and produces motor evoked potential (MEP) through single pulses of transcranial magnetic stimulation in the primary motor cortex of the brain to evaluate motorcortical excitability.Results:The MEP amplitude and slope of fi rst dorsal interosseous in table tennis athletes were significantly higher than those in the control group. The MEP amplitude and slope of extensor carpi radialis in badminton athletes were signi fi cantly higher than those in the table tennis group and the control group. Conclusion:Long-term and systematic exercise training can increase motorcortical excitability of agonist-related region “selectively” and improve recruitment of corticospinal projections to help athletes better achieve motor output.
motor skill;transcranial magnetic stimulation;brain plasticity;motorcortical excitability;motor evoked potential
G804.8
A
2017-07-08;
2017-09-25
国家自然科学基金资助项目(31371056);上海市人类运动能力开发与保障重点实验室项目(11DZ2261100)。
戴雯,女,硕士,主要研究方向为运动技能学习与控制,E-mail:348258655@qq.com。
谭晓缨,男,副教授,博士,主要研究方向为体育教育与训练、体育社会学,E-mail:178979334@qq.com。
上海体育学院,上海 200438 Shanghai University of Sport,Shanghai 200438,China.
1002-9826(2017)06-0127-06
10. 16470/j. csst. 201706016
猜你喜欢
幼儿智力世界(2022年3期)2022-03-12 15:52:26
昆明医科大学学报(2022年1期)2022-02-28 07:43:34
昆明医科大学学报(2021年8期)2021-08-13 08:59:20
昆明医科大学学报(2021年6期)2021-07-31 07:40:08
中国现代医药杂志(2020年3期)2020-05-08 04:33:08
中国生物医学工程学报(2019年6期)2019-07-16 07:52:48
幼儿100(2018年35期)2018-12-29 12:32:22
小天使·二年级语数英综合(2018年10期)2018-10-15 09:20:10
中外医疗(2016年15期)2016-12-01 04:25:39
小主人报(2016年2期)2016-02-28 20:46:35
