
南麂列岛海域蟹类群落结构及其与环境因子的关系
2017-10-09 01:01:49谢旭俞存根蔡厚才郑基陈万东伍尔魏夏陆军毕耜瑶
海洋学报 2017年10期
谢旭,俞存根*,蔡厚才,郑基,陈万东,伍尔魏,夏陆军,毕耜瑶
(1.浙江海洋大学 水产学院,浙江 舟山 316022;2.南麂列岛国家海洋自然保护区管理局,浙江 平阳 325401)
南麂列岛海域蟹类群落结构及其与环境因子的关系
谢旭1,俞存根1*,蔡厚才2,郑基1,陈万东2,伍尔魏2,夏陆军1,毕耜瑶1
(1.浙江海洋大学 水产学院,浙江 舟山 316022;2.南麂列岛国家海洋自然保护区管理局,浙江 平阳 325401)
根据2013年11月、2014年2月、5月、9月在南麂列岛国家海洋自然保护区海域进行生物资源调查所获得的资料,分析了南麂列岛海域蟹类种类组成、优势种、生物多样性等群落结构特征,并定量分析了群落结构与水文环境因子之间的关系。结果表明,周年4个季度月调查共鉴定出蟹类21种,隶属7科、12属,其中优势种为三疣梭子蟹(Portunustrituberculatus)、日本蟳(Charybdisbimaculata)和双斑蟳(Charybdisbimaculata)3种。不同季节的蟹类种类组成差异较大,而优势种类组成变化较少,其中,以秋季的蟹类种类数最多,冬季最少。从不同水深区域的渔获种类分布趋势来看,蟹类种类以20~30 m水深带较多,30~40 m水深带较少。夏季多样性指数低于其他季节,以水深来看,多样性指数在10~20 m水深带较高, 30~40 m水深带较低,蟹类多样性指数与水深呈反比。根据冗余分析认为, 水深、水温和盐度是影响调查海域蟹类种类组成和群落结构特征的主要环境因子。
蟹类;多样性;群落结构;环境因子;南麂列岛海域
1 引言
自20世纪80年代以来,我国近海主要经济鱼类资源不断衰退,导致作业结构不断调整,特别是20世纪90年代以后,随着蟹笼作业的发展,东海虾蟹类资源不断遭到开发,渔获量呈逐年递增趋势,2014年浙江省蟹类海洋捕捞产量达到了29.66×104t[1],利用品种也在逐渐增多,目前蟹类海洋捕捞作业在东海海洋捕捞业中占有越来越重要的地位[2-3]。南麂列岛国家海洋自然保护区位于我国东海中南部的浙南海域的温台渔场,该海域岛屿众多,水文环境主要受台湾暖流与江浙沿岸流的交互影响,特殊的地形及复杂的海流系统,形成了上升流,造就了该海域水质肥沃、营养物质丰富、饵料生物充足、水文环境适宜,是多种经济鱼类、虾类、蟹类、贝类、藻类等生物繁殖、索饵、生长的优良栖息地,生物种类繁多,生物多样性丰富,素有“贝藻王国”之称。因此,分析南麂列岛海域蟹类的群落结构特征与环境因子的关系,掌握其分布变化规律,对南麂列岛国家海洋自然保护区的保护,以及保护区的生态修复具有重要意义。
我国首批国家级海洋自然保护区中唯一一个位于东海区的南麂列岛国家海洋自然保护区,吸引了许多学者对其进行研究。过去,关于南麂列岛海域生物的研究,多集中在浮游动物、底栖生物以及贝藻类[4-11],而对南麂列岛海域的蟹类生物资源的研究并不多见,主要有仇林根[12]报道了南麂列岛蟹类种类组成和区系特点,晁文春等[13-14]研究报道了南麂列岛附近海域甲壳类群落结构特征。但是,对于浅海区的渔业资源系统定点专业调查研究,至今未见有报道。本文主要根据2013-2014年在南麂列岛海域开展渔业资源拖网调查所获得的蟹类数据,分析研究南麂列岛海域的蟹类的群落结构特征与水文环境之间的关系,目的是掌握南麂列岛海域蟹类资源现状,以期为南麂列岛国家海洋自然保护区蟹类资源的科学保护、管理及其可持续利用提供依据。
2 材料与方法
本文所采用数据采自2013年11月(秋季)、2014年2月(冬季)、2014年5月(春季)、2014年9月(夏季)在南麂列岛国家海洋自然保护区40 m水深以内浅海区开展渔业资源定点拖网调查所获得的资料。调查范围为27°24′30″~27°30′30″N,120°56′30″~121°08′30″E,共设置20个调查站位(图1)。调查船租用“浙苍渔0942号”底拖网渔船,主机功率为202 kW,网具为拖网,规格为750目×80 mm,作业时网口宽度为10.8 m。船上配备有卫导、探鱼仪、对讲机等导航、定位等助渔助航设备。调查时每站拖曳约1 h,拖速约为3 kn。渔业资源调查方法按照《海洋渔业资源调查规范》(SC/9403-2012)[15]进行。每个调查站位拖网所获的渔获物全部取样装入样品袋,并进行编号、记录后,冰鲜保存,带回实验室分析、鉴定,使用电子天平称重,精确度为0.1 g。同时,与底拖网调查同步,利用CTD测定各调查站位的水温、盐度数据。

图1 调查站位图Fig.1 The survey stations of fishery resources
优势种的计算采用相对重要性指数,计算公式如下:
IRI=[(ni/N+wi/W)×fi/m]×105,
(1)
式中,ni、wi分别为第i种蟹类的个体数和生物量;N、W分别为蟹类总个体数和总生物量;fi为第i种蟹在m次取样中出现的频率;m为取样次数。
优势种的季节更替变化运用种类更替率[16]分析,计算公式如下:

(2)
式中,A为更替率,即与前一季节比较的更替情况;C为相临两个季节间减少及增加的物种数;S为相临两个季节间相同的物种数。
物种多样性主要采用Shannon-Weaver(H′)指数、均匀度(J′)、丰富度(D)3个公式计算,计算公式如下:
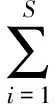
(3)
J′=H′/log2S,
(4)
D=(S-1)/log2N,
(5)
式中,H′为蟹类多样性指数;J′为蟹类均匀度指数;D为蟹类丰富度指数;ni是第i种蟹的个体数,N为蟹类总尾数;S为蟹类总种数。
群落季节更替指数AI和迁移指数MI[17],计算公式为:
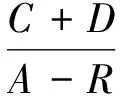
(6)
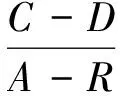
(7)
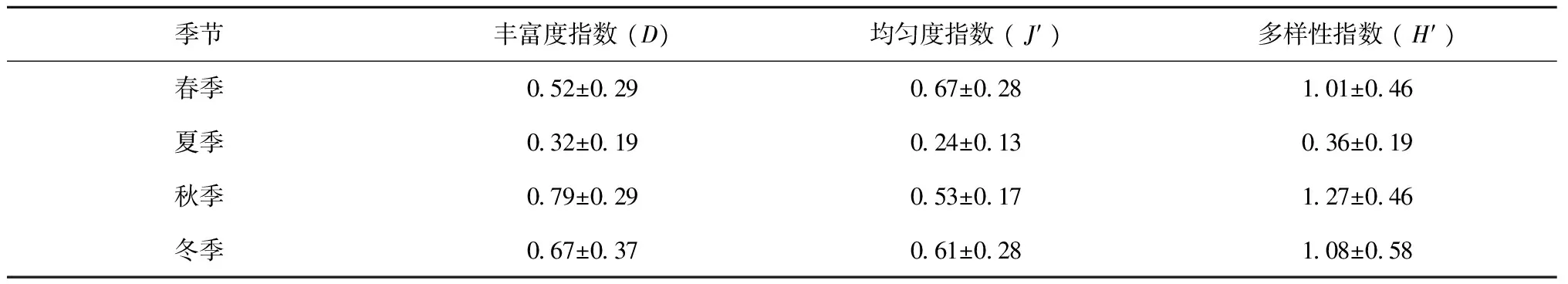
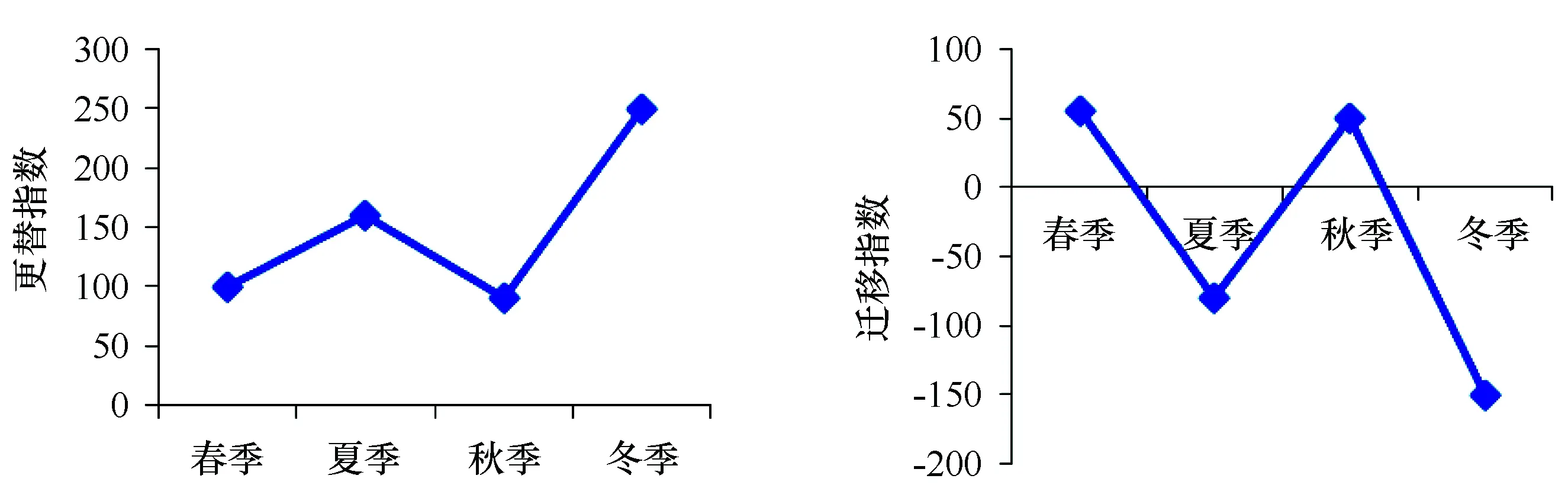
式中,A为各季节间实际物种数,C为本季节新迁入物种数,D为本季节将要迁出物种数,R为全年均出现的物种数。AI反映物种更替导致群落稳定性降低的节律,其值越大,表示群落稳定性越差;MI表示研究系统外迁入与迁出种的两种成分在群落中的相对比例,当C>D时,MI为正值,表示迁入种多于迁出种,当C=D时,MI则为零,表示群落动态平衡,当C 蟹类数量分布图采用Surfer8软件绘制。生物量和水文环境之间的关系采用冗余分析(Redundancy analysis, RDA)方法分析[18-19],在CANOCO3.0软件进行运算, 排序结果用物种-环境因子关系的双序图表示。 3.1 种类组成 根据周年4个季度月调查采集样品,共鉴定出蟹类21种,隶属7科、12属;秋季出现种类最多,为13种;春季次之,为12种;夏季和冬季较少,分别为8种和7种(表1)。各季均以梭子蟹科出现种类数最多,周年出现的种类,均隶属于梭子蟹科,可见梭子蟹科是南麂列岛海域蟹类的主要类群。四季共有种为三疣梭子蟹、日本蟳和矛形梭子蟹3种,其生物量占蟹类总生物量的98.34%;仅在一个季节出现的蟹类12种,其生物量占蟹类总生物量的0.11%。 表1 南麂列岛海域蟹类的科、属和种的组成 注:本文的生物种类名录主要参照《中国海洋生物名录》[20]。 将调查海域按水深分成水深10~20 m、20~30 m、30~40 m 3个水深带(表2),来分析南麂列岛海域的蟹类种类组成。结果如表3所示,春、夏、秋3个季节均是水深在20~30 m之间区域的蟹类种类最多,水深在10~20 m之间区域的蟹类种类次之,水深在30~40 m之间区域的蟹类种类最少;冬季蟹类种类随水深增加呈递减趋势,水深在10~20 m之间区域的蟹类最多,水深在20~30 m之间区域的蟹类种类次之,水深在30~40 m之间区域的蟹类种类最少;水深在10~20 m之间区域的蟹类秋季最多有11种,春、夏季节较少均有6种,相差5种;水深在20~30 m之间区域蟹类秋季最多有12种,冬季最少有6种,相差6种;水深在30~40 m之间区域的蟹类秋季最多有7种,春、季最少有3种,相差4种;3个水深带均有分布的共有种较少,秋季共有种最多,有7种,夏冬季共有种次之,春季共有种最少,仅有2种。说明栖息在水深20~30 m之间区域的蟹类种类最多,栖息水深在30~40 m之间区域的蟹类种类最少。 表2 南麂列岛海域不同水深带站位点分布 表3 南麂列岛海域不同水深带蟹类种类组成 3.2 优势种 根据相对重要性指数(IRI)分析研究蟹类群落中的优势种成分,不同种类的相对重要性指数如表4所示。将相对重要性指数(IRI)大于1 000的定为优势种,100~1 000的定为常见种。结果得知,周年4个季度月调查优势种为三疣梭子蟹、日本蟳和双斑蟳等3种,不同季节的优势种存在明显的季节更替,春-夏季、夏-秋季、秋-冬季之间优势种更替率分别为66.67%、50.00%和33.33%,春季优势种有三疣梭子蟹、日本蟳和双斑蟳3种,夏季优势种有三疣梭子蟹1种,秋季优势种有三疣梭子蟹和日本蟳2种,冬季优势种有三疣梭子蟹、日本蟳和双斑蟳3种。 表4 南麂列岛海域主要蟹类相对重要性指数值(IRI) 3.3 物种多样性 比较不同季节蟹类的多样性,由表5可见,蟹类各季节的丰富度指数(D)以秋季最高,春冬季其次,夏季最低;均匀度指数(J′)以春季最高,秋冬季其次,夏季最低;多样性指数(H′)以秋季最高,春冬季其次,夏季最低。多样性指数(H′)分布如图2所示,春季呈东西两端向岛礁区递减趋势,夏冬季呈南北两端向岛礁区递减趋势,秋季呈自北向南递减趋势。 表5 南麂列岛海域蟹类群落多样性季节变化 图2 南麂列岛海域蟹类群落多样性指数(H′)分布Fig.2 Distribution of crab diversity(H′)in the Nanji Islands ground shallow sea area waters 水深/m多样性春季夏季秋季冬季平均10~20D06704910208407520~3004802307006305130~4004205207205005410~20J′07402305506605420~3006702405206105130~4005002905104504410~20H′13404614212911320~3009702912310308830~40050057113090078 比较不同水深蟹类的多样性,由表6可以看出,蟹类的丰富度指数(D)四季平均值随水深增加先减小后增大,春冬季蟹类的丰富度指数(D)随水深加深而减小,夏秋季蟹类的丰富度指数(D)随水深加深先递减后递增。蟹类的均匀度指数(J′)四季平均值随水深增加而减小,夏季蟹类的均匀度指数(J′)随水深增加而增大,春秋冬季蟹类的均匀度指数(J′)随水深增加而减小。蟹类的多样性指数(H′)四季平均值随水深增加而减小,夏季蟹类的多样性指数(H′)随水深增加先减小后增大,春秋冬季蟹类的多样性指数(H′)随水深增加而减小。 3.4 聚类、排序结果分析 南麂列岛海域蟹类生物量进行聚类分析和NMDS排序的结果如图3所示,4季NMDS分析结果的胁强系数在0.06~0.19之间,表明该图形较好的反映了群落间的相似性程度[21]。各季节站位聚类分析和NMDS分析结果基本一致,春季该海域的蟹类可划分为3个群落,分别为群落Ⅰ、群落Ⅱ和群落Ⅲ;夏季该海域的蟹类可划分为3个群落,分别为群落Ⅰ、群落Ⅱ和群落Ⅲ;秋季该海域的蟹类可划分为2个群落,分别为群落Ⅰ和群落Ⅱ;冬季该海域的蟹类可划分为2个群落,分别为群落Ⅰ和群落Ⅱ。 图3 南麂列岛海域蟹类采样站点的聚类分析和NMDS排序图Fig.3 Group average clustering and NMDS ordination of sampling stations in the Nanji Islands ground shallow sea area waters 3.5 群落相似性分析 如表7所示,群落Ⅰ中的主要种类有三疣梭子蟹、日本蟳、双斑蟳、矛形梭子蟹(Portunushastatoides)和锈斑蟳(Charybdisferiatus),三疣梭子蟹和日本蟳群落内相似性较高;群落Ⅱ中的主要种类有三疣梭子蟹、双斑蟳、日本蟳、矛形梭子蟹、锈斑蟳、红星梭子蟹、隆线强蟹(Eucratecrenata)、变态蟳和豆形短眼蟹(Xenophthalmuspinnotheriodes),三疣梭子蟹、双斑蟳和日本蟳群落内相似性较高;群落Ⅲ中的主要种类有三疣梭子蟹、双斑蟳、红星梭子蟹、日本蟳和锈斑蟳,三疣梭子蟹、双斑蟳和日本蟳群落内相似性较高。 如表8所示,春季,群落Ⅰ和群落Ⅱ的相异性为94.23%,主要分歧种为三疣梭子蟹、双斑蟳、日本蟳和红星梭子蟹等,群落Ⅰ和群落Ⅲ的相异性为94.28%,主要分歧种为三疣梭子蟹、日本蟳、红星梭子蟹和绒毛细足蟹等,群落Ⅱ和群落Ⅲ的相异性为94.28%,主要分歧种为三疣梭子蟹、双斑蟳和日本蟳等;夏季,群落Ⅰ和群落Ⅱ的相异性为85.43%,主要分歧种为三疣梭子蟹等,群落Ⅰ和群落Ⅲ的相异性为58.42%,主要分歧种为三疣梭子蟹等,群落Ⅱ和群落Ⅲ的相异性为57.14%,主要分歧种为三疣梭子蟹和红星梭子蟹等;秋季,群落Ⅰ和群落Ⅱ的相异性为65.33%,主要分歧种为三疣梭子蟹和双斑蟳等;冬季,群落Ⅰ和群落Ⅱ的相异性为68.18%,主要分歧种为三疣梭子蟹、双斑蟳和日本蟳等。 3.6 群落结构稳定性 根据更替、迁移指数研究调查海域4个季节群落稳定性变化趋势,由图4可知,调查海域各季节群落更替指数均在90以上,说明调查海域蟹类群落稳定性较差,其中冬季蟹类群落稳定性最差,秋季蟹类群落稳定性最好。由图4可知,调查海域蟹类春季和秋季迁入量大于迁出量,夏季和冬季迁入量小于迁出量。 图4 南麂列岛海域蟹类群落更替指数和迁移指数Fig.4 The AI and MI of crab community in the Nanji islands ground shallow sea area waters 3.7 蟹类群落与水文环境因子的关系 RDA是一种线性多元直接梯度分析方法[22],它把对应分析与多元回归结合起来, 每一步计算结果都与环境因子进行回归, 以确定蟹类群落与环境的对应关系[19]。在RDA排序图中, 环境因子用带有箭头的线段表示, 连线的长短表示蟹类群落分布和种类分布与该环境因子关系的相关性大小,连线越长,说明相关性越大,反之越小。箭头连线与排序轴的夹角表示该环境因子与排序轴相关性大小,夹角越小,相关性越高,反之越低。箭头连线与某一站位(或种类)的垂直线距离表示其相关性大小,垂线距离越小,正相关性越高,反之越低, 处于另一端的则表示与该类环境因子具有负的相关性。调查海域蟹类群落与表层温度(ST)、表层盐度(SS)、底层温度(BT)、底层盐度(BS)和水深(D)等5个环境因子的RDA排序结果如图5和表9所示。 春季调查海域蟹类群落受底层盐度影响较大,底层盐度与第一、第二排序轴的相关性分别为0.964和0.267,三疣梭子蟹受其影响较大。夏季调查海域蟹类群落受底层温度影响较大,底层温度与第一、第二排序轴的相关性分别为0.992和-0.126,三疣梭子蟹和变态蟳受其影响较大。秋季调查海域蟹类群落受水深、底层温度和底层盐度影响较大,水深与第一、第二排序轴的相关性分别为0.604和0.797,双斑蟳受其影响较大;底层温度与第一、第二排序轴的相关性分别为0.105和0.994,红星梭子蟹、矛形梭子蟹和双斑蟳受其影响较大;底层盐度与第一、第二排序轴的相关性分别为0.097和0.995,红星梭子蟹、矛形梭子蟹和双斑蟳受其影响较大。冬季调查海域蟹类群落受表层温度和底层温度影响较大,表层温度与第一、第二排序轴的相关性分别为0.073和-0.997,豆形短眼蟹和兰氏三强蟹(Tritodynamiarathbunae)受其影响较大;底层温度与第一、第二排序轴的相关性分别为0.621和0.784,矛形梭子蟹受其影响较大。 图5 南麂列岛海域蟹类站位与环境因子的各季节RDA排序Fig.5 The seasonal distribution RDA sort of crab stance and environmental factors in the Nanji Islands ground shallow sea area waters 表9 南麂列岛海域蟹类群落结构和水文环境因子之间的相关性分析 注:*表示0.01 4.1 蟹类群落种类组成 周年4个季度月调查共鉴定出蟹类21种,季节种类由低到高依次为:秋季(13种)、春季(12种)、夏季(8种)、冬季(7种),4个季节均有分布的种类仅有三疣梭子蟹、日本蟳和矛形梭子蟹3种,约占蟹类总种数的14.29%,大部分种类只在某些季节出现。表明南麂列岛海域蟹类种类以季节性种类为主。与相关研究结果[13-14]相比,周年4个季度月调查捕获蟹类种类较多,梭子蟹科种类数最多。从生态类型看,21种蟹大多为广温、广盐性种类,这可能与台湾暖流和浙江沿岸流交互影响[10]有关。从不同水深区域分析,蟹类种类数随水深增加呈先递增后递减趋势,以水深在20~30 m之间的区域蟹类种数最多,水深在30~40 m之间的区域蟹类种数最少。说明秋季许多蟹类洄游到此索饵、生殖,并且主要栖息于水深在20~30 m之间区域。 4.2 优势种 与仇林根[12]调查结果相比较,调查海域的蟹类的优势种发生了较大的变化,优势种类组成已从90年代初的锯缘青蟹(Scyllaserrata)、红星梭子蟹、三疣梭子蟹、矛形梭子蟹、日本蟳、锈斑蟳、双斑蟳、隆线强蟹等31种演变为周年4个季度月调查的三疣梭子蟹、日本蟳和双斑蟳3种。南麂列岛海域蟹类优势种春季和夏季更替比较明显,更替率分别为66.67%,这可能是由于春季优势种三疣梭子蟹在4-7月处于繁殖期[23],产下大量幼体,春夏季水温环境适宜,饵料生物丰富,幼体生长迅速,使其占据绝对优势,所以优势种由春季的三疣梭子蟹、日本蟳和双斑蟳3种演变为夏季的三疣梭子蟹1种。秋冬季随着水温降低,大量三疣梭子蟹开始向外部较深海域进行洄游,调查海域三疣梭子蟹生物量减少,日本蟳和双斑蟳逐渐成为优势种。 4.3 物种生物多样性 从不同季节蟹类的多样性来看,夏季调查海域蟹类丰富度指数(D)、均匀度指数(J′)和多样性指数(H′)均低于其他季节。这可能是由于调查海域受台湾暖流和浙江沿岸流交互影响导致的, 夏季调查海域主要受台湾暖流影响[24], 其温度、盐度相对较高,导致夏季广温广盐性种类减少,从表4也可以看出,夏季优势种仅有三疣梭子蟹1种,而其他季节均有2种以上。根据何贤保等[25]研究南麂列岛海域夏季鱼类数量最多,而夏季许多蟹类处于幼体阶段,被鱼类大量捕食[26],也可能是夏季蟹类生物多样性最低的原因之一。从不同水深区域分析南麂列岛海域的蟹类的多样性可以看出,春、秋、冬3季蟹类丰富度指数(D)、均匀度指数(J′)和多样性指数(H′)随水深增加呈递减趋势,夏季蟹类丰富度指数(D)、均匀度指数(J′)和多样性指数(H′)随水深增加呈先递减后递增趋势。 4.4 群落结构特征 从图3可以看出,南麂列岛海域蟹类群落结构变化明显。根据CLUSTER分析,春季和夏季可划分为3个不同的群落,秋季和冬季可划分为2个不同的群落,NMDS二维标序图较好地反映了4个季节群落间的相似性程度。夏季和秋季群落相似性较高,春季群落相似性较低,对相似性影响较大的特征种主要有日本蟳、三疣梭子蟹和双斑蟳。春季群落间相异性较大,其他季节次之,对相异性影响较大的分歧种主要有三疣梭子蟹、双斑蟳、日本蟳、红星梭子蟹和绒毛细足蟹。冬季随着台湾暖流的减弱,浙江沿岸流影响增强[24],水温的降低,大量蟹类向外部较深海域进行洄游,迁入量小于迁出量,冬季更替率大于其他季节,群落稳定性最差。 4.5 群落结构特征与环境因子的关系 根据蟹类群落与水文环境因子相关性进行分析,结果表明:周年4个季度月调查所获21种蟹类中,4个季节均出现的种类仅有3种,85.71%的蟹类是洄游性蟹类,而蟹类的洄游能够直接影响蟹类的分布情况,通过RDA分析,可能是因为调查海域水深相对较浅,离大陆较近,且处在台湾暖流和浙江沿岸流的交汇区,受其交互影响,以及降水、大陆径流等,导致底层温度是影响蟹类分布最为重要的环境因子,夏秋冬3个季节底层温度变化范围较大,分别为2.43℃、1.44℃和0.85℃,蟹类分布受底层温度影响显著,春季底层温度变化范围较小,为0.60℃,春季蟹类分布受其影响很弱;其次是底层盐度对蟹类春秋2个季节的分布影响较明显;而表层温度仅在冬季对蟹类的分布影响较明显,这可能是因为蟹类主要栖息在底层,采用底拖网作业的采样方式有关;水深仅在秋季对蟹类的分布影响较明显,其他季节其影响很弱。 周年4个季度调查所获21种蟹类,主要是以广温、广盐性种类为主,4个季节均出现的种类仅有3种,85.71%的蟹类是洄游性蟹类;从不同水深区域分析,分布在水深20~30 m之间的区域蟹类最多,分布在水深30~40 m区域的蟹类种数最少。优势种为三疣梭子蟹、日本蟳和双斑蟳等3种。多样性指数以秋季最高,春冬季其次,夏季最低,随水深增加多样性指数呈先递减后递增趋势,在水深20~30 m之间区域多样性最低,春秋冬3个季节多样性指数均随水深增加而递减。对相似性影响较大的特征种主要有日本蟳、三疣梭子蟹和双斑蟳等,对相异性影响较大的分歧种主要有三疣梭子蟹、双斑蟳、日本蟳、红星梭子蟹和绒毛细足蟹等;春秋季蟹类物种迁入量大于迁出量,群落稳定性较好,夏冬季蟹类物种迁入量小于迁出量,群落稳定性较差。通过RDA分析,底层温度和底层盐度是影响调查海域蟹类分布的主要环境因子。 致谢:倪孝品、曾贵侯等同志参加海上样品采集和室内种类鉴定,赵盛龙老师、章飞军老师、王迎宾老师、叶深、李德伟、李新、郭小雨等同志参加室内种类鉴定和数据处理, 谨致谢忱。 [1] 浙江省海洋与渔业局. 2014年浙江省渔业经济统计资料[Z]. 2015. Marine and Fisheries Bureau of Zhejiang Province. 2014 Zhejiang fishery economic statistics[Z]. 2015. [2] 俞存根, 宋海棠, 姚光展. 东海大陆架海域蟹类资源量的评估[J]. 水产学报, 2004, 28(1): 41-46. Yu Cungen, Song Haitang, Yao Guangzhan. Assessment of the crab stock biomass in the continental shelf waters of the East China Sea[J]. Journal of Fisheries of China, 2004, 28(1): 41-46. [3] 俞存根, 宋海棠, 姚光展, 等. 东海大陆架海域经济蟹类种类组成和数量分布[J]. 海洋与湖沼, 2006, 37(1): 53-60. Yu Cungen, Song Haitang, Yao Guangzhan, et al. Composition and distribution of economic crab species in the East China sea[J]. Oceanologia et Limnologia Sinica, 2006, 37(1): 53-60. [4] 尤仲杰, 孙建章, 王一农. 南麂列岛的贝类[C]//浙江省海洋管理局. 南麂列岛国家级海洋自然保护区论文选(一). 北京: 海洋出版社, 1992: 34-54. You Zhongjie, Sun Jianzhang, Wang Yinong. Shellfish in the Nanji islands[C]//Oceanic Administration of Zhejiang Province. Selected Papers in Nanji Islands National Marine Conservation Area. Beijing: China Ocean Press, 1992: 34-54. [5] 陈赛英, 王一婷, 孙建章, 等. 浙江南麂列岛贝类区系的研究[C]//南麂列岛国家级海洋自然保护区论文选. 北京: 海洋出版社, 1992: 55-60. Chen Saiying, Wang Yiting, Sun Jianzhang, et al. The research area of shellfish in the Nanji Islands[C]//Selected Papers in Nanji Islands National Marine Conservation Area. Beijing: China Ocean Press, 1992: 55-60. [6] 高爱根, 董永庭, 王慧珍, 等. 南麂列岛邻近海域贝类生态分布的初步研究[J]. 东海海洋, 1998, 16(2): 49-54. Gao Aigen, Dong Yongting, Wang Huizhen, et al. Preliminary study on the distribution of mollusca ecology in Sublittoral Area of Nanji Islands[J]. Donghai Marine Science, 1998, 16(2): 49-54. [7] 蔡如星, 江福连, 车建国, 等. 浙江南麂水域的蔓足类[C]//浙江省海洋管理局. 南麂列岛国家级海洋自然保护区论文选(一). 北京: 海洋出版社, 1992: 69-76. Cai Ruxing, Jiang Fulian, Che Jianguo, et al. Cirripedia in the Nanji islands, Zhejiang Province[C]//Oceanic Administration of Zhejiang Province. Selected Papers in Nanji Islands National Marine Conservation Area. Beijing: China Ocean Press, 1992: 69-76. [8] 彭欣, 谢起浪, 陈少波, 等. 南麂列岛潮间带底栖生物时空分布及其对人类活动的响应[J]. 海洋与湖沼, 2009, 40(5): 584-589. Peng Xin, Xie Qilang, Chen Shaobo, et al. Distribution of intertidal benthos and the human impact in Nanji Islands, China[J]. Oceanologia et Limnologia Sinica, 2009, 40(5): 584-589. [9] 张晓辉, 周燕, 龙华, 等. 南麂列岛海洋保护区浮游动物调查[J]. 动物学杂志, 2006, 41(4): 83-86. Zhang Xiaohui, Zhou Yan, Long Hua, et al. Investigation on zooplankton in Nanji marine protected area (MPA)[J]. Chinese Journal of Zoology, 2006, 41(4): 83-86. [10] 纪焕红, 叶属峰, 刘星, 等. 南麂列岛海洋自然保护区浮游动物的物种组成及其多样性[J]. 生物多样性, 2006, 14(3): 206-215. Ji Huanhong, Ye Shufeng, Liu Xing, et al. The species composition and diversity of zooplankton in Nanji Islands national nature reserve[J]. Biodiversity Science, 2006, 14(3): 206-215. [11] 纪焕红, 叶属峰, 刘星, 等. 南麂列岛海洋自然保护区浮游动物丰度和生物量的时空分布[J]. 海洋通报, 2007, 26(1): 55-60, 88. Ji Huanhong, Ye Shufeng, Liu Xing, et al. Temporal and spatial distribution of abundance and biomass of zooplankton in the Nanji Islands marine nature reserve[J]. Marine Science Bulletin, 2007, 26(1): 55-60, 88. [12] 仇林根. 南麂海区的海洋鱼类及主要甲壳类[C]//浙江省海洋管理局. 南麂列岛国家级海洋自然保护区论文选(一). 北京: 海洋出版社, 1992: 77-87. Qiu Lingen. Marine fish and curstaceans in the Nanji Islands[C]//Oceanic Administration of Zhejiang Province. Selected Papers in Nanji Islands National Marine Conservation Area. Beijing: China Ocean Press, 1992: 77-87. [13] 晁文春. 南麂列岛附近海域甲壳类群落结构与生物多样性分析[D]. 舟山: 浙江海洋学院, 2013. Chao Wenchun. Community structure and species diversity of crustaceans in the waters of Nanji Islands[D]. Zhoushan: Zhejiang Ocean University, 2013. [14] 晁文春, 何贤保, 苗振清, 等. 春夏季南麂列岛海域甲壳类种类组成及分布特征[J]. 浙江海洋学院学报: 自然科学版, 2013, 32(3): 214-221. Chao Wenchun, He Xianbao, Miao Zhenqing, et al. The species composition and diversity of crustaceans in the waters of Nanji Islands[J]. Journal of Zhejiang Ocean University:Natural Science, 2013, 32(3): 214-221. [15] 中华人民共和国国家质量监督检验检疫总局, 中国国家标准化管理委员会. GB/T 12763.6-2007, 海洋调查规范 第6部分: 海洋生物调查[S]. 北京: 中国标准出版社, 2008: 1-157. General Administration of Quality Supervision, Inspection and Quarantine of the People’s Republic of China, GB/T 12763.6-2007, Standardization Administration of the People’s Republic of China. Specifications for oceanographic survey—Part 6: marine biological survey[S]. Beijing: Standards Press of China, 2008: 1-157. [16] 刘瑞玉. 胶州湾生态学和生物资源[M]. 北京: 科学出版社, 1992: 1-460. Liu Ruiyu. Jiaozhou Bay Ecological and Biological Resources[M]. Beijing: Science Press, 1992: 1-460. [17] 朱鑫华, 吴鹤洲, 徐凤山, 等. 黄渤海沿岸水域游泳动物群落多样性及其相关因素的研究[J]. 海洋学报, 1994, 16(3): 102-112. Zhu Xinhua, Wu Hezhou, Xu Fengshan, et al. Swimming Huang Bohai coastal waters research community, diversity and its related factors[J]. Haiyang Xuebao, 1994, 16(3): 102-112. [18] ter Braak C J F. Canonical correspondence analysis: a new eigenvector technique for multivariate direct gradient analysis[J]. Ecology, 1986, 67(5): 1167-1179. [19] ter Braak C J F, Prentice I C. A theory of gradient analysis[J]. Advances in Ecological Research, 1988, 18: 271-317. [20] 刘瑞玉. 中国海洋生物名录[M]. 北京: 科学出版社, 2008: 1-1267. Liu Ruiyu. Checklist of Marine Biota China Seas[M]. Beijing: Science Press, 2008: 1-1267. [21] Quetglas A, Carbonell A, Snchez P. Demersal continental shelf and upper slope cephalopod assemblages from the Balearic Sea (North-Western Mediterranean). Biological aspects of some deep-sea species[J]. Estuarine, Coastal and Shelf Science, 2000, 50(6): 739-749. [22] 刘淑德, 线薇微, 刘栋. 春季长江口及其邻近海域鱼类浮游生物群落特征[J]. 应用生态学报, 2008, 19(10): 2284-2292. Liu Shude, Xian Weiwei, Liu Dong. Characteristics of ichthyoplankton assemblages in Yangtze Estuary and adjacent waters in spring[J]. Chinese Journal of Applied Ecology, 2008, 19(10): 2284-2292. [23] 宋海棠, 俞存根, 薛利建, 等. 东海经济虾蟹类[M]. 北京: 海洋出版社, 2006: 83-85. Song Haitang, Yu Cungen, Xue Lijian, et al. The Economical Crabs and Shrimps in East China Sea[M]. Beijing: China Ocean Press, 2006: 83-85. [24] 许建平, 杨士英. 南麂列岛及其附近海域的水文和气候特征[C]//浙江省海洋管理局. 南麂列岛国家级海洋自然保护区论文选(一). 北京: 海洋出版社, 1992: 1-9. Xu Jianping, Yang Shiying. Hydrological and climatic features in the Nanji islands[C]//Oceanic Administration of Zhejiang Province Selected Papers in Nanji Islands National Marine Conservation Area. Beijing: China Ocean Press, 1992: 1-9. [25] 何贤保, 章飞军, 林利, 等. 南麂列岛岛礁区域鱼类种类组成和数量分布[J]. 海洋与湖沼, 2013, 44(2): 453-460. He Xianbao, Zhang Feijun, Lin Li, et al. Species composition and quantitative distribution of fishes in island-reef water of Nanji Islands[J]. Oceanologia et Limnologia Sinica, 2013, 44(2): 453-460. [26] 程济生, 郭学武. 渤海底栖生物的种类、数量分布及其动态变化[J]. 海洋水产研究, 1998, 19(1): 31-42. Cheng Jisheng, Gao Xuewu. Distribution and dynamic variations of species and quantity of benthos in the Bohai Sea[J]. Marine Fisheries Research, 1998, 19(1): 31-42. The crab community structure and its relationship with environment factors in the Nanji Islands ground shallow sea area waters Xie Xu1, Yu Cungen1, Cai Houcai2, Zheng Ji1, Chen Wandong2, Wu Erwei2, Xia Lujun1,Bi Siyao1 (1.MarineFisheryCollege,ZhejiangOceanUniversity,Zhoushan316022,China; 2.NanjiIslandsNationalMarineNatureReserveAdministration,Pingyang325401,China) Based on the data obtained via the marine biological resources surveys of the Nanji Islands ground shallow sea area waters in November, 2013 and in February, May and September,2014, characteristics of crabs community including species composition, dominant species and biodiversity, were analyzed, and the related environmental factors were also examined. A total of 21 species, belonging to 12 genera in 7 families, were identified. The dominant species arePortunustrituberculatus,CharybdisbimaculataandCharybdisbimaculata. The species composition rather than dominant species showed clear seasonal changes. The maximum and minimum number of crab species were found in autumn and winter, respectively. The distribution of caught fisheries resources along water depths was documented that the morespecies were found at depths of 20-30 m and less species were found at depth of 30-40 m. The Shannon-Weaver diversity index of community in summer were lower than other seasons, and along water depths decreased from 10-20 m to 30-40 m, which means the Shannon-Weaver diversity index of crab community was in inverse proportion of the depth. Further redundancy analysis showed that water depth, temperature and salinity were the main environmental factors influencing crab species composition and community structure in this coastal conservation area. crab; diversity; community structure; environmental factors; Nanji Islands ground shallow sea area waters 10.3969/j.issn.0253-4193.2017.10.006 S932.5+1 :A :0253-4193(2017)10-0065-13 2017-01-09; :2017-05-13。 国家自然科学基金项目(31270527);南麂列岛国家海洋自然保护区管理局委托项目(NJKJ-2013-003)。 谢旭(1991—),男,安徽省宿州市人,研究方向为渔业资源。E-mail:1426619449@qq.com *通信作者:俞存根(1960—),男,教授。 E-mail:cgyu@zjou.edu.cn 谢旭,俞存根,蔡厚才,等. 南麂列岛海域蟹类群落结构及其与环境因子的关系[J].海洋学报,2017,39(10):65—77, Xie Xu, Yu Cungen, Cai Houcai,et al. The crab community structure and its relationship with environment factors in the Nanji Islands ground shallow sea area waters[J]. Haiyang Xuebao,2017,39(10):65—77, doi:10.3969/j.issn.0253-4193.2017.10.0063 结果










4 讨论
5 结论
猜你喜欢
热带林业(2022年4期)2023-01-10 10:44:04华东师范大学学报(自然科学版)(2021年2期)2021-11-10 03:10:35江苏农业科学(2021年3期)2021-04-29 16:12:13海峡姐妹(2018年11期)2018-12-19 05:18:30北京广播电视报(2018年39期)2018-11-23 06:01:14安徽农业科学(2017年26期)2017-06-11 16:38:25现代营销·经营版(2017年3期)2017-05-17 10:19:46东南文化(2016年4期)2016-08-22 09:35:42作文周刊·小学五年级版(2016年1期)2016-06-29 03:31:18中学生天地(A版)(2015年7期)2015-08-06 05:39:07
