
生态模型在渔业管理中的应用
2017-10-09 01:05:58张崇良陈勇韩东燕邢磊任一平
海洋学报 2017年10期
张崇良,陈勇,2,韩东燕,2,邢磊,任一平*
(1. 中国海洋大学 水产学院 渔业生态系统监测与评估实验室,山东 青岛 266003; 2. 缅因大学 海洋科学学院,美国 奥罗诺,04469)
生态模型在渔业管理中的应用
张崇良1,陈勇1,2,韩东燕1,2,邢磊1,任一平1*
(1. 中国海洋大学 水产学院 渔业生态系统监测与评估实验室,山东 青岛 266003; 2. 缅因大学 海洋科学学院,美国 奥罗诺,04469)
随着人们对于海洋生态的认识逐渐深入,渔业等人类活动的影响受到了更多的关注,“基于生态系统的渔业管理”(EBFM)这一概念被越来越多的研究者和国际组织所接受。生态模型为EBFM的实践提供了科学的评估方法和技术工具。与单鱼种评估模型相比,生态模型更多地考虑了系统内的各种生态过程和作用机制,栖息地等环境条件的变化,以及生态系统整体的结构和功能等,依此来反映生态系统的动态变化。由于生态模型是多样化的,关注和涉及不同的目标对象、机制过程、模型结构和参数,本文按照生态模型所关注的生态层次和组分将其分为多物种模型、群落结构模型和生态系统模型3个类型,分析了各类模型的优劣点。本文进一步阐述了生态模型在渔业管理中的应用领域,以管理策略评估为中心的应用方式,以及实践中模型选择和构建所需要注意的问题,还探讨了生态模型中降低不确定性和提高预测能力的方法。
基于生态系统的渔业管理;生态系统效应;管理策略评估;不确定性;预测能力
1 生态模型的研究背景
1.1 渔业危机
渔业的可持续发展是渔业资源开发和海洋环境保护的中心问题[1—3]。近年来随着人类活动对海洋环境影响的日益加剧,资源衰退和生态恶化等问题越来越严重。FAO公布的世界渔业统计数据显示,在全球捕捞努力量(CPUE)持续上升的形势下,捕捞渔业总产量自20世纪90年代逐渐持平或稍有下降,且其中充分和过度开发的资源比例逐渐升高,总体CPUE下降显著[4]。同时很多研究报道了世界范围内渔业资源崩溃现象[5—11]。例如,Mullon等[6]根据FAO渔业数据库,评估了1950年后的捕捞数据,在 1 519个渔业资源群体中,366个种群发生了崩溃。Myers和Worm[11]评估了全球13个海洋生态系的渔业资源状态,指出其中捕食性鱼类的生物量降低了超过80%。渔业活动对非目标种类也具有重大影响,如兼捕和混捕可能是导致一些濒危物种高死亡率的主要因素[12];底层拖网导致的海底环境改变,也破坏了众多底栖动物的栖息地[13]。而在另一方面,由于海洋生态系统的失衡,某些物种的数量可能在短时间内激增造成严重生态灾害,如赤潮、水母、浒苔等暴发[14—16],严重影响着滨海景观和海洋生态系统健康。海洋渔业与环境危机一方面与近年来全球变暖、海洋酸化和ENSO等海洋环境变化密切相关[17—20],另一方面则很大程度上受到过度捕捞和栖息地破坏等人为活动的影响[21]。这些因素的综合作用可能扰乱海洋生态系统的稳定性,使得生态系统结构和功能急剧变化并难以恢复[22—23]。应当注意到,渔业不仅是一个经济问题,更是重要的社会问题,涉及到生态系统服务、生存环境质量、食品安全和渔民就业等方方面面,渔业资源的崩溃可能会对国民经济和社会安定造成严重影响。合理的开发、保护和管理渔业资源是维护海域环境健康和生态系统稳定的重要基础,也是应对当前全球性渔业危机的关键。
1.2 EBFM
渔业资源的监测、评估和预报是渔业管理的基础。传统上,渔业资源评估一般基于单鱼种种群模型,如剩余产量模型(surplus production model)、实际种群分析(virtual population analysis)和种群综合分析(stock synthesis)等[24]。这些评估模型为我们认识渔业资源的变动规律提供了有力的工具,但同时需要注意到模型本身也具有一定的局限性。根本上讲,单鱼种评估模型以及对应的管理方法聚焦于渔业的直接影响,即捕捞死亡率对群体组成、繁殖和补充等的作用。其管理目标通常追求产量最大化,而忽略了渔业的间接效应,如渔业活动对栖息地破坏,生态系统内部捕食竞争等物种间关系,食物网结构变化,社会经济因素等的影响[25]。对于大多数种类来说,捕食死亡率和食物竞争是引起鱼群资源变动的重要因素,而这些因素取决于群落组成结构并随着资源开发过程产生相应变化[26—27]。此外,在多种类混合的渔业中,由于兼捕的存在,基于单鱼种模型对每个物种分别实施MSY的管理策略是不可行的[28—29]。因而单鱼种模型在许多实际问题中是不适用的,可能导致渔业资源过度开发甚至枯竭等严重后果,因此在制定长期渔业管理目标和参考点时必须慎重考虑[30]。
随着人们对于海洋生态认识的逐渐深入,渔业等人类活动的生态效应受到越来越多的关注。对海洋生态系统的深入认识启发了海洋管理思路和方式的演变,并催化了渔业的生态系统方法(ecosystem approach to fisheries, EAF)或基于生态系统的渔业管理(ecosystem-based fisheries management, EBFM) 的概念。简而言之,EBFM是一个综合的管理框架,涵盖了系统各组分的交互作用和环境的影响,将人类作为生态系统的一部分,旨在维持生态系统健康和实现可持续的生态系统服务[31]。尽管在原则上被广泛认同,而且生态学原则在渔业研究和管理中也受到越来越多的关注,然而全球范围内EBFM的实践仍非常局限[32]。这主要是因为,EBFM原则在渔业管理中的可操作性、成本、技术复杂性和实施效率有很大的限制[33-34]。在过度捕捞状况下,无论EBFM和单鱼种管理的首要目标都是降低捕捞努力量,EBFM相对于传统渔业管理方式并无优势[35]。
1.3 生态模型
目前对于EBFM的研究仍处于初级阶段,其相应的原则、目标、技术和方法尚不成熟。为了能够取代当下单鱼种管理范式,EBFM的实践中需要在生态系统层次建立明确的、可操作的管理标准和参考点[36—37],这依赖于科学的评估方法和技术手段,而生态模型则是其中的有力工具。与单鱼种评估模型相比,生态模型更多的考虑了系统内的各种生态过程和作用机制,例如物种间竞争捕食等相互作用,栖息地等环境条件的变化,以及生态系统整体的结构和功能等[38],能更真实地反映生态系统的动态变化。由于海洋生态系统是一类典型的复杂自适应系统[39],系统内各个组分之间的交互作用可能产生复杂的生态反馈机制,因此简化的、依赖直觉的渔业管理策略可能会产生难以预料的后果[27,40]。渔业管理中的参考点和控制规则必须随系统过程而产生动态变化,而生态模型作为研究生态系统结构和机制的主要工具,为生态系统评估提供了理论框架,在该研究方向具有明显优势[25]。
近10年间,生态模型领域有着较多的发展[38]。但是,由于目前的生态模型是如此复杂而多样化,很难全面地对现存模型作充分的总结和分析。本文仅对应用较为广泛的渔业生态模型进行分类,讨论各类模型的优劣点,但对模型的结构和参数等不作详述。另一方面,由于模型的多样化,模型的选择、构建和应用成为关键性问题。因此,本文将阐述生态模型在渔业中的应用方式,以及模型选择和构建所需要注意的问题,分析模型的发展方向,为EBFM提供研究思路。
2 生态模型的类型
生态模型狭义上指研究生态学问题的数学模型方法,本论文以此概念区别于传统单一的鱼种评估模型,指“包含了物种间竞争捕食等生理过程和生态机制,考虑环境条件变化以及社会经济因素对生态系统影响,以描述生态结构和功能动态变化”的一类模型。需要注意的是,单鱼种模型在更新发展过程中也越来越多的考虑到了物种间的捕食关系和温度等环境变化的影响[41—42],但其研究关注的仍是单目标鱼类,在研究目标上与生态模型有所区别。
生态模型是多样化的,涉及不同的目标对象、机制过程、模型结构和参数数量等。许多研究从不同的角度提出了模型的分类方式。例如Kerr 和Ryder[43]综述了渔业中的生态研究方法,并按其中对生态系统的解析程度大致分为4类:(1)经验性多元分析模型,主要基于数据统计,较少涉及系统过程;(2)多物种动态模型,考虑了主要物种(渔业的目标种)的种间关系,不涉及环境条件的变化;(3)系统简并模型,考虑了生态系统内部的能量流动和物质平衡,并以功能群的形式简并物种组成;(4)系统综合模型,动态模拟系统内部自下而上和自上而下的生态驱动机制,模型一般包含复杂的生物相互作用机制和物理耦合过程。Hollowed等[25]沿袭了Kerr的分类方式,并将生物-物理耦合模型等更多的模型纳入到该分类框架。Whipple 等[44]根据模型构建的技术方法将生态模型分为4类:(1)经验模型,依赖于变量之间的统计学相关性,主要用于生态模式识别、过程探索和参数估算;(2)定性生态模型,用于描述食物网联结结构,不涉及量化过程;(3)静态定量模型,用于描述食物网中的能量流动和群落粒径谱组成;(4)动态定量模型,用于模拟种群以及完整生态系统动态。按模型其他方面的属性,生态模型有着多样化的分类方法。Bolker[45]按生态模型的应用方向和建模数学方法,列举了一系列的二分性模型类别(表1),其中包括理论性与应用性的区分,定性/定量的过程,动态/静态的处理,以及数学推导/数字模拟计算等不同建模方式。

表1 生态模型不同研究目标和建模方法的分类[45]
本文参考了FAO 2007研究报告简化了生态模型的分类[38],按照模型所关注的生态层次和组分大致将其分为3类:(1)多物种模型(multi-species model),(2)群落结构模型(community model),(3)生态系统模型(whole ecosystem model)。其中每个类型中按其模型构建方式分为静态/动态和定性/定量等多个子类型(表2)。需要注意的是该分类并不是绝对化的,因为一些模型可以通过信息简并或具体化以及模型耦合来包容不同生态组分和生态层次。以下简要介绍各类模型的主要特点。
2.1 多物种模型
该类模型关注主要物种间相互作用,特别是种间捕食所造成的死亡率。模型一般针对于某些目标种类而构建,涉及对目标种有重要生态影响的有限种类。模型包含的物种数相对较少,一般少于10种[38]。这类模型包含多种子类型,例如:
(1)符号有向图(signed directed graph)是一种定性的生态模型。这类模型主要考虑到在数据有限的情况下,量化模型难以拟合生态系统不同组分和尺度下的相互作用,并针对该问题提供了一个简化的处理方式,避免了量化过程中对数据精确性的过高要求,因此该方法适用于数据稀缺的海域。符号有向图描述了生态组分间的直接和间接的关联性,以及系统结构的稳定性。Dambacher 等[46]利用符号有向图方法构建了一系列简化的生态模型,包括核心生态系统、杂食性系统、蜂腰型系统、兼捕渔业系统和多目标渔业系统等。这类模型为研究生态系统的反馈机制提供了有效的工具。此外,模型能够包含经济和社会组分,为综合性的管理策略的制定提供了参考[47]。符号有向图的缺点是精确度较差,并且模型隐含的假设是生态系统处于平衡态以及生态组分的关系为线性,这些假设条件削弱了模型的真实性[44]。
(2)非线性预测模型(nonlinear forecasting model)又称作非线性时间序列分析(nonlinear time series analysis),该方法利用时间序列数据来解析生态系统内部复杂动态关系。此类模型不依赖于具体的数学公式,而是利用状态空间重建(state space reconstruction)方法,将系统的不同状态变量视为多维空间,利用单个或多个变量的滞后性时间序列体系(time-delayed coordinate system)来重构整个生态系统的吸引子空间并预测其动态变化[48]。结合多元变量的系统重构可为研究整个生态系统的结构机制提供有力工具[49]。在渔业中该方法可以结合捕捞和相对丰度数据来预测渔业动态[50]。模型的优势在于能够处理复杂的、非线性的系统动态,且避免了大部分数学模型中涉及的参数不确定性;缺点在于其预测能力强烈依赖于数据的准确度,仅能进行短期预测(少于5年)[51],且一般不能预测发生在建模状态空间外的情况。
(3)多物种Lotka-Volterra模型是对传统捕食者-被捕食者模型扩展。该类模型以微分/差分方程描述物种间的相互作用以及渔业对种群的影响,利用数学分析和数值积分方法求解各个种群丰度或生物量的动态[52]。模型的主要假设是物种间的相互作用可以由Lotka-Volterra参数表征。Pimm和Rice[53]利用该模型研究了多物种、多生活史阶段的食物网结构稳定性,分析了多物种模型中生活史杂食性(life-history omnivory)、亲体-补充量关系以及食物丰富度对食物网稳定性的影响,其研究与传统模型结果显著不同。多物种Lotka-Volterra模型的优势在于结构简单,所需参数较少,对种群资源量和渔业产量的稳态分析较为简洁明了;其缺点在于某些参数如作用强度系数和总容纳量等可能较难获取。
(4)多种群评估模型将物种间的相互作用加入到剩余产量模型、实际种群分析和补充量模型等传统种群模型当中,例如多物种实际种群分析MSVPA模型 (multi-species virtual population analysis), 多物种统计模型MSM(multi-species statistical models),多物种产量模型(multi-species production models),汇总产量模型(aggregated production model),和多物种YPR模型(multi-species yield per recruitment model)等[54-58]。此类模型涉及的生物交互作用主要有两方面[59]:捕捞交互作用也称技术性交互(technological interactions),即同一网具对不同物种的兼捕;生物交互(biological interaction),主要涉及捕食死亡率以及生长速率的变异性。这类模型一般将自然死亡率区分为被捕食死亡率和其他因素导致的背景死亡率,并将前者与其捕食者种群动态相关联。因此在模型拟合过程中除了需要常规的年龄结构捕捞数据外,还需要胃含物分析数据以确定主要种类间摄食关系的强度。多种群评估模型的需要较多的数据确定摄食作用关系,且作用强度固定,不涉及食物选择性变化等过程。
(5)多物种空间结构模型(spatial-structured multispecies model)将物种的空间分布加入到多种群动态模型当中,模型组分和结构较为复杂。例如MULTSPEC是模拟Barents海域的一个包含鱼类长度、年龄和空间分布的多种类模型[60],其生态组分包含鳕鱼、鲱鱼等多种鱼类以及海豹和鲸等种类。该模型说明了鲸的种群数量能够控制鲱资源量,而海豹数量则对胡瓜鱼和鳕鱼种群有重要影响。BORMICON (boreal migration and consumption model)是模拟极地生态系统一个类似的模型,也包含了年龄结构和生物的洄游习性以及捕捞努力量的空间分布[61]。GADGET (globally applicable area-disaggregated general ecosystem toolbox)是在前述两者基础上的继续发展,其结构更为灵活[62]。该模型进一步细化了种群的体长和年龄组成,考虑了不同空间和时间尺度,并能够容纳不同类型的生长、繁殖和捕食关系等。该模型在欧洲的多个海域(如Celtic Sea, Icelandic waters, southern Benguela 和 North Sea)有着较为广泛的应用。这类模型的优势是模拟了某些物种特别是海洋哺乳类的洄游习性,为研究鱼类资源量变化与渔业活动和哺乳动物分布的时空匹配问题提供了有力的工具;同时建模需要数据量较大,特别是需要生物空间分布特征数据较难获取。
总体而言,多物种模型在理论上较为简单,对模型参数的不确定性有着较好的控制,能够较为准确的做短期预测;另一方面,模型仅关注目标种类,包含的生态组分较少,忽略了一定的生态真实性,在涉及气候变化等长期预测方面可能效果较差。
2.2 群落结构模型
群落结构模型一般包含生态系统内主要的生物种类和其生态关系,与前一类模型相比,该类模型一般涉及更多的物种数(大于10种),包含更为复杂的种间关系,但较少涉及水文、底质环境等非生物因素。模型的理论基础更多的涉及生态学理论而非种群生物学认识。
(1)多元统计模型是最简单的一类群落结构模型。这类方法在群落结构分析中较为常见,包括聚类分析、多维标度排序、主成分分析、对应分析、冗余分析和典范对应分析等[63]。这类模型以经验性观测数据描述变量间的相关关系,以及环境变量对于各个物种的影响,其特点是一般不包含物种间的相互作用,也较少用于用做预测,在此不做详细介绍。近来出现了另一类基于多元统计的群落结构模型,称物种联合分布模型(joint species distribution model),可同时分析生物对环境的响应以及物种间的交互作用[64—65]。该类模型主要通过混合效应模型(generalized mixed model)和隐变量模型(latent variable model)方法实现,目前其理论和方法还处在发展过程之中。这类模型对于了解生物时空分布规律和评估种间作用关系具有重要意义。
(2)粒径谱模型(particle-size models)是基于群落内个体大小(即粒径),如长度质量组成的规律性而构建的生态模型。该模型将群落内不同物种按个体大小分为粒径组,计算各个粒径组的生物量或者丰度,并分析其分布规律。粒径谱模型的理论基础是食物网的能量传递效率,相关研究表明,丰度或生物量与所其对应的粒径组在对数尺度下能够形成规则的线性关系,即粒径谱或生物量谱[66—67]。Moloney和Field利用粒径谱模型研究了浮游生物与渔业种群生产力之间的关系,并与该种渔业的实际产量数据做了比较[68],并证明粒径谱方法能够有效估算渔业的潜在产量,其研究建议将此估计值作为渔业管理的底线。该模型的优势在于模型形式简单,所需参数较少,易与构建和使用。因而粒径谱模型能够应用于产量统计、鱼类生物学参数等数据稀缺的海域,并在一定程度上反映渔业活动对生态系统的影响。其缺点在于模型假设群落处于静态平衡,因而模型仅能反映稳定的渔业状况,难以预测捕捞强度变化后的系统动态[44]。
(3)Size-spectrum model[69]与粒径谱模型在名称上类似,但在原理和构建机制上截然不同(为便于区别,以下均以size-spectrum model表示)。这类模型以生物个体大小和能量收支过程为基础,以微分方程的形式描述了新陈代谢、竞争、捕食和生长、繁殖、死亡等具体生理过程。该类模型主要关注个体大小的生态学意义,即个体大小与新陈代谢速度、摄食选择性、营养级以及自然死亡率的关系,而弱化了分类学上的物种特征,这与传统基于物种的生态模型有显著不同。根据模型的复杂程度和结构细节,该模型大致可以分为3个类型[70]:简化群落模型(community model),将群落中所有物种简化为单一种群并忽略不同物种生活史参数的差异;特征模型(trait-based model),以单一特征如渐近体长、性成熟体长等体现不同物种的差异,食物选择性仅由个体大小决定;物种模型(species-specific model),使用较多的参数分别描述每个物种的生活史参数和食物选择性。这类模型有着较为广泛的应用,例如Hall构建了基于体长组成的群落结构模型LeMANs(length-based multispecies analysis by numerical simulation),研究了不同生态学指标对于水平的捕捞死亡率的敏感度[71]。Blanchard等以基于个体质量的群落模型(mizer)模拟了欧洲北海鱼类群落,该模型对于实际渔业动态和群落结构有着较好的拟合[72]。Maury和Poggiale根据类似的理论框架构建了复杂的群落模型,并包含了温度对不同生活史参数的影响[73],研究探讨了生物多样性和密度制约机制对于群落稳定性的影响。该类模型的特点是以捕食者-被捕食者的相对个体大小来决定摄食选择性,以此描述机会性捕食关系和食性转变等生态过程,这在常规的食物网研究中难以实现。此外,由于渔业中网具选择性也取决于个体大小,因此size-spectrum model为研究渔业选择性的生态影响提供了便捷的工具。
(4)基于个体的模型IBM(individual-based model)或基于代理的模型ABM(agent-based model)从个体水平构建了一种由底至顶的生态系统模拟方法[74—76]。这类模型描述了个体的整个生活史动态,并基于一系列简单的个体行为或决策的规则模拟个体行为的变化对整个生态系统的影响。二者在模型构建思路上基本一致,其不同之处在于,ABM以鱼群等超个体结构为模拟单元,其中的agent包含不同的属性特征以及行为模式,如繁殖和洄游分布规律等。如Shin 和 Cury构建了一个基于个体的多层次、多物种模型OSMOSE (object-oriented simulator of Marine ecosystem exploitation)[77],该模型中的种间捕食关系取决于个体的体长比以及其空间分布重叠程度,且鱼卵和幼鱼的死亡率或补充量均受捕食关系调控。该模型已用于生态学指标的评价[78]和渔业管理策略评估[79—80]等。Gray等构建了基于代理的生态模型 InVitro[81],并将之用于澳大利亚近岸海域的渔业管理策略评估 (http://www.cmar.csiro.au/research/mse/invitro.htm)。InVitro可以包含多种不同类型的agents以实现对整个生态系统的模拟,如海洋理化环境、初级生产者、游泳/固着生物以及人类活动。相较于其他模型,这类模型更能反映个体层次的生态特征如游动能力、生长速度、繁殖习性以及生物间的相互作用对生态系统动态的影响,但随着生态组分种类和数量的增加,模型构建的难度迅速增大。
总体而言,群落结构模型更好地反映了真实的生态过程,能够模拟生态系统内较为复杂的种间关系,以及捕捞等人为活动的长期影响。模型的复杂程度在不同类别之间差别很大,对于结构较为复杂的模型特别是个体模型的不确定性较难评估和控制。
2.3 生态系统模型
生态系统模型包含了从初级生产者到顶级捕食者在内的全部营养级、生物所需的栖息环境和营养化学元素循环动态以及气候变化和人类活动的影响等。也有研究称之为end to end模型[82—83]。
(1)EwE(ecopath with ecosim)是目前应用最为广泛的生态系统模型[38]。EwE模型最初由Polovina创建[84],并逐渐发展为泛用性的生态系统模拟工具,主要用于评估渔业的生态效应以及渔业的管理策略[85]。EwE模型基于食物网结构研究,以物质平衡为限制条件,量化了生态系统能量流动和物质循环过程。根据对生态系统时空解析度,EwE模型可划分为Ecopath、Ecosim和Ecospace 3个类型。其中,Ecopath模型描述了生态系统静态特征,即物种间的营养交互作用和系统内的能量流动速率。由于该模型反映的是系统的静态结构,因此主要用于研究生态系统能量流动效率和营养结构,以及生态参数的估计和不确定评估,较少用于系统动态预测[86]。Ecosim中加入了捕食者和被捕食者的种群动态,以及生长率、死亡率和迁移率等过程参数,并按摄食的难易度将被捕食者分为两类,模拟了栖息环境庇护机制(prey refugia)。Ecosim模型能够模拟生态系统的动态变化,适于研究渔业活动对于系统状态的影响,特别是单鱼种模型中无法处理的多种类交互和多管理目标冲突问题[87]。Ecospace模型包含了生物的空间分布动态,可以模拟栖息地适宜度和摄食效率的空间异质性,以及捕捞努力量的空间分布等,可用于指导海洋保护区构建以及鱼类洄游机制的研究。EwE有着较好的易用性,其软件在持续发展中(http://ecopath.org,版本6.5,2016年7月),此外Ecobase数据库的建立也方便了不同生态系统中EwE模型的比较。Heymans等综述了EwE模型构建、检查、校正以及模型间比较的原则,特别是蒙特卡洛模拟在处理参数不确定性方面的应用[88]。
(2)生物地球化学系统模型(biogeochemical system model)是一类具有复杂结构的完整生态系统模型。与其他大部分模型不同的是,该类模型不以生物量为基本模拟变量,而是关注了碳、氮、磷、硅等营养元素在生态系统中的传递和流动过程。生物地球化学系统模型在其发展过程中衍生出了多个子模型。其中ERSEM (the european regional seas ecosystem model)模拟了North Sea表层和底栖生物群落中碳、氮、磷、硅的周年循环过程[89]。模型包含了浮游动植物、微生物和碎屑等生物组份,以及日照、温度、大气氮输入、水动力过程以及河流输入等环境变量;IGBEM (integrated generic bay ecosystem model) 整合了ERSEM模型,包含了更多的物理过程,并具有水体、底表和沉积物3个层次的空间结构[90]。该模型应用于澳大利亚Port Phillip Bay中渔业和富营养化对于生态系统的影响。ATLANTIS模型是在IGBEM上的进一步发展,涵盖生物地球化学循环等完整的生态过程[91]。该模型追溯了氮和硅等元素在生态系统中的流动过程,并涵盖了自初级生产者到高级捕食者以及碎屑在内的完整营养结构,以及生产、消耗、捕食、补充、洄游、栖息环境选择以及自然和捕捞死亡等过程。模型中初级生产者和无脊椎动物组分以总生物量表示,而脊椎动物则划分了年龄结构。ATLANTIS模型以地理多边形描述了生物的空间分布特征,与一般模型不同的是,该模型可在垂直方向分层模拟三维的空间结构。此外,该模型还可以模拟不同的渔具选择性,作业渔船的分布和不同的管理方式等。该模型为研究气候变化和人类活动对生态系统的影响提供了强有力的工具[92]。通过与其他模型的耦合,ATLANTIS模型也可以用于优化渔业的经济和社会效益,权衡生态-经济-社会等不同方面的管理目标。目前ATLANTIS模型已被应用于全球超过30个海洋以及湖泊生态系统(http://atlantis.cmar.csiro.au)。该模型结构复杂、参数繁多,需要大量的数据支撑,大部分数据稀缺的海域难以满足其模型构建的需求。此外模型运行所需时间较长,模型的验证和不确定评估较难进行,对于群体补充过程等假设条件较为敏感。
总体而言,生态系统模型涉及了从生物到环境因素的各个方面,能够更为有效的评估大尺度气候变化和人类活动对生态系统的影响。但模型构建所需的数据较难获取,需要整合多方面的研究资料,受限于对生态系统的理解和计算机的运算能力,一般需要对生态系统的某些部分进行简化处理。此外模型的验证一般较为困难,这也大大限制了此类模型的应用。

表2 渔业生态学研究中常见的生态模型
3 模型应用概述
3.1 应用领域
无论是单鱼种模型、多物种模型或生态系统模型,在渔业管理中的根本目的是一致的,即理解渔业活动对渔业种群和生态系统的影响,并为管理者提供决策依据。不同之处在于,生态模型涉及更多的生态组分和交互作用机制,因而可以探讨的问题和应用范围更为广泛。概括而言,生态模型在研究中的应用体现在以下方面[38]:
(1) 生态机制研究:
理解生态系统结构和功能,以及系统动态的驱动机制;
理化气候变化对于渔业资源的影响,特别是对群体补充的影响;
自然灾害如赤潮、水母爆发对于生态系统结构和功能的影响;
外来种入侵的风险和可能造成的影响等。
(2)渔业影响研究
拖网等作业方式对栖息地破坏,以及栖息地变化对于种群的影响;
渔业对生态系统平衡、抵抗力和恢复力的影响;
捕捞目标种的选择,及目标种的移除对生态系统其他组分的影响;
对于非目标鱼类的直接和间接生态作用,包括兼捕的影响;
饵料生物的移除对于其捕食者的影响,以及捕食者对于摄食条件的适应性;
海洋鸟类和哺乳类与渔业的关系;
(3)资源管理与养护研究
近岸人类活动如海港工程、溢油事故、陆源污染对濒危物种以及生态系统的影响;
分析单鱼种评估模型的偏差,以及基于单鱼种管理策略的风险;
从生态和经济学角度上优化渔业的捕捞努力量、渔具选择性以及渔业的时空分布;
渔业种群的重建过程中的主要影响因素;
增殖放流的效果评估以及放流群体的生态影响;
海洋保护区构建的生态原则与实践;
生态模型在渔业管理当中研究的目标是多样的,但其应用方式基本一致,主要通过模拟渔业开发的后果来评估不同管理策略的有效性。其中一种行之有效的方法被称为操作性管理规程(operational management procedure,OMP)[101]或管理策略评估(management strategy evaluation,MSE)[102]。完整的 MSE过程一般包含3个组分:操作模型(operating model)、观测模型(observation model)和评估管理模型(management/implementation model)[103]。其中操作模型用来模拟渔业种群和生态系统的关键过程和动态变化机制,以代表生态系统的“真实”情况;观测模型用来模拟实际的科学调查和监测中难以避免的偏差和误差,以代表通过观测从生态系统获取的“真实”数据;评估管理模型模拟了管理策略的实际执行情况,以及其中可能存在的偏差,其中对于渔业来说即为渔获控制规则(harvest control rule)。MSE的基本步骤包括:(1)设置管理目标;(2)明确管理策略中可能存在的不确定性;(3)构建操作、观测和管理模型,需要考虑不同的模型以涵盖不同来源的不确定性;(4)明确在实际中可行的管理策略;(5)利用模型模拟不同的管理策略;(6)分析统计结果,优化管理目标。Punt等对MSE的整个过程做了详尽的阐述,以指导其具体应用[102]。MSE与一般资源评估的不同之处在于,评估管理的结果可反馈到操作模型当中,以不断改进对管理策略,优化下一步MSE的效果。由于渔业本身涉及到环境、生态、经济和社会等多个方面,单一的管理措施往往很难满足多样化利益相关者的需求,而基于生态模型的MSE则能够为不同目标的权衡和管理措施的制定提供科学的依据。此外, MSE中多个模型联合使用,能够更好的体现多种来源(如环境/观测/管理中)的不确定性,并为生态监测的设计和数据分析方法的选择提供指导[104]。
基于生态模型的MSE在指导渔业管理中具有显著地优势,这首先体现在MSE中能够进行各种管理策略的(模拟)试验。涉及到海洋渔业的问题,研究的时空尺度往往非常宏大,控制性试验条件一般很难实现;此外由于涉及到渔民和其他相关者的生存和经济利益,以及对濒危物种的可能危害,一些试验性的管理策略在社会层面也无法开展。MSE则对上述问题提供了行之有效的解决方案。其次,通过对不同管理策略的可行性、潜在后果、对不确定因素的稳健性等方面的评估,MSE可以很好地将很多利益相关者纳入到管理系统中来,从而增加了管理政策的透明度和接受度,这对管理策略的成功实践至关重要。其三,MSE中明确地提出了不同管理目标的权衡,综合考虑了生态过程、取样、测量和政策执行中的各种不确定性。通过系统动态与管理单元间的反馈而不断更新管理目标和管理策略,而形成适应性管理策略[103]。MSE的缺点在于,构建整个系统花费的时间较长且费用较高,这很大程度上限制了其在某些生态系统中的应用[105]。此外,MSE中涉及较为复杂的模型模拟过程,某些输入参数的可靠性对其结果有着重要影响,特别是其不确定性的估计范围可能决定了最终管理策略的选择[44]。
3.2 模型选择与构建
需要指出的是生态系统本身是复杂的而多变的[39],尽管海洋生态学和生态模型已经经历了较长时间的发展,总体而言我们对于生态系统的认识仍非常有限。从全球范围来看,目前生态模型在EBFM中的应用还是比较局限[106],其关键问题是生态模型能否反映出生态系统的真实过程,即模型的可靠性问题[107]。尽管复杂生态模型包含了更多的物理和生物因素,能够更真实的反映了生态系统的变化过程,但生态系统过程是相互联系的,而研究中难以将大量的生态过程都纳入模型当中,因此建模过程中需要对关键过程权衡取舍。同时模型所需要拟合的参数随着模型结构的复杂度迅速增多,因此过度拟合(model over-fitting)在生态模型研究中是一个广泛存在而有严重影响的问题[108]。在有限的数据下,模型的结构和参数具有很大的不确定性[109—110],预测结果的精确性相对较低。生态模型的复杂性使其预测结果很难验证,同时大量的假设条件也降低了生态模型在管理者群体中的接受度。许多个不同研究领域的共识是,大型的复杂生态模型的预测能力并不一定优于简单的模型,就渔业资源评估而言,其精确性和预测能力并不一定优于单鱼种评估模型[111]。换言之,在目前的研究中,单鱼种模型和生态模型具备各自的优势和劣势,不能够相互取代,更有的研究者甚至认为,最好的预测模型不应该涉及生态机制,而是完全依赖数据中体现的统计学关系[111]。
总结来说,在当前的发展水平下,生态模型还不能够完全真实地反映生态系统的全部过程[112],但模型对于生态系统部分过程的真实反映依然能够为渔业和海洋管理提供依据。不同模型往往对生态系统的不同组分有所侧重,因此它们能够准确模拟和解答的生态问题也存在差异,因此在实际应用中需针对具体的科学问题,选择合适的生态模型。Levins提出一个模型选择框架[113],其中涉及3个必要属性之间的权衡,即真实性(realism)、精确性(precision)和普适性(generality)(图1)。Levins指出,(至少在目前)这三者是无法同时达到的,实际的模型仅能在3个属性中实现其二,根据三者间的不同权衡策略可以划分3类模型,即体现生态真实性和精确性,但适用范围较为局限的经验模型(empirical model);体现精确性和普适性,但对生态系统加以抽象和简化的分析性模型(analytical model);和体现普适性与真实性,但往往带有很大不确定性的机制性模型(mechanistic model)。
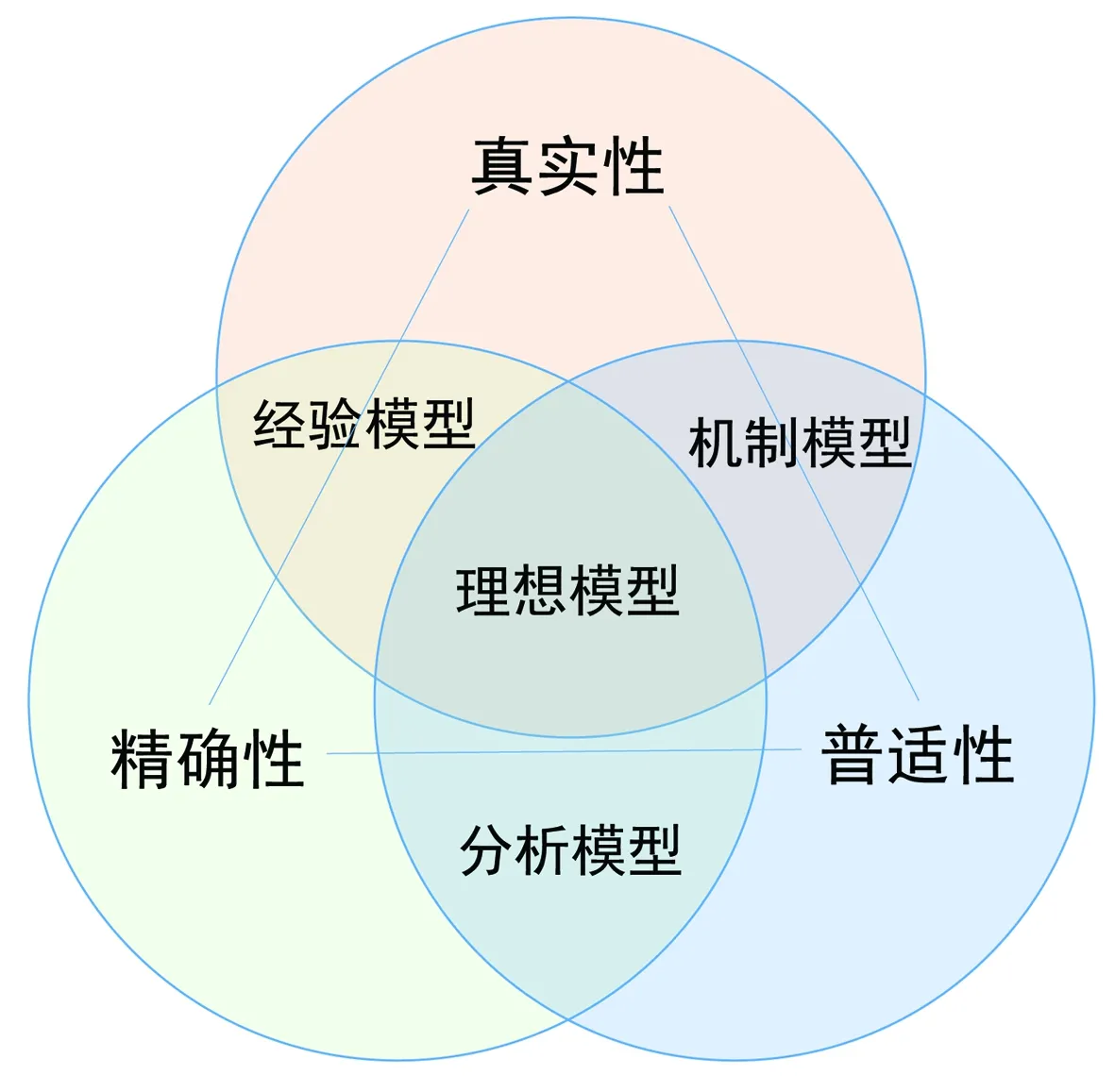
图1 Levins的三分法模型选择框架[113]Fig.1 Levins trichotomous model classification scheme[113]
尽管一些研究对Levins的三分法提出了质疑[114],但总体而言,三分法为生态模型的选择提供了较为直观的理论框架。在渔业资源评估与管理的应用中,该框架可简化为两类科学问题[115],即策略性(strategic)应用问题和技术性(tactical)应用问题。其中前者对模型的生态真实性和普适性要求较高,对应三分法中的机制性和分析性模型。渔业中的策略性问题主要涉及长期的和大范围的管理规划,如预测气候变化对渔业的影响,渔业管理目标的评估,渔业政策效用评估等方向性的问题。后者则对预测的精确度有更高的需求,对应三分法中的经验性模型。技术性问题主要涉及生态系统某个方面的精细动态,而忽略模型时空尺度之外的相关过程,用于制定短期的或特定区域的管理决策,如捕捞努力量和TAC等具体指标。针对于具体的科学问题合理选择是生态模型成功的关键,这就要求根据研究目标在模型的生态结构、系统误差、观测误差以及经济成本等方面权衡利弊[30]。
生态模型的构建是一个复杂的过程,为了保证模型能够满足渔业评估和管理的目标要求,需要对模型的构建过程进行规划。简而言之,生态模型的构建一般遵循以下步骤[45]:
(1)明确模型的目标和预期解答的科学问题;
(2)根据科学问题选择模型的类型,首先构建概念模型,明确模型所需要涉及的生态组分和过程;
(3)分析模型拟合对数据的数量和质量的需求,综合调查数据、实验分析、经验判断和相关文献资料等不同来源的数据。在数据有限的情况下优先考虑相对简单的模型;
(4)以数学形式分别描述概念模型中的生态过程、物种间的相互作用以及环境因素的影响等;
(5)估算模型参数。在相对简单的统计模型中,一般使用最大似然估计等方法求解算数;在复杂的生态模型中很多参数往往难以直接估算,而需借鉴相关研究的结果;
(6)模型验证。一般通过比较模型预测值与实际观测值来评价模型的真实性。交叉验证(cross-validation)等标准化方法为模型验证提供了更为客观的依据;
(7)模型敏感性和不确定性评估。模型参数直接影响了其预测结果的准确性,明确哪些参数对于预测结果有主要影响对于模型优化十分重要。该方法在单鱼种模型中有着常规的应用,而在大多生态模型中应用较少;
(8)模型的应用。结合MSE方法,解决研究目标中所提出的问题;多个模型的结合使用能够减小研究中出现严重偏差的风险。
除了建模的基本过程之外,结合具体的渔业管理问题,生态模型研究还需要注意以下几个方面:(1)模拟物种的数量以及年龄/体长结构;(2)模型的时空尺度和解析度;(3)是否涉及物种的洄游等空间结构;(4)是否涉及环境因素以及栖息地的变化;(5)模型是否能够准确反映生态系统短、中、长期的动态;(6)模型的易用性和可操作性;(7)现有数据质量和数量能否满足模型对于数据的需求;(8)模型的关键假设是否适合对科学问题的解析;(9)是否能够模拟特定的渔业管理策略。
为系统性展示生态模型的应用所涉及的各个方面,本文将整个应用系统分为4个模块,即监测系统、评估系统、管理系统以及外部变量(图2)。这几个系统以MSE为中心,通过数据收集、模型构建、验证以及渔业评估管理等相关流程,进行系统的反馈和更新,以实现适应性的渔业管理。
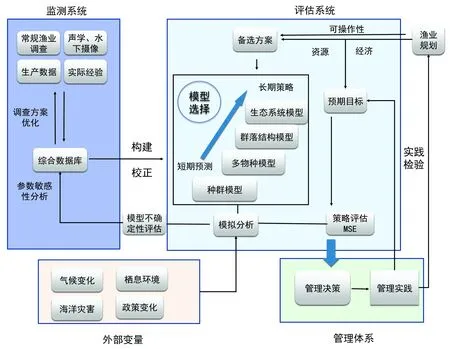
图2 以MSE为核心的数据收集、模型模拟以及渔业评估管理流程Fig.2 A flowchart of MSE-based processes for data collection, model implementation and fisheries assessment
4 生态模型的发展方向
4.1 降低不确定性
生态模型的主要作用在于理解生态系统结构和功能,以及预测生态系统动态。但目前的应用往往倾向于前者,因为模型的预测能力还有很大的局限[116]。模型预测的误差被归因于模型的不确定性,其产生的原因是多方面的,如生态系统本身的随机性、不同时空尺度上的异质性、数据的有限性以及复杂系统的混沌特征和涌现性(emergence)等。较高水平的不确定性一方面降低了模型的实际应用价值,另一方面也减弱了生态模型的可信度而大大降低了其公众接受度,并容易在实践中引发争议。因此,生态模型今后的发展中的一个关键问题就是降低模型的不确定性和管理失误造成的风险。
相关研究对生态模型不确定性的来源进行了分类,如过程随机性(process stochasticity),观测误差(observation error),模型结构的失真(model misspecification),拟合技术缺陷(technical uncertainty)以及参数不确定性(parameter uncertainty)等[109,117]。其中观测误差和参数误差一般可以通过丰富数据加以改善,这需要在今后研究中开展持续性和一致性的调查监测工作,以提高数据的质量与数量。需要指出的是,数据的收集工作应紧密联系模型对数据的要求,特别注意数据的类型、时间和空间分辨率和覆盖范围等。此外,由于生态模型往往涉及多个研究领域,不同领域之间的交流、借鉴和信息共享对促进数据的有效利用十分重要。然而在众多海洋生态系统中,由于社会经济发展水平、渔业的重要性以及公众的关注度等的限制,在建模中可用的数据是有限的,难以通过丰富数据的方式来提高模型的精度。此外,补充数据的方法自身也有一定的局限性,即该方法可以有效降低参数误差但对于改进模型结构误差作用不大。这种情况下就需要通过对现有模型的合理利用以降低其在实际应用中的不确定性。
由于在理论和结构上的差异,不同生态模型的不确定性水平是显著差别的,因此最直接地降低不确定性方法是合理选择模型,而模型的复杂度是其中主要的参考标准。Collie 等综述了在渔业管理中应用较多的生态系统模型[115],并按照模型的复杂程度从4个维度做了比较,即(1)种群时空、年龄和发育阶段的结构复杂性;(2)环境变化对于各营养级以及生长繁殖的影响的解析度;(3)直接、间接交互作用的物种数量;(4)捕捞和渔业管理等人类活动,以及经济社会因素的涉及深度。其研究认为,在4个维度上,随着模型复杂性的增加其偏差(model bias)减小,即复杂模型更能代表生态系统的真实过程;但其参数不确定性(parameter uncertainty)增加,即预测精密度下降。在从简单到复杂的中间梯度,即中等复杂模型(model of intermediate complexity)能够较好的平衡模型的精确度(accuracy)与精密度(precision),因而在实际的渔业管理当中更为有用[118]。
稳健性渔业管理是应对模型不确定性的一种适应性策略,该方法明确考虑了模型结构和参数的不确定性并将其纳入到模拟和决策当中。具体而言,稳健性管理评估了模型不确定性所对应的管理失误的风险,并将降低该风险作为管理目标之一,以权衡资源开发与生态保护等不同管理目标间的冲突[119]。稳健性渔业管理从原先的产量最优化策略,如MSY、MEY等,转变为适当性开发,或称预防性管理策略(precautionary approach),以避免或尽量降低渔业遭受重大损失的可能性。预防性管理策略在渔获控制规则上体现为更低的捕捞限额,以及更保守的种群生物量和捕捞死亡率的参考点和阈值等[120—121]。该方法在传统的单鱼种渔业评估中有着较为广泛的应用,但在基于生态系统的渔业评估中研究较少。
多个模型的综合使用是降低模型本身不确定性的方法[110]。该方法的应用是在研究中构建多个类型不同,基本原理、参数设置、假设条件和模型结构有较大差别的模型。如果不同模型的预测结果能够大致吻合,即说明模型的预测较为可靠。但实际操作中不同模型的预测结果往往不完全一致,这种情况下就需要对多个模型进行的比较和权衡,这种方法也叫多模型推断(multiple model inference)[122]。一方面,通过比较不同模型并与实际观测数据对照,可以检验哪些参数设置、生态过程和假设条件对于模型的预测更为关键,并识别出可信度较高的模型结果;另一方面,可以通过加权平均的方式综合多个模型的预测结果,以优化模型精确度。对于后者,权重选取的方式是多样化的,如简单平均、误差校正化平均、基于信息准则(AIC)的加权平均以及贝叶斯方法等。这些方法在其他学科如气象和海洋预报中有着常规的应用。
除模型比较和平均化方法之外,多模型的综合使用还有另一途径。由于不同模型聚焦于生态系统的不同组分,其模拟的优势存在差异,因此通过耦合多个模型可以在生态系统的不同层次和方面进行互补。一个简单的例子是,多物种生态模型能够较为真实地反应鱼类捕食死亡率,因此该模型可为单鱼种评估模型提供更为准确的自然死亡率估计值及其变化趋势,而后者可实现较为准确的短期预测以及不确定性评价[25]。实际上,由于生态系统是一种典型的复杂、非线性系统,其对于初始条件的敏感性使得任何单一模型的预测能力非常有限[51]。但原则上系统的非线性的动态和混沌行为都是由基本的生态学关系所支配,因此对系统的短期预测是可能的,这也是很多复杂自适应系统的共同特征[39]。基于以上观点,本文对多模型的综合使用的方法进行了整理,提出分层模型结构框架(hierarchical modelling framework),以结合单鱼种模型、多物种模型、群落模型和生态系统模型各自的优势。简单而言,该框架以完整生态系统模型为基础,分层次构建了复杂度不同的模型,并通过模型之间的耦合,以复杂的模型的指导简单模型的构建。分层模型过程主要包括:(1)根据完整生态系统模型分析生态系统各个组分的相对重要性,识别影响生态系统的关键因素以及气候变化等外部条件对系统的影响机制;(2)聚焦于关键生态过程构建群落模型或多物种模型,识别对目标种类具有重要影响的摄食和竞争种类,以及捕捞等过程的间接影响;(3)以单物种模型预测资源动态,并评估结果的不确定性;根据模型敏感性分析,改善单物种模型的结构并优化自然死亡率、补充量等关键参数,为渔业提供更为准确的预测。下一步的研究将进一步完善分层模型结构框架的理论和实践应用。
4.2 提高预测能力
随着气候变化和人类活动强度的加剧,全球范围内许多海洋生态系统的结构和功能发生了重大变化,这种现象称为“稳态转换”(regime shift)或“生态相变”(phase transitions)[123]。这种转变对于渔业生态学研究是一个巨大的挑战,因为在新的系统稳态下,依赖于过去观测数据的经验性规律可能并不适用。传统的渔业评估模型以及一些生态学模型一般以生长率和死亡率等统计学变化率(demographic rate)来描述种群动态,在环境条件相对稳定的前提下,这些参数的变化不大,模型能够给出较为准确的预测;但在生态条件有显著变化的情况下,该类模型并不适用。未来的研究需要考虑在稳态转换过程中生态系统动态,并评估生态系统的稳定性,以支持渔业资源的可持续利用。因此,生态模型未来发展的一个重要方向就是提高模型在生态系统发生重大变化情况下的预测能力。
Grimm 和 Berger研究指出,要提高模型预测能力需在建模思路上做出改变[124],即以基于过程的建模(process-based modelling)取代基于统计学的思路(demographic thinking)。这种建模方式不以生长率、死亡率等变化速率为基础,而是聚焦于基础层次的生态过程和个体的行为及个体间的相互作用。例如种群的死亡率由捕食选择性、栖息地分布以及水文环境等多个因素共同决定,而基于过程的模型明确描述了这些具体过程,并将总的死亡率表示为相关过程的涌现性结果(emergence)。生态系统的这种涌现性特征一般难以通过传统模型来模拟,而基于个体或代理的模型(IBM与ABM)为该方向的研究提供了有力工具。在这类模型中,基于过程的思路得以充分体现:生态系统的结构和功能取决于个体的特征和行为,系统整体的动态是个体状态、行为及其相互作用的结果[125]。IBM/ABM不仅可以有效描述生物对环境变化的响应,还能够很好地反映生物之间的交互作用,以及生物对非生物环境的作用。此外,IBM模型中还可以模拟生物对环境的适应性,这种适应性可能表现在行为的变化,也可以是基因上的改变。例如,渔业作为一种高强度的人工选择过程,对鱼类的种群组成和基因结构有着显著地影响[126]。上述特点使得该类模型特别适于研究环境变化对生态系统的影响以及生物对环境的适应,IBM类模型在未来的研究中必将受到更多的关注。
随着模型的发展,所需涉及的生态过程和机制越来越多,因此模型的结构和参数必然变得越来越复杂。从基础层次开始构建模型无疑是一项繁重的工作,而相关研究提出了以子模型为基础的模块化建模方式[124]。这种方法是将整个生态系统划分为相对独立的模块,以子模型分别描述不同生态组分和过程。子模型可能包括物种的生理特征、行为模式、摄食选择、能量收支、物种相互作用以及栖息地选择等特征。子模型的参数则根据观测数据或实验数据分别估算。因此,在建模过程中,研究者可根据研究目标选取关键子模型,根据实际观测数据对模型进行参数,最后通过模型间的耦合完成完整生态模型的构建。这种方法使得建模更加高效和节约成本。此外,不同子模型可能来自于生理、生态以及动物行为等不同研究领域,每个子模型在各自的领域能够被较好的检验和完善,因此整个建模过程更加可靠。但同时由于子模型的研究背景不尽相同,模型之间的耦合可能存在一定困难[82],这在今后的研究中也应引起注意。
5 结语
无论在学术界还是公众认识上,当下渔业科学的主题都是修复受损的海洋生态系统,养护渔业资源和实现资源的可持续利用。基于生态系统的管理是实现渔业资源可持续目标的必经途径,这也是相关研究的一个重要共识[30—31,36]。但由于数据、研究方法和技术等的限制,EBFM目前还很难切实有效的开展[32]。生态模型作为支撑EBFM的主要工具,其构建、优化、检验和合理应用是目前亟待解决的科学问题。同时也应认识到,由于生态系统本身极度的复杂性,生态模型的相关研究必然是困难的。从本文论述中也可以看出,生态模型相关研究的理论、方法和技术手段都远未成熟,本研究领域还有大量工作需要开展。我国在渔业生态模型领域有一定的研究基础[127—128],特别是EwE模型应用方面积累了丰富经验[129—130],但同时与国际前沿水平上存在一定差距。这一方面是由于历史原因,渔业科学相关研究时间尚短,经验积累不足,数据往往分散不连续;另一方面,以往的研究更多的倾向于鱼类生物学和生活史特征,在生态模型方面研究力量较弱,对于渔业资源的评估管理的关注度较低。但考虑到社会发展现状与未来需求,这种研究格局必须加以改变,特别是在我国着力建设海洋生态文明的大形势之下,研究渔业对于海洋生态系统的影响显得尤为迫切。
以生态模型为基础,解析生态系统结构和动态,预测渔业活动的直接和间接生态效应,合理的开发、保护和管理渔业资源,是维护海域环境健康和生态系统稳定的关键。未来的研究中需要更多的考虑生态、经济和社会等不同方面的需求,权衡短期与长期利益,协调开发与保护的策略[131]。在认识不足和不确定性的状况下采取稳健和预防性的管理措施[104],并构建群落和生态系统层次的管理标准、参考点和控制规则等。此外,相关研究也需将渔民和管理者等人文要素囊括在内,以确保管理措施的有效实施。尽管生态模型还有显著的不确定性,EBFM的实践还存在诸多问题,但充分认识其缺陷,在理论和实践上的不断探索,生态系统的管理框架将逐渐完善,这也是世界范围内渔业管理的必经之路。正如Tukey所强调的[132],“对一个正确问题的模糊回答远远好过对一个错误问题的准确答案”。
[1] NRC. Sustaining Marine Fisheries[M]. Washington DC: National Academy Press, 1999.
[2] Pauly D, Christensen V, Guénette S, et al. Towards sustainability in world fisheries[J]. Nature, 2002, 418(6898): 689-695.
[3] Sherman K. Sustaining the world′s large marine ecosystems[J]. ICES Journal of Marine Science: Journal du Conseil, 2015, 72(9): 2521-2531.
[4] FAO. The state of world fisheries and aquaculture 2014[R]. Rome: FAO, 2014.
[5] Baum J K, Myers R A, Kehler D G, et al. Collapse and conservation of shark populations in the Northwest Atlantic[J]. Science, 2003, 299(5605): 389-392.
[6] Mullon C, Fréon P, Cury P. The dynamics of collapse in world fisheries[J]. Fish and Fisheries, 2005, 6(2): 111-120.
[7] Hutchings J A. Collapse and recovery of marine fishes[J]. Nature, 2000, 406(6798): 882-885.
[8] Hutchings J A, Reynolds J D. Marine fish population collapses: consequences for recovery and extinction risk[J]. BioScience, 2004, 54(4): 297-309.
[9] Pinsky M L, Jensen O P, Ricard D, et al. Unexpected patterns of fisheries collapse in the world′s oceans[J]. Proceedings of the National Academy of Sciences of the Unites States of America, 2011, 108(20): 8317-8322.
[10] Jackson J B C, Kirby M X, Berger W H, et al. Historical overfishing and the recent collapse of coastal ecosystems[J]. Science, 2001, 293(5530): 629-637.
[11] Myers R A, Worm B. Rapid worldwide depletion of predatory fish communities[J]. Nature, 2003, 423(6937): 280-283.
[12] Venter O, Brodeur N N, Nemiroff L, et al. Threats to endangered species in Canada[J]. BioScience, 2006, 56(11): 903-910.
[13] Collie J, Hiddink J G, van Kooten T, et al. Indirect effects of bottom fishing on the productivity of marine fish[J]. Fish and Fisheries, 2016, 18(4):619-637.
[14] Purcell J E, Uye S I, Lo W T. Anthropogenic causes of jellyfish blooms and their direct consequences for humans: a review[J]. Marine Ecology Progress Series, 2007, 350: 153-174.
[15] Haag A L. Algae bloom again[J]. Nature, 2007, 447(7144): 520-521.
[16] Paerl H W, Huisman J. Blooms like it hot[J]. Science, 2008, 320(5872): 57-58.
[17] Cheung W W L, Watson R, Pauly D. Signature of ocean warming in global fisheries catch[J]. Nature, 2013, 497(7449): 365-368.
[18] Hoegh-Guldberg O, Bruno J F. The impact of climate change on the world′s marine ecosystems[J]. Science, 2010, 328(5985): 1523-1528.
[19] Walther G R, Post E, Convey P, et al. Ecological responses to recent climate change[J]. Nature, 2002, 416(6879): 389-395.
[20] Fabry V J, Seibel B A, Feely R A, et al. Impacts of ocean acidification on marine fauna and ecosystem processes[J]. ICES Journal of Marine Science, 2008, 65(3): 414-432.
[21] Jennings S, Kaiser M J. The effects of fishing on marine ecosystems[J]. Advances in Marine Biology, 1998, 34: 201-212, 212e, 213-352.
[22] Scheffer M, Carpenter S R. Catastrophic regime shifts in ecosystems: linking theory to observation[J]. Trends in Ecology & Evolution, 2003, 18(12): 648-656.
[23] Folke C, Carpenter S, Walker B, et al. Regime shifts, resilience, and biodiversity in ecosystem management[J]. Annual Review of Ecology, Evolution, and Systematics, 2004, 35(1): 557-581.
[24] Hilborn R, Walters C J. Quantitative Fisheries Stock Assessment: Choice, Dynamics and Uncertainty[M]. US: Springer, 1992.
[25] Hollowed A B, Bax N, Beamish R, et al. Are multispecies models an improvement on single-species models for measuring fishing impacts on marine ecosystems[J]. ICES Journal of Marine Science, 2000, 57(3): 707-719.
[26] Gårdmark A, Casini M, Huss M, et al. Regime shifts in exploited marine food webs: detecting mechanisms underlying alternative stable states using size-structured community dynamics theory[J]. Philosophical Transactions of the Royal Society B: Biological Sciences, 2014, 370(1659): 20130262.
[27] Bakun A, Weeks S J. Adverse feedback sequences in exploited marine systems: are deliberate interruptive actions warranted[J]. Fish and Fisheries, 2006, 7(4): 316-333.
[28] Walters C, Christensen V, Martell S, et al. Possible ecosystem impacts of applying MSY policies from single-species assessment[J]. ICES Journal of Marine Science, 2005, 62(3): 558-568.
[29] Mackinson S, Deas B, Beveridge D, et al. Mixed-fishery or ecosystem conundrum? Multispecies considerations inform thinking on long-term management of North Sea demersal stocks[J]. Canadian Journal of Fisheries and Aquatic Sciences, 2009, 66(7): 1107-1129.
[30] Fogarty M J. The art of ecosystem-based fishery management[J]. Canadian Journal of Fisheries and Aquatic Sciences, 2014, 71(3): 479-490.
[31] Pikitch E K, Santora C, Babcock E A, et al. Ecosystem-based fishery management[J]. Science, 2004, 305(5682): 346-347.
[32] Pitcher T J, Kalikoski D, Short K, et al. An evaluation of progress in implementing ecosystem-based management of fisheries in 33 countries[J]. Marine Policy, 2009, 33(2): 223-232.
[33] Hilborn R. Future directions in ecosystem based fisheries management: a personal perspective[J]. Fisheries Research, 2011, 108(2/3): 235-239.
[34] Hunsicker M E, Ciannelli L, Bailey K M, et al. Functional responses and scaling in predator-prey interactions of marine fishes: contemporary issues and emerging concepts[J]. Ecology Letters, 2011, 14(12): 1288-1299.
[35] Mace P M. A new role for MSY in single-species and ecosystem approaches to fisheries stock assessment and management[J]. Fish and Fisheries, 2001, 2(1): 2-32.
[36] Hall S J, Mainprize B. Towards ecosystem-based fisheries management[J]. Fish and Fisheries, 2004, 5(1): 1-20.
[37] Smith A D M, Fulton E J, Hobday A J, et al. Scientific tools to support the practical implementation of ecosystem-based fisheries management[J]. ICES Journal of Marine Science, 2007, 64(4): 633-639.
[38] Plagányi E E. Models for an ecosystem approach to fisheries[R]. FAO Fisheries Technical Paper No. 477. Rome: FAO, 2007.
[39] Levin S A. Ecosystems and the biosphere as complex adaptive systems[J]. Ecosystems, 1998, 1(5): 431-436.
[40] Walters C, Kitchell J F. Cultivation/depensation effects on juvenile survival and recruitment: implications for the theory of fishing[J]. Canadian Journal of Fisheries and Aquatic Sciences, 2001, 58(1): 39-50.
[41] Maunder M N, Watters G M. A general framework for integrating environmental time series into stock assessment models: model description, simulation testing, and example[J]. Fishery Bulletin, 2003, 101(1): 89-99.
[42] Moustahfid H, Link J S, Overholtz W J, et al. The advantage of explicitly incorporating predation mortality into age-structured stock assessment models: an application for Atlantic mackerel[J]. ICES Journal of Marine Science, 2009, 66(3): 445-454.
[43] Kerr S R, Ryder R A. Current approaches to multispecies analyses of marine fisheries[J]. Canadian Journal of Fisheries and Aquatic Sciences, 1989, 46(3): 528-534.
[44] Whipple S J, Link J S, Garrison L P, et al. Models of predation and fishing mortality in aquatic ecosystems[J]. Fish and Fisheries, 2000, 1(1): 22-40.
[45] Bolker B M. Ecological Models and Data in R[M]. Princeton, NJ: Princeton University Press, 2008.
[46] Dambacher J M, Gaughan D J, Rochet M J, et al. Qualitative modelling and indicators of exploited ecosystems[J]. Fish and Fisheries, 2009, 10(3): 305-322.
[47] Dambacher J M, Rothlisberg P C, Loneragan N R. Qualitative mathematical models to support ecosystem-based management of Australia′s Northern Prawn Fishery[J]. Ecological Applications, 2015, 25(1): 278-298.
[48] Takens F. Detecting strange attractors in turbulence[M]//Rand D, Young L S. Dynamical Systems and Turbulence, Warwick 1980. Berlin, Heidelberg: Springer, 1981: 366-381.
[49] Sugihara G, May R, Ye Hao, et al. Detecting causality in complex ecosystems[J]. Science, 2012, 338(6106): 496-500.
[50] Ye Hao, Beamish R J, Glaser S M, et al. Equation-free mechanistic ecosystem forecasting using empirical dynamic modeling[J]. Proceedings of the National Academy of Sciences of the United States of America, 2015, 112(13): E1569-E1576.
[51] Glaser S M, Fogarty M J, Liu Hui, et al. Complex dynamics may limit prediction in marine fisheries[J]. Fish and Fisheries, 2014, 15(4): 616-633.
[52] May R M, Beddington J R, Clark C W, et al. Management of multispecies fisheries[J]. Science, 1979, 205(4403): 267-277.
[53] Pimm S L, Rice J C. The dynamics of multispecies, multi-life-stage models of aquatic food webs[J]. Theoretical Population Biology, 1987, 32(3): 303-325.
[54] Sparre P. Introduction to multispecies virtual population analysis[J]. ICES Marine Science Symposia, 1991, 193: 12-21.
[55] Pope J G. The effect of biological interaction on the theory of mixed fisheries[R]. International Commission for the Northwest Atlantic Fisheries Selected Papers No. 1. International Commission for the Northwest Atlantic Fisheries, 1976: 157-162.
[56] Brown B E, Brennan J A, Grosslein M D, et al. The effect of fishing on the marine finfish biomass in the Northwest Atlantic from the Gulf of Maine to Cape Hatteras[R]. International Commission for the Northwest Atlantic Fisheries Research Bulletin No. 12. International Commission for the Northwest Atlantic Fisheries, 1976: 49-68.
[57] Murawski S A. Mixed-species yield-per-recruitment analyses accounting for technological interactions[J]. Canadian Journal of Fisheries and Aquatic Sciences, 1984, 41(6): 897-916.
[58] Jurado-Molina J, Livingston P A, Ianelli J N. Incorporating predation interactions in a statistical catch-at-age model for a predator-prey system in the eastern Bering Sea[J]. Canadian Journal of Fisheries and Aquatic Sciences, 2005, 62(8): 1865-1873.
[59] Jennings S, Kaiser M J, Reynolds J J D. Marine Fisheries Ecology[M]. London: Wiley-Blackwell, 2001.
[60] Bogstad B, Hauge K H, Ulltang Ø. MULTSPEC-A multi-species model for fish and marine mammals in the Barents sea[J]. Journal of Northwest Atlantic Fishery Science, 1997, 22: 317-341.
[61] Stefansson G, Palsson O K. Points of view: a framework for multispecies modelling of Arcto-boreal systems[J]. Reviews in Fish Biology and Fisheries, 1998, 8(1): 101-104.
[62] Begley J, Howell D. An overview of Gadget, the globally applicable area-disaggregated general ecosystem toolbox[R]. ICES CM 2004/FF: 13. 2004.
[63] Legendre P, Legendre L. Numerical Ecology[M]. 2nd ed. Amsterdam, The Netherlands: Elsevier, 1998.
[64] Pollock L J, Tingley R, Morris W K, et al. Understanding co-occurrence by modelling species simultaneously with a Joint Species Distribution Model (JSDM)[J]. Methods in Ecology and Evolution, 2014, 5(5): 397-406.
[65] Warton D I, Blanchet F G, O′Hara R B, et al. So many variables: joint modeling in community ecology[J]. Trends in Ecology & Evolution, 2015, 30(12): 766-779.
[66] Sheldon R W, Prakash A, Sutcliffe W H Jr. The size distribution of particles in the ocean[J]. Limnology and Oceanography, 1972, 17(3): 327-340.
[67] Blanchard J L. Body size and ecosystem dynamics: an introduction[J]. Oikos, 2011, 120(4): 481-482.
[68] Moloney C L, Field J G. Use of particle-size data to predict potential pelagic-fish yields of some southern African areas[J]. South African Journal of Marine Science, 1985, 3(1): 119-128.
[69] Andersen K H, Jacobsen N S, Farnsworth K D. The theoretical foundations for size spectrum models of fish communities[J]. Canadian Journal of Fisheries and Aquatic Sciences, 2016, 73(4): 575-588.
[70] Scott F, Blanchard J L, Andersen K H. Multispecies, trait-based and community size spectrum ecological modelling in R (mizer)[R]. 2014.
[71] Hall S J, Collie J S, Duplisea D E, et al. A length-based multispecies model for evaluating community responses to fishing[J]. Canadian Journal of Fisheries and Aquatic Sciences, 2006, 63(6): 1344-1359.
[72] Blanchard J L, Andersen K H, Scott F, et al. Evaluating targets and trade-offs among fisheries and conservation objectives using a multispecies size spectrum model[J]. Journal of Applied Ecology, 2014, 51(3): 612-622.
[73] Maury O, Poggiale J C. From individuals to populations to communities: a dynamic energy budget model of marine ecosystem size-spectrum including life history diversity[J]. Journal of Theoretical Biology, 2013, 324: 52-71.
[75] Stillman R A, Railsback S F, Giske J, et al. Making predictions in a changing world: the benefits of individual-based ecology[J]. BioScience, 2015, 65(2): 140-150.
[76] Grimm V, Revilla E, Berger U, et al. Pattern-oriented modeling of agent-based complex systems: lessons from ecology[J]. Science, 2005, 310(5750): 987-991.
[77] Shin Y J, Cury P. Using an individual-based model of fish assemblages to study the response of size spectra to changes in fishing[J]. Canadian Journal of Fisheries and Aquatic Sciences, 2004, 61(3): 414-431.
[78] Travers M, Shin Y J, Shannon L, et al. Simulating and testing the sensitivity of ecosystem-based indicators to fishing in the southern Benguela ecosystem[J]. Canadian Journal of Fisheries and Aquatic Sciences, 2006, 63(4): 943-956.
[79] Yemane D, Shin Y J, Field J G. Exploring the effect of marine protected areas on the dynamics of fish communities in the southern Benguela: an individual-based modelling approach[J]. ICES Journal of Marine Science, 2009, 66(2): 378-387.
[80] Grüss A, Harford W J, Schirripa M J, et al. Management strategy evaluation using the individual-based, multispecies modeling approach OSMOSE[J]. Ecological Modelling, 2016, 340: 86-105.
[81] Gray R, Fulton E A, Little R, et al. Ecosystem model specification with an agent based framework[R]. North West Shelf Joint Environmental Management Study Technical Report No. 16. CSIRO, 2006.
[82] Fulton E A. Approaches to end-to-end ecosystem models[J]. Journal of Marine Systems, 2010, 81(1/2): 171-183.
[83] Travers M, Shin Y J, Jennings S, et al. Towards end-to-end models for investigating the effects of climate and fishing in marine ecosystems[J]. Progress in Oceanography, 2007, 75(4): 751-770.
[84] Polovina J J. Model of a coral reef ecosystem: Ⅰ. The ECOPATH model and its application to French Frigate Shoals[J]. Coral Reefs, 1984, 3(1): 1-11.
[85] Pauly D, Christensen V, Walters C. Ecopath, Ecosim, and Ecospace as tools for evaluating ecosystem impact of fisheries[J]. ICES Journal of Marine Science, 2000, 57(3): 697-706.
[86] Christensen V, Walters C J. Ecopath with Ecosim: methods, capabilities and limitations[J]. Ecological Modelling, 2004, 172(2/4): 109-139.
[87] Walters C, Christensen V, Pauly D. Structuring dynamic models of exploited ecosystems from trophic mass-balance assessments[J]. Reviews in Fish Biology and Fisheries, 1997, 7(2): 139-172.
[88] Heymans J J, Coll M, Link J S, et al. Best practice in Ecopath with Ecosim food-web models for ecosystem-based management[J]. Ecological Modelling, 2016, 331: 173-184.
[89] Baretta J W, Baretta-Bekker J G, Ruardij P. From EMS to ERSEM, towards generic coastal ecosystem models[J]. Aquabiol, 1996, 104: 197-209.
[90] Fulton E A, Smith A D, Johnson C R. Biogeochemical marine ecosystem models Ⅰ: IGBEM-a model of marine bay ecosystems[J]. Ecological Modelling, 2004, 174(3): 267-307.
[91] Fulton E, Smith A, Punt A. Which ecological indicators can robustly detect effects of fishing[J]. ICES Journal of Marine Science, 2005, 62(3): 540-551.
[92] Fulton E A, Link J S, Kaplan I C, et al. Lessons in modelling and management of marine ecosystems: the Atlantis experience[J]. Fish and Fisheries, 2011, 12(2): 171-188.
[93] Horbowy J. The dynamics of Baltic fish stocks on the basis of a multispecies stock-production model[J]. Canadian Journal of Fisheries and Aquatic Sciences, 1996, 53(9): 2115-2125.
[94] Gaichas S, Gamble R, Fogarty M, et al. Assembly rules for aggregate-species production models: simulations in support of management strategy evaluation[J]. Marine Ecology Progress Series, 2012, 459: 275-292.
[95] Tjelmeland S, Lindstrøm U. An ecosystem element added to the assessment of Norwegian spring-spawning herring: implementing predation by minke whales[J]. ICES Journal of Marine Science, 2005, 62(2): 285-294.
[96] Hamre J, Hattlebakk E. System model (systmod) for the norwegian sea and the barents sea[M]//Rødseth T. Models for Multispecies Management. Heidelberg: Springer-Verlag, 1998: 93-115.
[97] Colomb A, Shin Y J, Gascuel D, et al. Using MOOVES, an individual-based model of the Guinean ecosystem, for assessing the response of Trophodynamic and size-based indicators to fishing pressure[C]//ICES CM 2004/FF. ICES, 2004: 14.
[98] Sekine M, Nakanishi H, Ukita M, et al. A shallow-sea ecological model using an object-oriented programming language[J]. Ecological Modelling, 1991, 57(3/4): 221-236.
[99] Finnoff D, Tschirhart J. Protecting an endangered species while harvesting its prey in a general equilibrium ecosystem model[J]. Land Economics, 2003, 79(2): 160-180.
[100] Bax N J. Application of multi- and univariate techniques of sensitivity analysis to SKEBUB, a biomass-based fisheries ecosystem model, parameterized to georges bank[J]. Ecological Modelling, 1985, 29(1/4): 353-382.
[101] Butterworth D S, Punt A E. Experiences in the evaluation and implementation of management procedures[J]. ICES Journal of Marine Science, 1999, 56(6): 985-998.
[102] Punt A E, Butterworth D S, de Moor C L, et al. Management strategy evaluation: best practices[J]. Fish and Fisheries, 2016, 17(2): 303-334.
[103] Bunnefeld N, Hoshino E, Milner-Gulland E J. Management strategy evaluation: a powerful tool for conservation[J]. Trends in Ecology & Evolution, 2011, 26(9): 441-447.
[104] Marasco R J, Goodman D, Grimes C B, et al. Ecosystem-based fisheries management: some practical suggestions[J]. Canadian Journal of Fisheries and Aquatic Sciences, 2007, 64(6): 928-939.
[105] Hockley N J, Jones J P G, Andriahajaina F B, et al. When should communities and conservationists monitor exploited resources[J]. Biodiversity & Conservation, 2005, 14(11): 2795-2806.
[106] Garcia S M, Charles A T. Fishery systems and linkages: implications for science and governance[J]. Ocean & Coastal Management, 2008, 51(7): 505-527.
[107] Brander K, Neuheimer A, Andersen K H, et al. Overconfidence in model projections[J]. ICES Journal of Marine Science, 2013, 70(6): 1065-1068.
[108] Silver N. The Signal and the Noise: Why So Many Predictions fail-but Some Don’T[M]. New York: Penguin Press, 2012.
[109] Walker W E, Harremo⊇s P, Rotmans J, et al. Defining uncertainty: a conceptual basis for uncertainty management in model-based decision support[J]. Integrated Assessment, 2003, 4(1): 5-17.
[110] Hill S L, Watters G M, Punt A E, et al. Model uncertainty in the ecosystem approach to fisheries[J]. Fish and Fisheries, 2007, 8(4): 315-336.
[111] Schindler D E, Hilborn R. Prediction, precaution, and policy under global change[J]. Science, 2015, 347(6225): 953-954.
[112] Dickey-Collas M, Payne M R, Trenkel V M, et al. Hazard warning: model misuse ahead[J]. ICES Journal of Marine Science, 2014, 71(8): 2300-2306.
[113] Levins R. The strategy of model building in population biology[J]. American Scientist, 1966, 54(4): 421-431.
[114] Orzack S H. The philosophy of modelling or does the philosophy of biology have any use[J]. Philosophical Transactions of the Royal Society B: Biological Sciences, 2012, 367(1586): 170-180.
[115] Collie J S, Botsford L W, Hastings A, et al. Ecosystem models for fisheries management: finding the sweet spot[J]. Fish and Fisheries, 2016, 17(1): 101-125.
[116] Planque B. Projecting the future state of marine ecosystems, “la grande illusion”[J]. ICES Journal of Marine Science: Journal du Conseil, 2016, 73(2): 204-208.
[117] Harwood J, Stokes K. Coping with uncertainty in ecological advice: lessons from fisheries[J]. Trends in Ecology & Evolution, 2003, 18(12): 617-622.
[118] Plagányi É E, Punt A E, Hillary R, et al. Multispecies fisheries management and conservation: tactical applications using models of intermediate complexity[J]. Fish and Fisheries, 2014, 15(1): 1-22.
[119] Andersen K H, Brander K, Ravn-Jonsen L. Trade-offs between objectives for ecosystem management of fisheries[J]. Ecological Applications, 2015, 25(5): 1390-1396.
[120] Gabriel W L, Mace P M. A review of biological reference points in the context of the precautionary approach[C]//Proceedings of the 5th NMFS NSAW. NOAA, 1999.
[121] Cadrin S X, Pastoors M A. Precautionary harvest policies and the uncertainty paradox[J]. Fisheries Research, 2008, 94(3): 367-372.
[122] Townsend H M, Harvey C, Aydin K Y, et al. Report of the 3rd national ecosystem modeling workshop (NEMoW 3): mingling models for marine resource management-multiple model inference[R]. NOAA Technical Memorandum NMFS-F/SPO-149. U.S. Department of Commerce, 2014.
[123] Möllmann C, Folke C, Edwards M, et al. Marine regime shifts around the globe: theory, drivers and impacts[J]. Philosophical Transactions of the Royal Society B: Biological Sciences, 2014, 370(1659): 20130260.
[124] Grimm V, Berger U. Structural realism, emergence, and predictions in next-generation ecological modelling: synthesis from a special issue[J]. Ecological Modelling, 2016, 326: 177-187.
[125] DeAngelis D L, Gross L J. Individual-based Models and Approaches in Ecology: Populations, Communities and Ecosystems[M]. New York: Chapman & Hall, 1992.
[126] Kuparinen A, Merilä J. Detecting and managing fisheries-induced evolution[J]. Trends in Ecology & Evolution, 2007, 22(12): 652-659.
[127] 李典谟, 马祖飞. 展望数学生态学与生态模型的未来[J]. 生态学报, 2000, 20(6): 1083-1089.
Li Dianmo, Ma Zufei. Prospect of mathematical ecology and ecological modeling[J]. Acta Ecologica Sinica, 2000, 20(6): 1083-1089.
[128] 陈新军, 李曰嵩. 基于个体生态模型在渔业生态中应用研究进展[J]. 水产学报, 2012, 36(4): 629-640.
Chen Xinjun, Li Yuesong. Review of application of individual-based model to fishery ecosystem[J]. Journal of Fisheries of China, 2012, 36(4): 629-640.
[129] 李云凯. 东海大陆架渔业生态系统模型研究[D]. 上海: 华东师范大学, 2009.
Li Yunkai. Ecological modeling of the East China Sea shelf ecosystem[D]. Shanghai: East China Normal University, 2009.
[130] 林群. 黄渤海典型水域生态系统能量传递与功能研究[D]. 青岛: 中国海洋大学, 2012.
Lin Qun. Studies on the ecosystem energy transfer and function in the typical waters of Yellow Sea and Bohai Sea[D]. Qingdao: Ocean University of China, 2012.
[131] Patrick W S, Link J S. Myths that continue to impede progress in ecosystem-based fisheries management[J]. Fisheries, 2015, 40(4): 155-160.
[132] Tukey J W. The future of data analysis[J]. The Annals of Mathematical Statistics, 1962, 33(1): 1-67.
The application of ecological models in fisheries management
Zhang Chongliang1, Chen Yong1,2, Han Dongyan1,2, Xing Lei1, Ren Yiping1
(1.LaboratoryofFisheriesEcosystemMonitoringandAssessment,CollegeofFisheries,OceanUniversityofChina,Qingdao266003,China; 2.SchoolofMaineSciences,UniversityofMaine,Orono04469 ,USA)
The ecological effects of fisheries and other human activities are increasingly concerned with the better understanding of marine ecosystems. Consequently, the concept of “ecosystem-based fisheries management” (EBFM) have been widely accepted by a range of researchers and institutes, for which ecological models have contributed to the basis of scientific methods and useful tools. Comparing to the singe-species assessment models, ecological models are superior in handling various ecological processes and mechanisms, habitat and environmental changes, and the structure and functions of ecosystems. Ecological models diversify in modeling objectives, mechanistic processes, model structure and parameters, and this study classified prevalent models into three categories on the basis of their ecological levels and components: multispecies models, community structure models and ecosystem models. We elaborated the advantages and disadvantages for each category of ecological models separately. We briefly introduced their potential use in guiding fisheries management on the basis of management strategy evaluation (MSE), and the caveats in model selection, building and implementation. Furthermore, we discussed the future development of ecological modeling in reducing uncertainty and improve predictability.
EBFM; ecosystem effects; MSE; uncertainty; predictability
10.3969/j.issn.0253-4193.2017.10.001
S932
:A
:0253-4193(2017)10-0001-18
2017-01-23;
:2017-03-08。
中央高校基本科研业务费专项资金资助(201562030,201612002)。
张崇良(1986—),男,山东省诸城市人,副教授,从事海洋生态系统评估研究。E-mail: zhangclg@ouc.edu.cn
*通信作者:任一平,男,山东省青岛市人,教授,从事渔业资源和生态修复研究。E-mail:renyip@ouc.edu.cn
张崇良,陈勇,韩东燕,等. 生态模型在渔业管理中的应用[J].海洋学报,2017,39(10):1—18,
Zhang Chongliang,Chen Yong,Han Dongyan,et al. The application of ecological models in fisheries management[J]. Haiyang Xuebao,2017,39(10):1—18, doi:10.3969/j.issn.0253-4193.2017.10.001
猜你喜欢
英语世界(2023年10期)2023-11-17 09:18:18
保健医苑(2021年7期)2021-08-13 08:48:02
学生天地(2020年36期)2020-06-09 03:12:30
小学科学(学生版)(2020年5期)2020-05-25 07:11:32
世界农药(2019年4期)2019-12-30 06:25:06
科学大众(中学)(2019年3期)2019-05-17 10:04:30
汽车观察(2018年10期)2018-11-06 07:05:26
中国水产(2017年2期)2017-02-25 07:56:08
湖南畜牧兽医(2016年3期)2016-06-05 08:37:56
少儿科学周刊·少年版(2015年1期)2015-07-07 17:15:12
