
奉贤滨海湿地蟹类和蟹洞的分布特征及影响因子
2021-11-10 03:10李翔田慧敏吴靖颖陈思雨赵明明徐平陈雪初由文辉
华东师范大学学报(自然科学版) 2021年2期
李翔 田慧敏 吴靖颖 陈思雨 赵明明 徐平 陈雪初 由文辉

摘要:研究了奉賢滨海湿地3种生境类型中的蟹类、蟹洞分布特征以及影响因子,揭示了影响蟹类及其洞穴分布的主要影响因子,加深了对穴居蟹类生活习性的认识。结果显示:①天津厚蟹多度在高潮滩芦苇生境高于低潮滩互花米草生境、中潮滩芦苇-互花米草混合生境(p<0.01),褶痕相手蟹多度在生境间无显著差异(p>0.05);②高潮滩芦苇生境中,蟹洞密度显著高于低潮滩互花米草生境、中潮滩芦苇-互花米草混合生境(p<0.05),蟹洞开口直径显著低于低潮滩互花米草生境、中潮滩芦苇-互花米草混合生境(p<0.05);③蟹类多度与蟹洞密度分布不存在显著的线性关系(p>0.05),但天津厚蟹多度与蟹洞密度具有极显著的正相关性(p<0.01);④蟹类多度与植物地下生物量呈负相关关系(p<0.01);⑤蟹洞密度与植被盖度与密度之间呈负相关关系,与相对高程、含水率、电导率、总有机碳含量与总氮含量呈正相关关系,其中,相对高程是与蟹洞密度的相关性排序最高的生境因子。
关键词:滨海湿地;蟹类;蟹洞;分布特征;影响因子
中图分类号:Q958.1文献标志码:ADOI:10.3969/j.issn.1000-5641.2021.02.016
DistributioncharacteristicsandinfluencingfactorsofcrabsandcrabburrowsintheFengxiancoastalwetland
LIXiang1,2,TIANHuimin1,2,WUJingying1,2,CHENSiyu1,2,ZHAOMingming1,2,XUPing1,2,CHENXuechu1,2,YOUWenhui1,2
(1.SchoolofEcologicalandEnvironmentalSciences,EastChinaNormalUniversity,Shanghai200241,China;2.ShanghaiKeyLaboratoryforUrbanEcologicalProcessesandEco-Restoration,EastChinaNormalUniversity,Shanghai200241,China)
Abstract:ThisstudyinvestigatedthedistributioncharacteristicsandinfluencingfactorsofcrabsandcrabburrowsinFengxiancoastalwetlandtorevealthemaininfluencingfactorsincrabsandcrabburrowsdistributionanddeepentheunderstandingofcrabslivinghabits.Theresultsshowedthat:①TheabundanceofHelicetientsinesisinthehigh-marshPhragmitesaustralishabitatishigherthanthatinthemiddle-marshPhragmitesaustralis-Spartinaalternifloramixedhabitatandlow-marshSpartinaalterniflorahabitat(p<0.01)。However,thereisnosignificantdifferenceintheabundanceofSesarmaplicatebetweenhabitats(p>0.05)。②Thedensityofcrabburrowsinthehigh-marshPhragmiteshabitatissignificantlyhigherthanthatinthemiddle-marshPhragmites-Spartinamixedhabitatandthelow-marshSpartinahabitat(p<0.05),whiletheaverageopeningdiameterofcrabburrowsissignificantlylowerthanthatinthemiddle-marshPhragmites-Spartinamixedhabitatandthelow-marshSpartinahabitat(p<0.05)。③Thereisnosignificantlinearrelationshipbetweencrababundanceandthedensityofcrabburrows(p>0.05),whilethereisasignificantpositivecorrelationbetweenthedensityofcrabburrowsandtheabundanceofHelicetientsinesis(p<0.01)。④Crababundanceisnegativelycorrelatedwithplantundergroundbiomass(p<0.01)。⑤Thereisanegativecorrelationbetweenthedensityofcrabburrowsandvegetationcoverageandplantdensity.Therelativeelevation,watercontent,conductivity,totalorganiccarboncontent,andtotalnitrogencontentarepositivelycorrelatedwiththedensityofcrabburrows.Amongthesefactors,therelativeelevationisthehabitatfactorwiththehighestcorrelationwiththedensityofcrabburrows.
Keywords:coastalwetlands;crabs;crabburrows;distributioncharacteristics;impactfactors
0引言
蟹类在湿地生态系统中的地位广受重视,被相关研究学者称为生态系统工程师[1-4]。滨海湿地受潮汐冲刷影响,其环境往往沿潮滩高度呈现梯度变化[5],很大程度上影响着蟹类及其洞穴在潮滩的空间分布[6]。由于蟹类活动性强、多数营穴居生活,在数量统计上具有较大的难度,而蟹洞是蟹类在滨海湿地中最显著的特征性结构[2],其密度和大小等特征在以往研究中常被用作估算不同体形蟹类密度的有效参数[7-8],甚至可以反映蟹类的行为模式以及生活习性等问题[9-10]。奉贤滨海湿地位于杭州湾北岸中段,潮汐侵蚀作用较强[11],互花米草自2000年被引种至奉贤滩涂用以促淤,而后在该区域迅速扩张,约占植被总面积的70%[12],湿地内植被特征以及沉积环境的变化对于该区域蟹类、蟹洞的分布特征的影响尚不明确。
目前,蟹类洞穴分布研究主要通过观察法[13]和蟹洞浇塑法[2,14]开展。由于蟹洞研究存在无法反映蟹类物种信息的缺点,所以本文采用陷阱捕获法[2,15-16]同步开展蟹类分布的研究,以探索奉贤滨海湿地蟹类地表活动与掘穴活动的分布特征。部分研究表明,蟹类与蟹洞的空间分布受非生物因子的影响更大[6,17]。Li[17]在黄河三角洲研究发现水深、含水量、盐度、土壤硬度、有机质和总碳等非生物因子对蟹类洞穴分布影响较大,蟹类偏好环境更柔软湿润的栖息地。Pombo等[6]则认为更细致的沉积物颗粒、更陡的斜坡将更适合沙蟹种群的分布,风暴潮将显著影响蟹类洞穴的分布,使蟹類向高潮滩聚集。也有研究认为植被特征是蟹类洞穴及蟹类分布更重要的因子[14,16]。高雪芹等[14]发现蟹类洞穴密度与凋落物的单宁含量及蛋白质含量、根系结构特点、植被盖度均显著相关。孟寒玉[16]发现红树林和互花米草群落能通过地上植被的遮阴作用,改变了、降低了沉积物表面温度,为植食性蟹类提供了充足的食物,成为褶痕相手蟹偏好的生境。本文将在上述的研究基础上,通过奉贤滨海湿地蟹类、蟹洞分布特征以及影响因子研究,揭示影响蟹类及其洞穴分布的主要影响因子,加深对穴居蟹类生活习性的认识,为湿地生境修复提供一定的指导。
1研究区域与方法
1.1研究区域概况
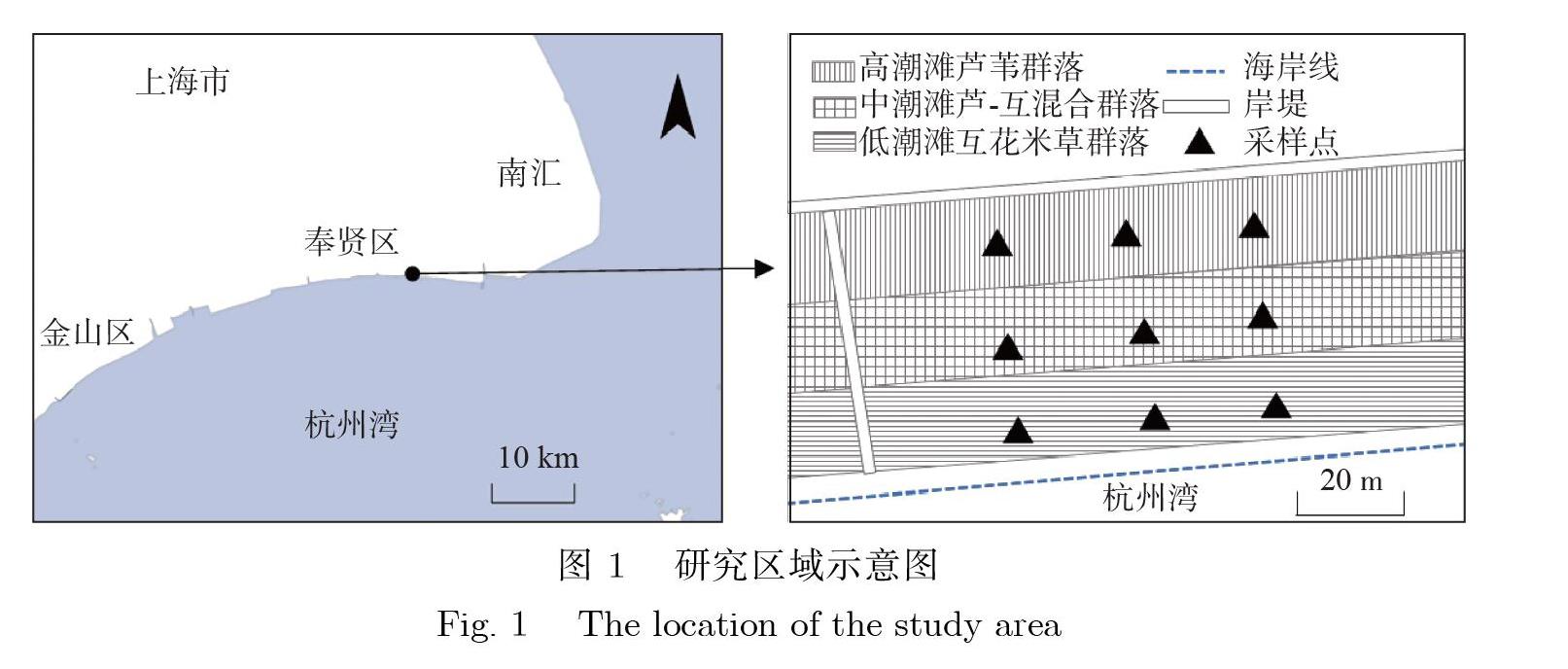
奉贤滨海湿地位于杭州湾北岸中段,属亚热带季风气候,1月均温6.7℃,7月均温30.7℃,全年日照1904.1h,年平均降水量1462.3mm[18]。研究区域位于东经121.65009°~121.65073°,北纬30.84210°~30.84273°,该岸段为非正规半日潮型,每月各有2次大小潮。20世纪90年代末以来,奉贤滨海湿地岸段沉积环境总体上以冲蚀为主,并有加剧的趋势[11,19]。湿地植被主要由芦苇(Phragmitesaustralis)与互花米草(Spartinaalterniflora)组成,沿着高程在潮间带呈带状分布,如图1所示。奉贤滨海湿地植被覆盖区蟹类组成以相手蟹科(Sesarmidae)的褶痕相手蟹(Sesarmaplicate)和方蟹科(Grapsidae)的天津厚蟹(Helicetientsinesis)为优势种[20]。
1.2样方设置
2019年9月,根据植被类型以及潮滩高度,奉贤滨海湿地内共被划分为3种生境类型,分别为低潮滩互花米草群落(简称互花米草生境)、中潮滩芦苇-互花米草混合群落(简称混合生境)以及高潮滩芦苇群落(简称芦苇生境),每种生境类型各设置1个40m×10m的条带状样地,在样地内随机设置3个1m×1m的蟹洞调查样方,如图1所示。样方调查与采样工作于9月20—22日9:00~11:00开展,调查期间天气晴朗,地温为23.5~30.0℃,沉积物盐度为11.5~18.0.
1.3蟹类分布调查以及蟹洞特征调查
在不同生境样带内各随机设置3个直径20cm、深度25cm的柱状陷阱[20],埋入地表后,使边缘与沉积物表面齐平,1d后收集所捕获的蟹类,识别计数后放回周围地表,实验重复两次,每个生境内蟹类的多度用单位努力捕获量(Catchperuniteffort,CPUE)来表示,单位为ind·trap–1·d–1[21],即通过在特定单位时间内使用陷阱捕获的蟹类数量来估计蟹类的总数量。在蟹洞调查样方内,使用游标卡尺测量所有蟹洞的洞口直径并记录蟹洞的数量。为了减少潮汐对蟹洞的掩埋影响,蟹洞调查选择在半日潮小潮的最低潮位前后2h期间开展。
1.4生境特征调查
由于蟹类生存于潮间带植被与沉积物中,并主要以植物和沉积物碎屑为食[22-23],在生境特征调查中选择了以植物特征以及沉积物特征代表蟹类生境。在9个样方内测量、记录并分析了4个植被特征与8个沉积物特征,包括植被盖度与密度、地上与地下生物量以及沉积物相对高程、温度、含水率、电导率、间隙水盐度、总有机碳(TOC)含量、总磷(TP)含量和总氮(TN)含量。
1.4.1植被特征
植被盖度与密度从蟹洞调查样方中,随机选取0.5m×0.5m的植被调查样方中通过目测计数获得;齐地收割植被调查样方表面所有活株擦净带回,使用烘箱105℃灭活15min,然后将温度调低至75℃,烘干至恒重后进行称重以获得地上生物量;地下生物量通过采集0.25m×0.25m面积内深度为20cm的地下部分,挑出石子碎渣、冲洗底泥以后,将植物根系进行烘干称重获得。
1.4.2沉积物特征
每个样方沉积物表面的相对高程使用U3测地型GNSS接收机测定[20]。由于蟹类多活动于0~5cm的沉积物表面[14,24],沉积物特征调查中仅原位测定或采样0~5cm深度的表层沉积物。沉积物温度、电导率均使用便携式土壤多功能测量仪(Procheck,Decagon,USA)原位测量获得。沉积物间隙水盐度由土壤间隙水采样管(Rhizonsoilmoisturesampler,Rhizon,Holland)原位抽取,再联合手持盐度计(MASTER-S/Milla,ATAGO,Japan)测量获得。含水率采用烘干法[23]。总有机碳、总氮、总磷指标均通过采样、除杂、风干、研磨后过100目土壤筛后通过以下实验室方法获得:总有机碳含量测定采用燃烧法,使用总有机碳分析仪(VarioTOC,Elementar,GERMAN)进行样品分析,测得的总碳即为总有机碳。总氮与总磷指标测定分别采用总凯氏氮法和总凯氏磷法,再联合化学分析仪(Smartchem200,ALLIANCE,FRENCH)测定。
1.5数据分析
数据分析采用统计软件包(IBMSPSSStatistics25)进行数据处理,采用单因素方差分析(OnewayANOVA)对于不同生境间的蟹类多度、蟹洞特征变化差异以及不同生境内植被和沉积物的特征差异进行比较分析,以p<0.05表示显著差异,p<0.01表示极显著差异,p>0.05表示无显著差异,再选用最小显著差法(LSD)进行均数间多重比较以开展后续结果分析。相关性分析中先使用Pearson相关系数对于蟹类多度与蟹洞分布以及生境因子之间的相关性分析,并进行双尾(Twotail)检验,再使用CANOCO5.0中的RDA分析对于相关性进行排序。
2结果
2.1蟹类分布特征
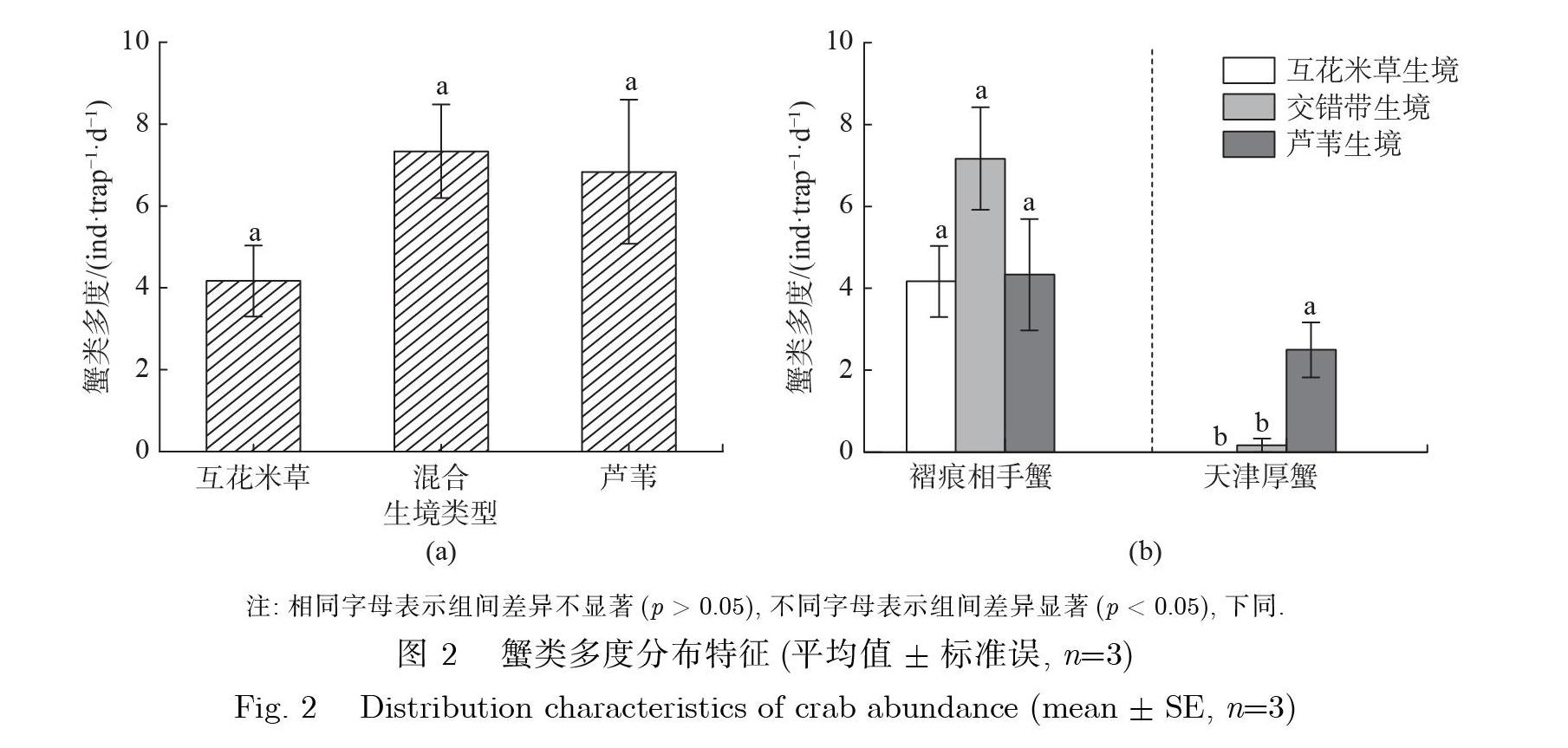
蟹类分布调查期间共捕获蟹类110只,属2科2种,分别为相手蟹科的褶痕相手蟹和方蟹科的天津厚蟹,分别占总捕获数的85.45%与14.55%。各生境间蟹类多度无显著差异(p>0.05)(见图2(a)),但不同种蟹类的分布特征表现不同(见图2(b)): 天津厚蟹在奉贤滨海湿地的多度分布表现为高潮滩芦苇生境极显著高于低潮滩互花米草与中潮滩混合生境(p<0.01),而褶痕相手蟹的分布则表现为广布于潮滩各个生境,蟹类多度在生境间无显著差异(p>0.05)。
2.2蟹洞分布特征
奉贤滨海湿地蟹洞密度在高潮滩芦苇生境内最高,达到(53.33±7.84)ind/m2,其次是在交错带生境,为(16.67±2.96)ind/m2,互花米草生境中蟹洞密度最低,为(7.00±1.53)ind/m2,最高值约为最低值的7.62倍。蟹洞密度在生境类型间存在极显著差异(p<0.01),高潮滩芦苇生境蟹洞密度显著高于低潮滩互花米草生境以及中潮滩混合生境(p<0.05),但低潮滩互花米草生境与中潮滩混合生境之间无显著差异(p>0.05),如图3所示。
在蟹洞开口直径调查中,互花米草生境内蟹洞平均开口直径最大,达到(15.38±2.47)mm,其次是交错带生境,为(14.03±1.35)mm,芦苇生境内蟹洞开口平均直径最小,为(10.72±0.69)mm。蟹洞开口直径在生境间存在显著差异(p<0.05),芦苇生境内蟹洞的开口直径显著低于低潮滩互花米草生境以及中潮滩混合生境(p<0.05),如图4所示。蟹洞开口直径与蟹洞密度之间存在显著的负相关关系(p<0.05),如图5所示。
蟹类总多度与蟹洞密度、蟹洞开口直径均无显著相关性(p>0.05),但在不同蟹类中存在不同的规律,褶痕相手蟹多度与蟹洞密度、蟹洞开口直径均无显著相关性(p>0.05),但天津厚蟹多度与蟹洞密度呈极显著的正相关关系(p<0.01),与蟹洞开口直径无显著相关关系(p>0.05),如表1所示。
2.3生境特征
植被盖度在生境间存在极显著差异(p<0.01),植被密度在生境间存在显著差异(p<0.05),植物地上生物量以及地下生物量在生境间均无显著差异(p>0.05)。高潮滩芦苇生境的植被盖度与植被密度均显著低于低潮滩互花米草生境与中潮滩混合生境(p<0.05),其余生境间植被特征无显著差异(p>0.05),如图6所示。
沉积物表面相对高程、温度和电导率在生境间存在极显著差异(p<0.01),含水率、总有机碳含量、总氮含量在生境间存在显著差异(p<0.05),间隙水盐度与总磷含量在生境间均无显著差异(p>0.05)。生境之间相对高程高潮滩芦苇生境>中潮滩混合生境>低潮滩互花米草生境(p<0.05)。在高潮滩芦苇生境中,沉积物温度、含水率、总有机碳含量以及总氮含量均显著高于其余生境(p<0.05),沉积物间隙水盐度、总磷含量分别显著低于、显著高于中潮滩混合生境(p<0.05),但与互花米草生境无显著差异(p>0.05)。互花米草生境电导率显著低于其余生境(p<0.05),其余生境間沉积物特征无显著差异(p>0.05),如图7所示。
2.4蟹类、蟹洞分布与生境因子间的相关性
蟹类、蟹洞分布与生境因子间的相关性结果显示(见表2),蟹类多度的分布与植物地下生物量存在负相关关系(p<0.01),与其他生境因子之间无显著相关关系(p>0.05)。蟹洞密度的分布与植被盖度、植被密度之间存在极显著的负相关关系(p<0.01),与相对高程、总有机碳含量之间存在极显著的正相关关系(p<0.01),与含水率、电导率以及总氮含量存在显著的正相关关系(p<0.05),与其他生境因子之间无显著相关关系(p>0.05)。由于各项生境因子之间存在着一些显著的相关关系(见表2),所以在生境因子与蟹类以及蟹洞分布关系的分析时需要谨慎、综合地考虑各项因子对于蟹类分布以及蟹洞分布的影响重要性。RDA排序结果显示(见图8): 蟹类分布与植物地下生物量具有较高负相关性;蟹洞密度与相对高程、含水率与总有机碳含量具有较高的正相关性(其中与相对高程相关性最高),与植被盖度、密度具有较高的负相关性。
3讨论
3.1不同生境内蟹类分布特征
以往研究发现不同植被类型[2,16]以及高程[5]的变化对于蟹类分布存在显著影响,并且不同蟹类在空间上的分布具有明显的差异。孟寒玉[16]曾在福建漳江口通过陷阱法监测红树林、互花米草丛和光滩3种生境中褶痕相手蟹与长足长方蟹(Metaplaxlongipes)分布的时空动态特征后,发现褶痕相手蟹更加偏好红树林和互花米草群落,而裸地则是长足长方蟹所偏好的生境类型。王金庆[2]在崇明东滩开展的蟹类分布研究发现植被类型对于天津厚蟹的分布无显著影响,但对无齿相手蟹存在显著影响,表现为互花米草生境>芦苇生境>裸地生境。本研究结果显示,奉贤滨海湿地中褶痕相手蟹的多度在生境间并无显著差异(p>0.05),但区域内天津厚蟹的多度在高潮滩芦苇生境高于低潮滩互花米草与中潮滩混合生境(p<0.01),该结论与骆蓓菁等[15]在奉贤潮滩开展的蟹类捕获实验结果一致,但与王金庆的结果存在差异,推测造成两地天津厚蟹分布结果差异的原因可能是样地的高程梯度不同。盛强[5]发现在崇明东滩不同高程下蟹类分布会随着高程的升高而增多,但本研究中总体的蟹类多度在各个潮滩高度间无显著差异(p>0.05),造成样地间蟹类分布特征差异的原因可能与蟹类的种类组成不同有关。
3.2不同生境内蟹洞分布特征
蟹洞密度在不同植被分布區之间常存在显著差异[14,25]。本研究结果显示蟹洞密度在芦苇生境中显著高于互花米草生境与交错带生境(p<0.05),该结果与王琰在崇明东滩的研究结果一致,但本文芦苇生境中的蟹洞密度要比该文章结果高出3~4倍,这可能是样地高程差异以及蟹种不同所致,有待进一步考证。洞穴密度高的高潮滩芦苇生境内蟹类洞穴平均开口直径较小,而在低洞穴密度的中、低潮滩生境内平均开口直径较大,二者呈显著的负相关关系,这与已有的研究结果一致[26-27]。由于蟹洞开口直径与蟹类甲壳宽存在显著相关性[14],使用开口直径推测蟹类大小可获得较为可靠的数据[7],可以推测在高潮滩芦苇生境中具有较多活跃的幼蟹个体,但由于幼蟹具有较小的活动范围以及较高的警惕性,难以用陷阱法对其数量进行准确量化,故该推测还待进一步野外观测与验证。
3.3蟹类、蟹洞分布特征与生境因子的相关性
本研究中蟹类总多度与蟹洞密度分布不存在显著的线性关系,因此,不能依据本区域的蟹洞密度准确估计蟹类的总多度[7],需要谨慎使用蟹洞去估计蟹类分布[6],其原因可能与蟹类的行为节律[28]有关,如招潮蟹属(Uca)会在涨潮前填堵洞口并经常更换住所,角眼沙蟹(Ocypodeceratophthalmus)经常在洞穴附近100~200m2的范围内活动后又回到自己的穴中[24]。但本研究还发现天津厚蟹多度与蟹洞密度具有极显著的正相关性,这可能是由于厚蟹属(Helice)动物偏爱在高潮滩建筑巢穴[29]有关。本文研究还发现蟹类分布与植物地下生物量呈负相关关系(p<0.01),即发达的地下根系会影响底栖动物的分布[30]。
已有研究发现蟹洞的分布受植被与沉积物特征的影响[2,13-14,17,26],但是不同研究的相关性分析结果受到样地类型以及不同蟹种的影响差异较大。在潮间带活动的蟹类通常会建造一个洞穴来躲避潮水和敌害[26],在植被覆盖率较低的区域,蟹类更易暴露在捕食者视野中,故蟹类会更多地依赖洞穴进行避敌,能较好地解释本文植被盖度、密度特征与蟹洞密度呈负相关的结果。另外,本研究发现蟹洞密度与沉积物特征中的相对高程、含水率、电导率、总有机碳含量与总氮含量呈正相关关系,其中,相对高程相关性排序最高,并且与含水率、电导率、总有机碳含量等其他显著相关的生境因子之间保持着与蟹洞密度一致的相关关系,可以初步推测本区域的蟹洞密度主要受高程的影响。奉贤滨海湿地潮汐侵蚀性强[12],有研究表明风暴潮将显著影响蟹类洞穴的分布,即蟹类会为了应对强潮冲击而向高潮滩聚集[6],而高潮滩生境内潮水的淹没频率较低、对蟹洞冲刷强度较弱,有利于蟹洞稳定存在。综上可得,奉贤滨海湿地互花米草入侵对于蟹洞分布有正向与负向两个方面的影响。首先,互花米草优异的促淤能力将促进潮滩沉积物的抬高,有利于蟹类获得更稳定的潮滩穴居环境,但随着互花米草向高潮滩的快速扩张[31],其过高的植被密度又将阻碍蟹类筑穴与活动[2],与此同时也会面临互花米草根系侵占蟹类生境的威胁,至于何种作用占主导,则还需要开展后续研究加以验证。
4结论
在奉贤滨海湿地3种不同生境类型中,褶痕相手蟹广布于潮滩各个生境,但天津厚蟹对高潮滩芦苇生境有显著偏好;高潮滩芦苇生境中蟹洞密度最高,平均蟹洞开口直径最小。蟹洞密度的分布受到植被特征与沉积物特征的综合影响,蟹洞密度与植物特征中的植被盖度与密度之间呈负相关关系,与沉积物特征中的相对高程、含水率、电导率、总有机碳含量与总氮含量呈正相关关系,其中,相对高程是与蟹洞密度的相关性排序最高的因子,即本区域蟹类偏好在安全、稳定的高潮滩上挖掘蟹洞。未来可在互花米草入侵的背景下开展不同时间尺度的蟹类、蟹洞分布研究,以全面反映其动态变化并判断其主导因子,更好地为奉贤滨海湿地生态环境保护和物种多样性保持提供科学指导。
[参考文献]
[1]KRISTENSENE.Mangrovecrabsasecosystemengineers;withemphasisonsedimentprocesses[J]。JournalofSeaResearch,2007,59(1): 30-43.
[2]王金庆。长江口盐沼优势蟹类的生境选择与生态系统工程师效应[D].上海:复旦大学,2008.
[3]GILP,FABRIZIOB,SAMWELL,etal.Arefiddlercrabspotentiallyusefulecosystemengineersinmangrovewastewaterwetlands?[J]。MarinePollutionBulletin,2009,58(11): 1694-1703.
[4]ALEXANDRIAM.WhatistheroleofecosystemengineersinnewEnglandsaltmarshes?Amesocosmstudyofthefiddlercrabandthepurplemarshcrab[J]。Wetlands,2019,3(2): 371-379.
[5]盛强。崇明东滩不同高程上蟹类对植物种间关系的影响[D].上海:复旦大学,2011.
[6]POMBOM,DEOLIVEIRAAL,XAVIERLY,etal.NaturaldriversofdistributionofghostcrabsOcypodequadrataandtheimplicationsofestimatesfromburrows[J]。MarineEcologyProgressSeries,2017,565:131-147.
[7]SCHLACHETA,LUCREZIS,PETERSONCH,etal.Estimatinganimalpopulationsandbodysizesfromburrows:Marineecologistshavetheirheadsburiedinthesand[J]。JournalofSeaResearch,2016,112:55-64.
[8]STELLING-WOODTP,CLARKGF,POOREAGB.Responsesofghostcrabstohabitatmodificationofurbansandybeaches[J]。MarineEnvironmentalResearch,2016,116:32-40.
[9]MACHADOGB,GUSM?O-JUNIORJB,COSTATM.BurrowmorphologyofUcauruguayensisandUcaleptodactylus(Decapoda:Ocypodidae)fromasubtropicalmangroveforestinthewesternAtlantic[J]。IntegrativeZoology,2013,8(3): 307-314.
[10]WANGM,GAOXQ,WANGWQ.DifferencesinburrowmorphologyofcrabsbetweenSpartinaalternifloramarshandmangrovehabitats[J]。EcologicalEngineering,2014,69(4): 213-219.
[11]赵建春,戴志军,李九发,等。强潮海湾近岸表层沉积物时空分布特征及水动力响应—以杭州湾北岸为例[J].沉积学报,2008,26(6): 1043-1051.
[12]黄华梅。上海滩涂盐沼植被的分布格局和时空动态研究[D].上海:华东师范大学,2009.
[13]王琰,童春富。长江口芦苇和互花米草盐沼湿地蟹类洞穴分布特征及主要影响因子[J].生态学报,2017,37(16): 5504-5513.
[14]高雪芹。互花米草群落与红树林群落蟹洞形态的比较研究[D].福建厦门:厦门大学,2011.
[15]骆蓓菁,李翔,罗鼎晖,等。天津厚蟹与褶痕相手蟹对杭州湾北岸典型盐沼植物的摄食偏好[J].华东师范大学学报(自然科学版),2019(6): 123-131.
[16]孟寒玉。互花米草入侵下漳江口紅树林区两种蟹类分布与生境选择研究[D].福建厦门:厦门大学,2017.
[17]LISZ,CUIBS,XIET,etal.Whatdrivesthedistributionofcrabburrowsindifferenthabitatsofintertidalsaltmarshes,YellowRiverDelta,China[J]。EcologicalIndicators,2018,92:99-106.
[18]上海奉贤区统计局。2019年奉贤统计年鉴(上)[EB/OL].(2019-09-06)[2020-05-29]。https://www.fengxian。gov.cn/tjj/tjsj/20190906/004002_218ac76d-7cb4-4c6e-9122-07480a7b67f5.htm.
[19]茅志昌,郭建强,虞志英,等。杭州湾北岸岸滩冲淤分析[J].海洋工程,2008,26(1): 112-117.
[20]骆蓓菁。杭州湾北岸优势蟹类对典型盐沼植物的摄食偏好及作用研究[D].上海:华东师范大学,2019.
[21]WALTONME,VAYLL,LEBATAJH,etal.Assessmentoftheeffectivenessofmangroverehabilitationusingexploitedandnonexploitedindicatorspecies[J]。BiologicalConservation,2007,138:180-188.
[22]KURODAM,WADAK,KAMADAM.Factorsinfluencingcoexistenceoftwobrachyurancrabs,HelicetfidensandParasesarmaplicatum,inanestuarinesaltmarsh[J]。JournalofCrustaceanBiology,2005(2): 146-153.
[23]余婕。河口潮滩湿地有机质来源、组成与食物链传递研究[D].上海:华东师范大学,2008.
[24]袁兴中,陆健健。潮滩微地貌元素-“生物结构”与小型底栖动物的空间分布[J].生态学杂志,2003,22(6): 124-126.
[25]鲁如坤。土壤农业化学分析方法[M].北京:中国农业科学技术出版社,2000.
[26]王琰。长江河口典型盐沼湿地蟹类洞穴的分布特征及其生态效应[D].上海:华东师范大学,2018.
[27]SCHUCHMANE,WARBURGMR.Dispersal,populationstructureandburrowshapeofOcypodecursor[J]。MarineBiology,1978,49(3): 255-263.
[28]戴爱云。蟹类的行为Ⅱ:群体行为[J].四川动物,1991,10(3): 16-19.
[29]戴爱云。蟹类的行为Ⅰ:个体行为[J].四川动物,1991,10(2): 19-22.
[30]杨泽华,童春富,陆健健。长江口湿地三个演替阶段大型底栖动物群落特征[J].动物学研究,2006(4): 411-418.
[31]王卿。互花米草在上海崇明东滩的入侵历史、分布现状和扩张趋势的预测[J].长江流域资源与环境,2011,20(6): 53-59.
(责任编辑:张晶)
猜你喜欢
安徽农学通报(2017年1期)2017-02-15
中国医药导报(2016年28期)2017-01-06
电脑知识与技术(2016年28期)2016-12-21
出版科学(2016年5期)2016-11-10
科技视界(2016年23期)2016-11-04
考试周刊(2016年50期)2016-07-12
读书(2016年5期)2016-05-21
赤峰学院学报·哲学社会科学版(2015年11期)2015-12-10
河北渔业(2015年9期)2015-09-15
河北渔业(2015年6期)2015-06-24
