
安吉毛竹林生态系统大小年水气通量变化特征
2017-08-01 00:06孙恒江洪龚莎莎陈晓峰
东北林业大学学报 2017年7期
孙恒 江洪 龚莎莎 陈晓峰
(浙江省森林生态系统碳循环与固碳减排重点实验室(浙江农林大学),临安,311300)(国际地球系统科学研究所(南京大学)) (浙江省森林生态系统碳循环与固碳减排重点实验室(浙江农林大学))
安吉毛竹林生态系统大小年水气通量变化特征
孙恒 江洪 龚莎莎 陈晓峰
(浙江省森林生态系统碳循环与固碳减排重点实验室(浙江农林大学),临安,311300)(国际地球系统科学研究所(南京大学)) (浙江省森林生态系统碳循环与固碳减排重点实验室(浙江农林大学))
利用涡度相关技术对安吉毛竹(Phyllostachysedulis)林生态系统进行观测,获得2014年(小年)和2015年(大年)毛竹林水气通量数据,并结合相关气象数据,对毛竹林大、小年水气通量变化特征进行对比分析。结果表明:毛竹林生态系统无论在大年还是小年水气通量值都为正值,月平均日变化和季节日变化都呈现单峰曲线;2015年8月份水气通量值最高(0.038 g·m-2·s-1),大于2014年7月份最高值(0.032 g·m-2·s-1),2014年水气通量值最低值出现在12月份,为0.009 6 g·m-2·s-1,2015年水气通量值最低值出现在1月份,为0.011 g·m-2·s-1;季节尺度上,具有明显的季节变化特征,水气通量值都是夏季最高,冬季最低,春秋季次之。大年毛竹春季受发笋抽叶的影响虽然大于秋季,但蒸散量却小于秋季;而小年春季降水量与蒸散量都小于秋季;大小年毛竹林各季节净辐射日变化都表现为单峰曲线,且水气通量与净辐射都表现出显著的相关性,但是受大小年差异的影响,大年春季的R2大于秋季R2,而小年则秋季R2大于春季R2。
毛竹林;水气通量;净辐射
Phyllostachysedulis; Water vapor flux; Net radiation
生态系统的水循环在植物生长发育和植被演替的生态学和生物化学过程中发挥着最为关键的作用[1-3]。作为水循环过程中的一个重要特征指标,水气通量(蒸散)是生态系统物质循环和能量流动的重要参与者。森林水气通量主要是指林下土壤表面蒸发通量,冠层截留水分蒸发通量和植被蒸腾通量三者之和,是反映森林植被水分状况的重要指标,是影响区域和全球气候变化的重要因子[4]。目前,在草地[5]、橡胶林[6]、红松林[7]等生态系统领域对水气通量都做了相关研究。而毛竹林作为我国重要的森林资源之一,是竹类中分布最广、面积最大、价值最高的竹种,在保持水土、维持环境碳平衡等方面作用明显[8]。毛竹具有明显的大小年,毛竹林的大小年反应竹林养分的制造、积累、分配、消耗不平衡的周期变化,使出笋、成竹年和换叶年交替进行[9],大年毛竹大量发笋长竹,具有其独特的爆发式生长,小年生鞭换叶,并且在小年的换叶期阶段会出现一个无叶期,使大约8%的太阳辐射不能被充分利用[10],姚希世等[9]对毛竹林大小年丰产差异的研究中认为,进入生理小年的毛竹由于竹叶转入生长衰退期而变黄,致使其群体光和效率降低,营养物质积累少,因此小年出笋、新成竹的数量和质量都不如大年。因而,毛竹大、小年的固碳能力、水分利用效率等方面可能会存在差异。毛竹作为重要的经济林种,其水分利用及消耗也是竹农们关心的问题,因此,探索毛竹林的水收支状况特别是大小年水气输送差异尤为重要。目前,国内对于毛竹的研究集中在光合生理[11]、能量分配[12]、生长动态[13]等方面。关于毛竹水气通量的研究较少,刘玉莉等[14]对单年毛竹林水气通量变化特征做了初步研究,舒海燕等[15]分析了连续3年毛竹的水气通量变化特征,但都没有对毛竹林大、小年的水气通量的差异进行分析探讨。
近些年,随着通量观测技术特别是涡度相关技术的发展,涡度相关法技术已经在陆地生态系统物质循环和能量流动中得到广泛应用,已成为国际通量观测网(FLUXNET)通量观测的标准方法[16-18]。利用涡度相关技术研究生态系统的水气或蒸散也受到各学者的关注[19-20]。本研究利用涡度相关观测技术对浙江省安吉县山川乡毛竹林生态系统进行观测,获得其水气通量数据,比较分析毛竹林大、小年的水气通量动态变化特征,探讨毛竹林生态系统在大、小年的水气通量变化特征差异及其与环境因子间相互关系,为毛竹林大、小年水收支状况的研究提供依据,更为当地毛竹林经营在大、小年的水分管理提供理论参考。
1 试验地概况
试验地位于浙江省安吉县山川乡(30°28′34.5″N,119°40′25.7″E),海拔高度380 m。该地区属于亚热带季风气候,四季分明,雨热同期,有典型的梅雨季节,空气比较湿润。年平均气温16.6 ℃,1月份最低,7月份最高,年降水量761~1 780 mm。当地竹林面积2 155 hm2,占该地区森林总面积的50.7%,其中毛竹林有1 693 hm2,占竹林总面积的78.6%,占森林总面积的39.8%。观测塔周围1 000 m范围内的植物类型以毛竹为主,且以粗放式经营为主。毛竹林林分密度为4 500株·hm-2,毛竹胸径12~18 cm,高度13~20 m,枝下高10~17 m,冠幅3.0 m×2.5 m,盖度90%,郁闭度0.9,毛竹节间短,壁厚,出笋期为3—5月份,且连年出笋能力较强,平均每年出笋产量1 000 kg·hm-2,林下有稀疏灌木和草本。毛竹有大小年之分,大年3—5月份为发笋期,5月底毛竹开始抽叶,6月份大量展开,7—9月份为主要生长季[21],小年出笋量少,在4—5月份为换叶期,至6月中上旬结束。
2 材料与方法
试验地建有高40 m的微气象观测塔,开路涡度相关系统的探头安装在距地面38 m高度上,由三维超声风速仪(CAST3,CampbellInc.,USA)和开路CO2/H2O分析仪(Li-7500,LiCorInc.,USA)组成,能量(风)的测定由三维超声波风速仪来完成,红外气体分析仪测量空气中的二氧化碳和水气浓度。利用数据采集器(CR1000,Campbell Inc.,USA)存储数据。气象因子观测由常规气象观测系统测得,该系统包括3层风速传感器(010C,metone,USA)和3层大气温度和湿度传感器(HMP45C,Vaisala,Helsinki,Finland),安装高度分别为1、7、38 m。2个SI-111红外温度仪分别置于2 m和23 m高处,用于采集地表和冠层温度;1个净辐射仪(CNR4,Kipp&Zonen)传感器安装高度为38 m,土壤温度传感器(109,Campbell,USA)、土壤含水量传感器(CS616,Campbell,USA),埋设深度分别为5、50、100 cm。常规气象观测系统数据通过数据采集器(CR1000,Campbell Inc.,USA)每30 min自动记录平均风速、气压、温度和净辐射等常规气象信息。此外,试验地内设有8个雨量筒用于记录信息,通量塔观测数据为实时观测数据。
水气通量(E)是利用实时测定的垂直风速与水气浓度的协方差求得。计算公式如下[22]:

式中:ρ表示干空气密度;q′表示比湿脉动;w′表示垂直风速;横线代表一段时间内的平均值;撇号表示脉动。水气通量的符号定义为,水气由生态系统进入大气,则水气通量值为正,反之为负[23]。
数据处理:毛竹的大年指毛竹换叶后当年孕笋,第二年春发笋成竹的期间,第二年发笋成竹后至第三年再换叶前称为毛竹的小年[24]。本试验采用2014年和2015年的水气通量数据,其中2014年正值当地毛竹林的小年,2015年为大年,所得水气通量数据为通量观测的30 min的平均值。数据处理方法主要采用2次坐标旋转来校正地形以及仪器安装不垂直对通量的影响,校正后的通量还进行了空气密度校正,即WPL校正,最后剔除由恶劣天气(有降水)、湍流不充分等导致的不合理数据,用平均日变化法(MDV)插补缺失的数据[25];>2 h的用其与净辐射的方程插补。利用Excel对所有数据进行整理,利用SPSS软件对数据进行相关性分析和多重比较,结果取平均值±标准差,用Excel作图,所有数据使用Excel软件进行筛选处理,用SPSS软件进行统计分析。
3 结果与分析
3.1 毛竹林大、小年水气通量月平均日变化和季节平均日变化特征
如图1、图2、图3所示,无论大年还是小年的毛竹林,水气通量各月平均日变化均呈现一定的规律性,各月最大值出现在11:00—14:00,夜间值较低;观察各月份水气通量值,从06:00—08:00开始逐渐升高至最高点后开始下降,在18:00—20:00处接近0,之后比较平缓。大年各月份水气通量日变化最大值为0.031~0.104 g·m-2·s-1,小年为0.022~0.084 g·m-2·s-1,2年最小值分别为-0.003~0.011 g·m-2·s-1和0.001~0.006 g·m-2·s-1,且出现在夜间。不同季节有明显差异,但基本集中在正午时分,夏季大小年变化规律相似,波动范围大,平均日变化最高值分别为0.083、0.069 g·m-2·s-1;冬季大小年日变化曲线最为平缓,起伏较小,平均日变化最大值分别为0.031、0.023 g·m-2·s-1;大年春秋两季的水气通量日变化趋势较一致,两曲线最高值波动于0.054~0.067 g·m-2·s-1,变化幅度较冬季大,显著小于夏季;小年春秋两季最高值波动范围较大年的波动范围大,为0.044~0.066 g·m-2·s-1。且小年水气通量月平均日变化值和季节日平均变化值都略小于大年。
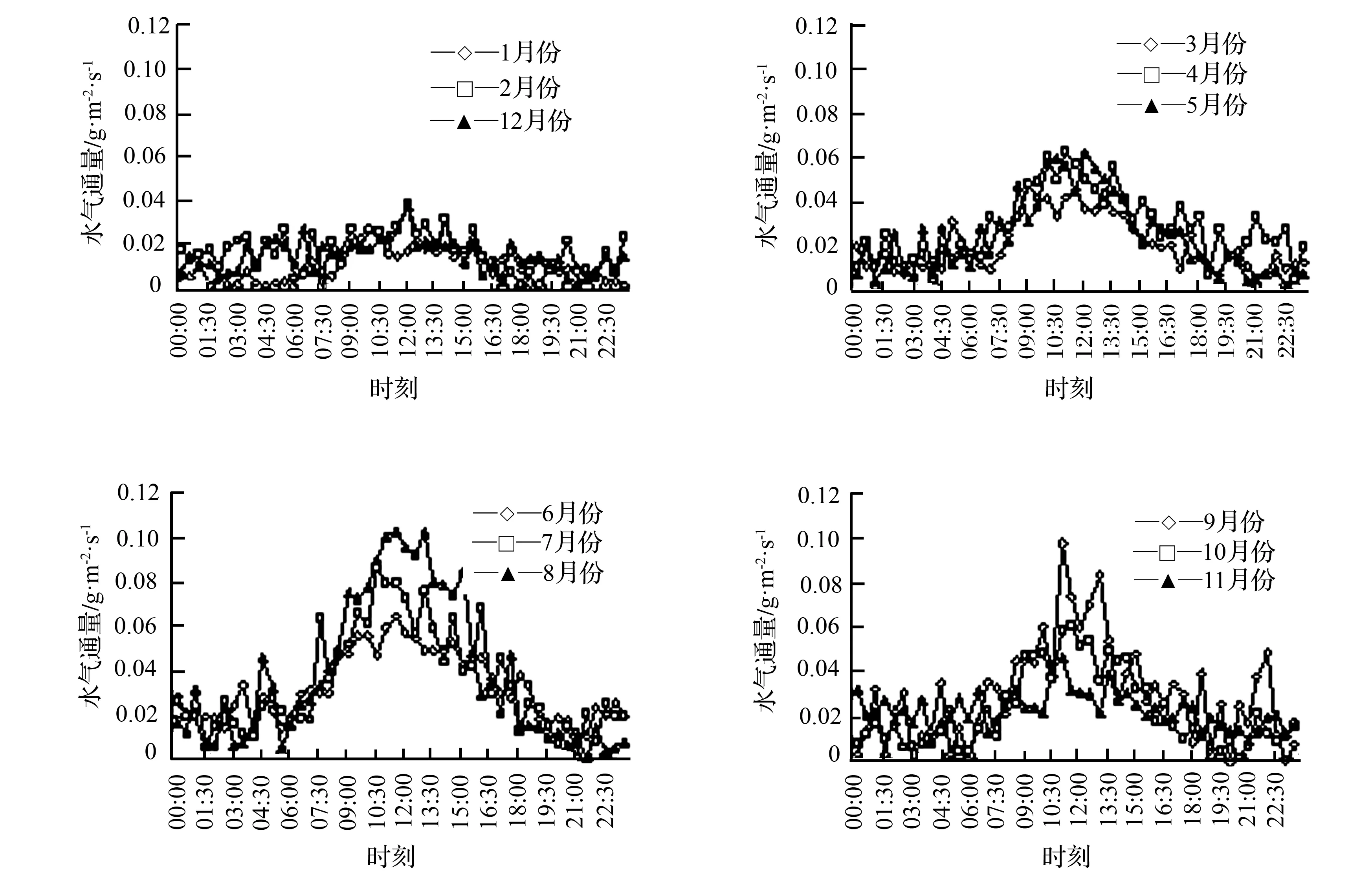
图1 毛竹林2015年(大年)水气通量各月份平均日变化分布
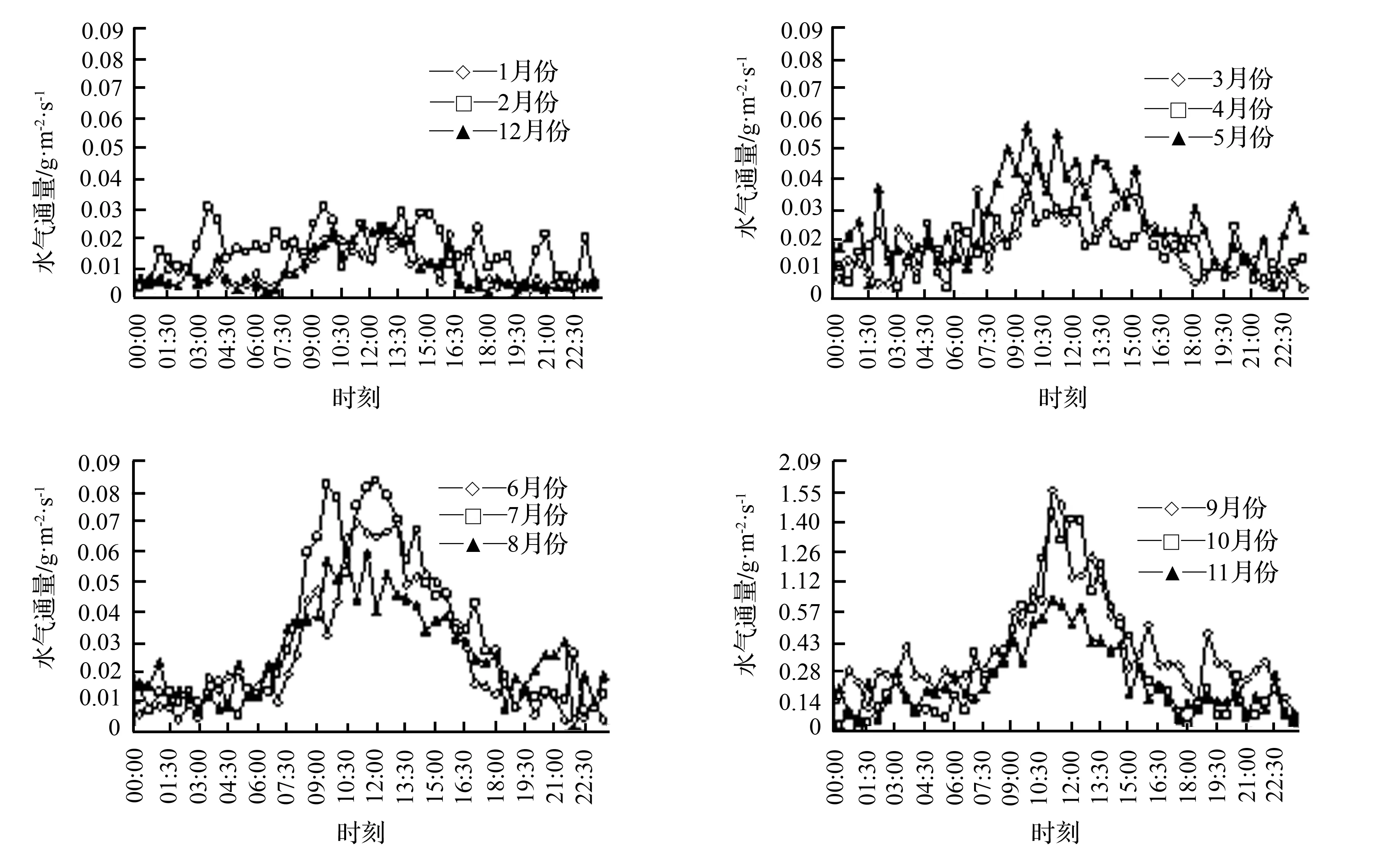
图2 毛竹林2014年水气通量各月份平均日变化分布

图3 2015、2014年毛竹水气通量各季节平均日变化分布
3.2 毛竹林大、小年水气通量月变化特征
如表1所示,2015年毛竹林各月水气通量值基本都略高于2014年的,2015年8月份水气通量值全年最高,为0.038 g·m-2·s-1,而2014年是7月份最高,最高值0.032 g·m-2·s-1,2014年水气通量值最低值出现在12月份,为0.009 6 g·m-2·s-1,2015年水气通量值最低值出现在1月份,为0.011 0 g·m-2·s-1。此外,两年水气通量值走势基本一致都出现一个峰值,但2015年4月份较2014年又出现一个小峰值,这可能与大小年有关,也可能与当月气候有关。

表1 2014、2015年毛竹水气通量月变化分布
3.3 蒸散量与降水量变化特征
蒸散是地表蒸发与植物蒸腾作用的总和,降落到地表的降水约有70%会通过蒸发与蒸散作用回到大气中[26],因此,蒸散作用是生态系统水循环的一个重要方式。年蒸散量是生态系统内地表蒸发和植被蒸腾的总耗水量,是全年水气通量的总和。
毛竹林2014年(表2)全年降水量1 455.30 mm,蒸散量654.47 mm,全年蒸散量占全年降水量的44.97%,夏季蒸散量和降水量最高,分别占全年蒸散量和降水量的34.46%和41.13%,显著高于其他各季的(p<0.05),春秋两季的蒸散量和降水量比较接近,差异不显著,蒸散量分别占25.04%和26.51%,降水量分别占21.84%和24.13%;冬季蒸散量与降水量最低,分别占13.99%和12.90%,春季降水量和蒸散量都小于秋季降水量和蒸散量。
2015年(表2)全年降水量1 723.00 mm,蒸散量801.59 mm,全年蒸散量占全年降水量的46.5%。各季度的降水量和蒸散量占全年降水量和蒸散量的比例悬殊差异不大,夏季蒸散量和降水量分别占全年蒸散量和降水量比例为35.68%和40.64%,显著高于其他各季的(p<0.05);春秋两季的蒸散量比较一致,约占25%,降水量分别占23.28%和19.14%;冬季蒸散量与降水量较低,分别占14.87%和16.94%,此外,春季降水量大于秋季,但其蒸散量却略小于秋季。

表2 2014、2015年毛竹林各季节蒸散量与降水量变化
注:表中数据为平均值±标准差;同列不同字母表示差异显著(p<0.05)。
3.4 各季节水气通量与净辐射的相关性
净辐射作为蒸散的驱动力,是白天水分蒸散的主要能量来源,因此,白天的蒸散量与净辐射是密切相关的[14]。从图4、图5可以明显看出,毛竹林水气通量对净辐射能做出不同的响应,呈正相关关系,且夏季水气通量对净辐射的响应最为明显,2014年由大到小的顺序为夏季(R2=0.890 1)、秋季(R2=0.817 6)、春季(R2=0.571 6)、冬季(R2=0.409 9),2015年由大到小的顺序为夏季(R2=0.812 2)、春季(R2=0.757 5)、秋季(R2=0.670 2)、冬季(R2=0.390 5),通过相关性检验(p<0.01),2年各季节水气通量与净辐射的相关性极显著,且2015年春季相关系数大于秋季相关系数,2014年与之相反。

图5 2015年毛竹林各季节水气通量与净辐射的响应拟合曲线
4 结论与讨论
大、小年毛竹林生态系统水气通量日变化特征比较相似,毛竹林水气通量都基本为正值,且水气通量日变化都表现为单峰曲线,至中午时分达到峰值,大年水气通量日变化最高值大于小年的,夜间水气通量值均较低,这是由于中午太阳辐射较强,温度高,湍流通量较大,导致植物叶片的蒸腾作用和地面水分的蒸发都较旺盛,而夜间温度低,植物蒸腾和地面蒸发较微弱,加之夜间净辐射值为负值,湍流通量很小,因而水气通量值较低。此研究结果与施婷婷等[20]对长白山阔叶红松林蒸散特征研究中生长季大于非生长季结果相同。季节日变化动态都表现为夏季变化规律性强,冬季最小,大年春秋两季的水气通量日变化趋势较一致,吻合度高,而小年春秋两季最高值波动范围较大年的波动范围大,这是因为2014年相比2015年夏季至秋季的温度降低幅度小,加上夏季高温多雨,空气湿度大而秋季气温刚转低,降水充足,较冬春两季加强了植被蒸腾作用和地面蒸发,进而导致水气通量的增大。月变化动态大年毛竹林各月水气通量值基本都略高于小年,只是2015年在4月份较2014年又出现一个小峰值。2015年4月份温度变化幅度、降水量及土壤5 cm含水量较2014年4月份大,这是因为在一定的空气温度范围内,伴随着温度的升高会使水分子获得的能量较大,运动加快[27],水易由液态变成气态,加上土壤含水量较大,能够满足蒸散的需要。龚莎莎等[28]对安吉2015年毛竹林净生态系统碳交换量研究中提到3月份出现过一次冻雨,受冷空气影响上旬气温偏低,多低温阴雨天气,整个生态系统光合能力比较弱,影响了CO2吸收及蒸腾作用。蒸腾是水气通量的重要组成部分,因此必会影响水气通量值。此外,5月份的温度、降水量与4月份相比都有所降低,4—5月份为毛竹林出笋旺期,需要足够的水分和养分供给,固5月份水气通量值低于4月份,因此,在4月份都出现一个小峰值,这主要与温度和降水有关。
2015年春季降水量大于秋季,土壤含水量比较充分,但其蒸散量却略小于秋季,这与张淑杰等[29]对辽宁省农田蒸散量研究中辽东地区降水量与蒸散量变化情况相似。而2014年则是春季降水量和蒸散量都小于秋季,这与橡胶林[6]生态系统旱雨季、太湖源雷竹林生态系统[30]降水量与蒸散量变化情况相似。这是因为毛竹林的水分需求基本上依赖自然降水,易受天气因素造成的水分胁迫的影响,春季的出笋及幼竹生长是毛竹的重要需水期[31],2015年毛竹林3月底开始发笋,5月底新竹抽叶,这期间老竹为保证幼竹的健康生长会加强对幼竹的水分传送[32],此时,新竹叶子还没有长出,至5月底新生竹刚开始抽叶光合作用能力较弱,其呼吸值较高,因此,虽然大年春季降水较大,但由于毛竹发笋和新竹生长,所需水分和养分较多,即使其温度、水分都比较适宜,但有限的土壤含水量不能完全满足于蒸散的需要,蒸散量减少。此外,由于夜间降水或露水对水气通量的观测有较大影响可能会低估通量值,加上数据在处理过程中使用的方法不同也可能会导致实际观测值降低,使蒸散量所占比例可能会略低于实际情况。
净辐射在不同季节变化不同,但不论是大年还是小年水气通量都能对其做出良好的响应。这是由于净辐射是植被与大气间物质和能量交换的主要驱动力,净辐射量在通常情况下是白天蒸发散的主要能量来源,因而,白天蒸发散随着净辐射的增加而增加[33]。本试验中2015年和2014年各季节毛竹净辐射日变化都表现为单峰曲线,最大净辐射值(分别为426.403、429.84 MJ·m-2·a-1)出现在12:00左右,夜间净辐射值小于零,为负值。这是由植物叶片对短、长波辐射的吸收特性造成的,日间,太阳短波辐射是构成净辐射的主要部分,夜间净辐射极小,叶片对长波辐射的吸收率大[34]。季节尺度上,大、小年水气通量值都与净辐射呈正相关,且夏季相关性最高,冬季相关性最低。这是因为夏季太阳辐射增强,而净辐射与太阳辐射高度呈正相关,叶片温度升高,使叶片水气扩散梯度增大,加上太阳辐射能够促使气孔导度增加,气孔开放,进而使蒸腾作用加强,导致夏季毛竹林水气通量对净辐射响应明显。此外,比较大、小年水气通量值与净辐射相关性系数(R2),大年春季相关系数大于秋季相关系数,这与牛晓栋等[35]对天目山常绿落叶阔叶混交林生态系统的研究结果相似;而小年确是春季相关系数小于秋季相关系数,与蔺恩杰等[30]等对雷竹林水气通量与净辐射关系的研究结果相似,这是因为小年在春季正值换叶期,并且会出现一段无叶期,净辐射会通过影响叶片蒸腾作用而影响水气通量,加上春季气候相对复杂,温度变幅较大,对水气通量有一定的影响,因此,其相关性小于秋季。
文中对安吉2014、2015年的水气通量及净辐射等数据进行分析,由于数据处理、校正、插补等因素,对于研究区域水气通量还存在一定的偶然性和局限性,也可能存在其他环境因子对水气通量造成影响,笔者将继续试验数据的观测和采集,以求研究有新的突破。
[1] 于贵瑞,高扬,王秋凤,等.陆地生态系统碳-氮-水循环的关键耦合过程及其生物调控机制探讨[J].中国生态农业学报,2013,21(1):1-13.
[2] 赵风华,于贵瑞.陆地生态系统碳-水耦合机制初探[J].地理科学进展,2008,27(1):32-38.
[3] 陈新芳,居为民,陈镜明,等.陆地生态系统碳水循环的相互作用及其模拟[J].生态学杂志,2009,28(8):1630-1639.
[4] 于贵瑞,孙晓敏.中国陆地生态系统通量观测的原理与方法[M].北京:高等教育出版社,2008:34-35.
[5] 常博,刘贤德,王顺利,等.祁连山不同坡向草地蒸散量及其影响因子的分析[J].中南林业科技大学学报,2014,34(4):90-95.
[6] 张晓娟,吴志祥,杨川,等.海南岛橡胶林生态系统水汽通量及其影响因子研究[J].热带作物学报,2015,36(8):1432-1439.
[7] 王秋凤,牛栋,于贵瑞,等.长白山森林生态系统CO2和水热通量的模拟研究[J].中国科学:D辑,2004(2):131-140.
[8] 蔡先锋,李洪吉,袁佳丽,等.临安毛竹快速生长期耗水特性及其影响因子[J].西北林业科学,2015,44(3):74-84.
[9] 姚希世,林建忠,林斌,等.毛竹丰产林大小年生长差异研究[J].湖南林业科技,2015,42(6):111-118.
[10] 张德值.大小年毛竹林改造试验[J].福建林业科技,2016,43(1):65-76.
[11] 张利阳,温国胜,张汝民,等.毛竹光合生理对变化的短期响应模拟[J].浙江林学院学报,2011,28(4):555-561.
[12] 洪伟,兰斌,吴承祯,等.毛竹林能量分配的研究[J].林业科学,1998,34(专刊1):78-81.
[13] 何东进,洪伟.毛竹生长动态模拟预测新模型[J].竹子研究汇刊,1998,17(3):32-36.
[14] 刘玉莉,江洪,周国模,等.安吉毛竹林水汽通量变化特征及其与环境因子的关系[J].生态学报,2014,34(17):4900-4909.
[15] 舒海蒋,江洪,陈晓峰,等.安吉毛竹林生态系统水汽通量的变化特征[J].生态学杂志,2016,35(5):1154-1161.
[16] 李琪,王云龙,胡正华,等.基于涡度相关法的中国草地生态系统碳通量研究进展[J].草业科学,2010,27(12):38-44.
[17] 薛红喜,李峰,李琪,等.基于涡度相关法的中国农田生态系统碳通量研究进展[J].南京信息工程大学学报(自然科学版),2012,4(3):226-232.
[18] 刘海军,黄冠华,JOSEF T,等.用涡度相关法测定网室内香蕉树蒸散量[J].农业工程学报,2008,24(9):1-5.
[19] 贾志军,张稳,黄耀,等.三江平原沼泽湿地垦殖对蒸散量的影响[J].环境科学,2010,31(4):834-841.
[20] 施婷婷,关德新,吴家兵,等.用涡动相关技术观测长白山阔叶红松林蒸散特征[J].北京林业大学学报,2006,28(6):1-8.
[21] 陈晓峰,江洪,牛晓栋,等.季节性高温和干旱对亚热带毛竹林碳通量的影响[J].应用生态学报,2016,27(2):335-344.
[22] 李菊,刘允芬,杨晓光,等.千烟洲人工林水汽通量特征及其与环境因素的关系[J].生态学报,2006,26(8):2449-2456.
[23] FALGE E, DENNIS B, RICHARD O, et al. Gap filling strategies for long term energy flux data sets[J]. Agricultural and Forest Meteorology,2001,107:71-77.
[24] 陈新安.毛竹林大小年生长规律探讨[J].中南林业调查规划,2010,29(1):21-23.
[25] 田义超,梁铭忠,胡宝清.2000—2013年北部湾海岸带蒸散量时空动态特征[J].农业机械学报,2015,46(8):146-158.
[26] 申双和,周英,李秉柏,等.冬小麦农田中净辐射的研究[J].植物生态学报,1999,23(2):171-176.
[27] SINGER K D, KUZYK M G, SOHN J E. Second-order nonlinear-optical processes in orientationally ordered materials: relationship between molecular and macroscopic properties[J]. Journal of the Optical Society of America B,1987,4(6):968-976.
[28] 龚莎莎,江洪,马锦丽,等.安吉毛竹林净生态系统碳交换量及叶绿素荧光参数的变化[J].浙江农业学报,2016,28(6):1003-1008.
[29] 张淑杰,班显秀,纪瑞鹏,等.农田蒸散量模型构建及蒸散状况分析[J].农业工程学报,2006,22(10):454-458.
[30] 蔺恩杰,江洪,陈云飞.太湖源雷竹林水汽通量变化及其对净辐射的响应[J].浙江农林大学学报,2013,30(3):313-18.
[31] 李迎春,杨清平,郭子武,等.毛竹林持续高温干旱灾害特征及影响因素分析[J].林业科学研究,2015,28(5):646-653.
[32] 袁佳丽,温国胜,张明如,等.毛竹快速生长期的水势变化特征[J].浙江农林大学学报,2015,32(5):722-728.
[33] 贾志军,宋长春.湿地生态系统CO2净交换、水汽通量及二者关系浅析[J].生态与农村环境学报,2006,22(2):75-79.
[34] 孙迪,关德新,袁凤辉,等.辽西农林复合系统中杨树水分耗散规律[J].北京林业大学学报,2010,32(4):114-120.
[35] 牛晓栋,江洪,方成圆,等.天目山常绿落叶阔叶混交林生态系统水汽通量特征[J].浙江农林大学学报,2016,33(2):216-224.
孙恒,女,1991年4月生,浙江省森林生态系统碳循环与固碳减排重点实验室(浙江农林大学),硕士研究生。E-mail:sunheng_@hotmail.com。
江洪,国际地球系统科学研究所(南京大学)、浙江省森林生态系统碳循环与固碳减排重点实验室(浙江农林大学) ,教授。E-mail:jianghong_china@hotmail.com。
2017年1月25日。
S715.4;S718.55
Variation Characteristics of Water Vapor Flux over AnjiPhyllostachysedulisForest Ecosystem in on-and-off Years//Sun Heng(Zhejiang Provincial Key Laboratory of Carbon Cycling in Forest Ecosystems and Carbon Sequestration, Zhejiang A&F University, Lin’an 311300, P. R. China); Jiang Hong(International Earth System Scientific Institute, Nanjing University); Gong Shasha, Chen Xiaofeng(Zhejiang Provincial Key Laboratory of Carbon Cycling in Forest Ecosystems and Carbon Sequestration, Zhejiang A&F University)//Journal of Northeast Forestry University,2017,45(7):22-27,39.
责任编辑:任 俐。
We compared the water vapor flux in on-and-off years ofPhyllostachysedulisusing the eddy covariance (EC) technique and related meteorological data in Anji Country of Zhejiang Province, China. The value of water vapor flux value ofP.edulisplantations was positive, and the diurnal variation of monthly and seasonal average showed a single peak curve in on-and-off years. The highest value (0.038 g·m-2·s-1) of on-year appeared in August, which was higher than July of off-year. The minimum value of the two year appeared in January. We observed obviously the water vapor flux for seasonal variation, and the highest value appeared in summer and lowest in winter. The rainfall of the spring was higher than the autumn in on-year, the evapotranspiration showed opposite trend. The rainfall and evapotranspiration in spring was higher than that in the autumn in off-year. The diurnal variation of net radiation in each season ofP.edulisplantations showed a single peak curve in on-and-off years. The water vapor flux was closely related to the net radiation. Due to the effect of on-and-off years, the determination coefficients (R2) of the spring was higher than that of the autumn in on-year, while that in off-year expressed opposite trend.
猜你喜欢
当代水产(2022年3期)2022-04-26
天津农林科技(2022年2期)2022-04-19
海洋石油(2021年3期)2021-11-05
非常规油气(2021年1期)2021-04-01
意林·少年版(2020年13期)2020-08-02
汉语世界(2020年3期)2020-06-19
东坡赤壁诗词(2020年2期)2020-06-04
乡村科技(2018年8期)2018-06-27
湖南林业科技(2017年1期)2017-02-06
现代检验医学杂志(2014年4期)2014-02-02
