
南方型美洲黑杨种质资源物候期特征1)
2017-08-01 00:06杨杉樊孝萍苏晓华黄国伟张新叶
东北林业大学学报 2017年7期
杨杉 樊孝萍 苏晓华 黄国伟 张新叶
(湖北省林业科学研究院,武汉,430075) (湖北省林业科学研究院石首杨树研究所) (中国林业科学研究院林业研究所) (湖北省林业科学研究院)
南方型美洲黑杨种质资源物候期特征1)
杨杉 樊孝萍 苏晓华 黄国伟 张新叶
(湖北省林业科学研究院,武汉,430075) (湖北省林业科学研究院石首杨树研究所) (中国林业科学研究院林业研究所) (湖北省林业科学研究院)
以湖北石首南方型美洲黑杨种质资源库中的443个黑杨种质为对象,对其进行物候观测分析。结果表明:不同美洲黑杨无性系物候之间存在较大差异,不同的物候期表现出不同的时序特征,变异最大的物候是叶变色始期,变异系数达到52.74%,其次是叶全部变色期和芽膨大始期。与变色落叶相关的各物候期之间相关性极显著,与萌芽相关的各物候间也显著相关;同时,第一主成分是与叶黄叶落有关的因子,第二主成分是与萌芽展叶有关的因子,第三主成分载荷较高的是落叶末期和全部落叶期,其中第一、第二、第三主成分累计贡献率达到87.29%。不同种源地的纬度与萌芽展叶物候期呈正相关,与叶变色落叶物候期呈明显的负相关。
南方型美洲黑杨;种质资源;物候特征
Southern type ofPopulusdeltoides; Germplasm; Phenological characteristic
林木种质资源是林木遗传多样性的载体、物种多样性的重要组成部分、开展林木育种的基础材料和国家重要的战略资源。美洲黑杨(Populusdeltoides)速生性能好、经济价值和遗传育种价值大,日益受到世界杨树育种学界和栽培工作者的普遍重视。我国最早引进的美洲黑杨栽培品种是山海关杨,1972年美洲黑杨I-69/55和I-63/51及欧美杨杂种I72/58无性系引种获得较大成功,在长江中下游和江淮平原地区创造了我国杨树速生丰产的新记录[1]。从上世纪80年代起,我国开始重视美洲黑杨种质资源的引进及保存利用研究,先后在山东长清、江苏泗洪、湖北石首等地建立美洲黑杨基因库[2-4]。这些基因库的建立消除了直接从国外引种优良无性系所带来的环境因素影响,为我国开展自己的黑杨派育种奠定了基础[5]。我国杨树育种工作者在全国不同区域开展了各种美洲黑杨与欧美杨、青杨及美洲黑杨间的杂交育种和遗传评价等相关研究[6-14],同时对美洲黑杨的遗传转化[15]、分子标记[16]及基因表达[17]等多方面进行了研究。
物候期观测分析是进行种质资源研究与利用的基础性工作,植物物候期作为重要参考资料,在农林业生产上,对苗木引种与育种工作具有很强的指导性[18]。因此,我国对引进的美洲黑杨资源开展了大量相关研究,但对其物候期研究相对较少,解荷锋等[19]对山东长清县美洲黑杨基因库中的9年生的49个无性系进行了物候观察分析;唐洁等[20]对60个美洲黑杨无性系苗期物候特征观测比较;杨成生等[21]对甘肃引种栽培的10个杨树品种只进行了苗期物候观测。这些观测结果对当地的杨树引种推广具有重要指导意义,其中针对美洲黑杨无性系虽然进行了物候观察,但数量有限,不能系统全面反映南方型美洲黑杨资源在长江中下游地区的物候特征。因此,本研究以湖北省林科院石首杨树研究所建立的南方型美洲黑杨种质资源库中的443个黑杨种质为对象,对其进行物候观测。初步了解443个黑杨种质的整体物候特征,为合理评价及正确利用这些种质提供翔实的基础资料,同时为南方型美洲黑杨种质资源保存、良种选育、林业创新资源共享和可持续经营奠定基础。
1 试验地概况
试验地位于湖北省石首市东升镇南碾垸,该地区位于江汉平原与洞庭湖平原结合部,土壤条件以潮土为主,pH值6.5~7.8,土壤颗粒均匀,质地疏松,理化性能良好,土壤有机质深厚,土壤有机质平均质量分数为1.39%~3.71%。该地区属典型的亚热带季风气候地区,具有阳光充足、热量丰富、无霜期长、降水充沛等特点。无霜期285 d,年平均气温16.4 ℃,年平均日照时间1 844.3 h,年平均降水量1 322.6 mm[4]。
2 材料与方法
试验材料:以湖北省林科院石首国家杨树良种基地美洲黑杨种质资源库中收集保存的448个南方型美洲黑杨无性系中的443个无性系为试验材料,2011年3月造林,株行距4 m×8 m[4]。
物候观测:2015年2月15日起,对资源库的443个无性系进行物候观测,每个无性系选择3株标准株,取其平均期作为原始记录值,定株观测8个物候指标特征,包括芽膨大始期(X1)、芽鳞脱落期(X2)、展叶始期(X3)、叶变色始期(X4)、叶全部变色期(X5)、落叶始期(X6)、落叶末期(X7)和全部落叶期(X8)。各个物候始期以出现明显的物候相为标志。
数据处理:以芽膨大始期、芽鳞脱落期、展叶始期这3个萌芽展叶相关物候期距2月15日的天数对物候期进行数据转换,以叶始变色期、叶全部变色期、落叶始期、落叶末期和全部落叶期这5个落叶相关物候期距9月1日的天数对物候期进行数据转换。根据转换后的定量数据,首先采用Excel软件进行数据初步统计分析,计算各物候期最大值、最小值、极差及变异系数等参数,分析不同物候期中参试的美洲黑杨种质的多样性特征,然后利用SPSS19.0软件进行各无性系物候期之间的主成分分析。
3 结果与分析
3.1 物候期差异
本研究对443个5年生南方型美洲黑杨无性系的物候期进行了观测,在此只随机列出部分无性系的物候相关信息(见表1、表2)。通过对443个无性系8个不同物候期定量数据的初步分析,结果表明各无性系物候之间存在较大差异。
由表1可知,变异最大的物候是叶变色始期(X4),变异系数达到52.74%,其次是叶全部变色期(X5)和芽膨大始期(X1)。从整体情况来看,萌芽展叶阶段的物候期变异较小,落叶阶段的物候期变异较大。落叶阶段中相关的5个不同物候期,极差跨度为45~64 d,而萌芽展叶阶段中3个不同物候期的极差跨度为21~24 d。仅从极差天数看,差异最大的是叶全部变色期(X5),最早和最晚时间相差64 d,差异最小的是展叶始期(X3),最早和最晚展叶的时间相差21 d。物候调查数据总体说明,萌芽展叶期不同无性系之间虽然物候有差异,但时间相对较一致,在2月底至3月下旬全部无性系基本都能完成萌芽及展叶。而落叶阶段中同一个物候期不同无性系间前后相差2个月左右,如叶变色始期(X4)最早是无性系TN03-N54,为当年9月1日,最晚的无性系是LA06-N58,其叶开始变色期为11月9日。
3.2不同物候期特征
由表1、表2、表3可知,在芽膨大开始期(X1),443个美洲黑杨无性系分成5个时序组,其中最早芽开始膨大的一组有16个无性系,于2月28日开始膨大,占总无性系的3.61%,最晚的一组有23个,于3月24日开始膨大,占5.19%,其余404个无性系分别于3月7日、3月12日和3月19日芽开始膨大,占91.20%,由此说明美洲黑杨无性系的芽开始膨大期相对较一致,主要集中在3月上旬和中旬。
在芽鳞脱落期(X2),443个美洲黑杨无性系分成7个时序组,其中有400个无性系(占总无性系90.29%)的芽鳞脱落主要集中在3月19日(120个)、3月24日(230个)和3月31日(50个)这3个时期,其他43个无性系的芽鳞脱落相对较分散,有3月7日、3月12日、3月22日和3月28日这4个时期。与芽膨大始期(X1)相比较,这两个物候期并没有表现出一致的规律性。分析X1和X2的组成特征,可以看出这两个物候期中,不同的无性系表现出不同的物候特性,不是芽膨大早就一定芽鳞脱落早,即不同的无性系其芽鳞在芽上留存的时期是不一样的。
在展叶始期(X3),443个不同的美洲黑杨无性系仅表现出4个不同时序组,即3月19日、3月24日、3月31日及4月9日,时间相对较集中在3月下旬。简单比较X1、X2和X3的组成特征,可以看出X3的多样度较X1和X2小,其中X2的多样度最丰富。

表1 部分南方型美洲黑杨无性系物候期观测表
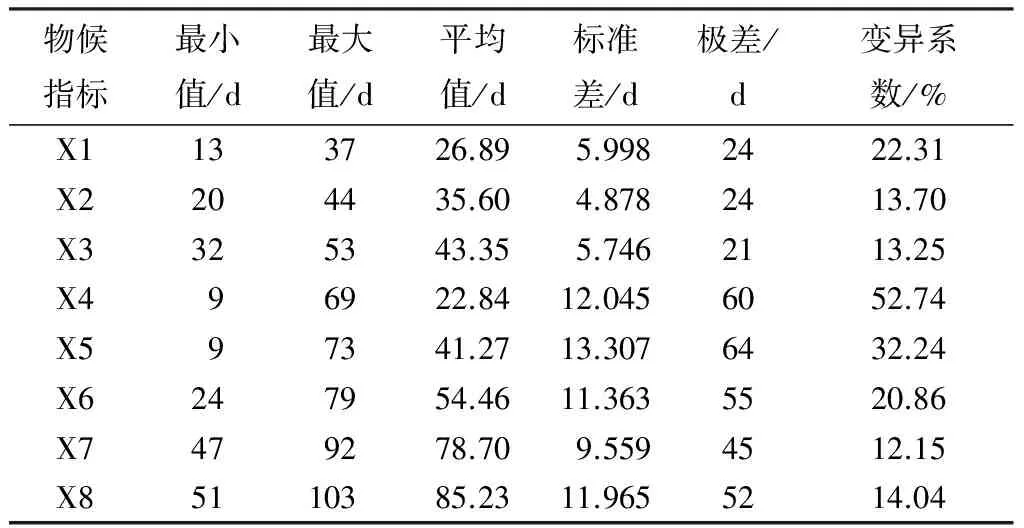
表2 南方型美洲黑杨无性系物候期观测信息
在叶变色始期(X4),443个无性系分布在11个不同时序组,最早变色的只有1个无性系TN03-N54,集中开始变色有两个时间,9月10日和9月25日,占总无性系的85.55%,说明在这两个时期出现了两次较大的气温变化。其余后期开始变色的55个无性系持续时间较长,最晚的1个无性系LA06-N58于11月9日才开始变色。该物候特征分析表明不同的无性系对气温变化的响应是不一样的,变色早的与变色晚的前后相差约70 d。
在叶全部变色期(X5),443个无性系相对集中叶片全部变色时间段为9月25日至10月27日,该时间段内全部变色的有405个无性系,占总种质的91.42%。比较X5和X4,两者总体变化规律似乎明显不同,但仔细比较可以发现,X4中第一次大的气温变化致使约120个种质在9月10日叶片开始变色,对应在X5中,9月25日时也出现了一次大的集中性叶片全部变色期,种质数量达113个,仔细比较X4与X5中相对应的具体无性系,结果表明是同一批无性系叶片在9月10日时开始变色,9月25日时全部变色。相类似,9月25日X4叶片开始变色的259个无性系,在X5中持续表现为10月13日至10月27日叶片全部变色的269个无性系。
在落叶始期(X6),443个无性系呈现出以10月27日为峰值的近正态分布特征,最早落叶的13个无性系于9月25日开始落叶,然后落叶无性系个数随时间推迟逐步增加,于10月27日达到最大值,随后又逐步降低,最迟的8个无性系于11月19日开始落叶,最早与最晚开始落叶的无性系前后相差55 d,表现出极大的遗传差异性。

表3 不同物候期特征
对于落叶末期(X7)和全部落叶期(X8),443个无性系表现出极其相似的规律和特征,不同观测期对应的无性系数量完全相同,只是一个时间的顺延,仅有1个例外,就是落叶末期中最后的79个无性系在全部落叶期时分为两类,一类是仅有1个无性系于12月9日全部落叶,另一类是其他78个无性系皆于12月13日全部落叶。
从8个不同物候期的特征分析可以看出,443个美洲黑杨无性系中物候多样性丰富,其中与落叶相关的几个物候期变异较大,其中部分无性系具有特殊性,如展叶早落叶晚类型、展叶晚落叶晚类型等,可作为特殊遗传种质材料进一步评价利用。
3.3 物候项目的主成分分析
从表4的相关矩阵及显著性分析结果中可以看出,在8个物候观测期项目中,X1与X2,X1与X3,X4与X5、X6、X7、X8,X5与X6、X7、X8,X6与X7、X8,X7与X8之间的相关达到极显著水平,同时,X2与X3之间显著相关,其他物候项目间相关不显著。值得注意的是芽始膨大期X1与叶全部变色期X5之间相关达到显著水平,虽然一个是春季的物候期,一个是秋冬季的物候期,但结果表明两者之间存在着一定的相关性,从表2的特征组成也可以看出两者存在一定的相关,即芽开始膨大较早的无性系叶全部变色的时期也较早,芽膨大较晚的无性系叶全部变色的时期也较晚。

表4 不同物候期相关矩阵及显著性
注:** 表示极显著相关(p<0.001),*表示显著相关(p<0.005)。
从表5可知,第一、第二、第三主成分累计贡献率达到了87.29%,即这3个因子可以反映出原变量87%左右的信息,因此,把原本应在8维空间考察的复杂问题可以简化为3维空间进行分析。

表5 主成分的特征值、贡献率、累计贡献率
从表6的因子载荷矩阵中可以看出,第一主成分载荷较高的都是与叶黄叶落有关的因子,如叶变色始期(X4)、叶全部变色期(X5)、落叶始期(X6)、落叶末期(X7)和全部落叶期(X8),其中全部变色期(X5)载荷量最大,在5个落叶相关物候期中具有代表性;第二主成分载荷较高的有芽膨大始期(X1)、芽鳞脱落期(X2)和展叶始期(X3),其中芽鳞脱落期(X2)载荷量最大,代表性最强;第三主成分载荷较高的有落叶末期(X7)和全部落叶期(X8),虽然第三主成分比第一和第二主成分的载荷量偏低较多,但其贡献率超过10%,其中全部落叶期(X8)作为一个重要物候特征,在第三主成分的2个落叶项目中具有代表性。
3.4 不同种源物候期与纬度的相关性
443份种质资源中,原产地较清楚的有198个无性系,分别为美国中南部的田纳西州(TN)105个无性系、密西西比州(MS)59个无性系、路易斯安娜州(LA)34个无性系。对这3个种源的所有无性系的不同物候期取平均值,然后与这3个州的平均纬度进行相关性分析(见表7)。由于田纳西州的经度范围几乎包含了密西西比州和路易斯安娜州,因此,未分析其经度与各物候的相关性。

表6 因子载荷矩阵
从表7中可以看出,种源地的纬度与芽膨大始期(X1)、芽鳞脱落期(X2)和展叶始期(X3)存在明显的正相关关系,而与叶变色始期(X4)、全部变色期(X5)、落叶始期(X6)、落叶末期(X7)和全部落叶期(X8)都与种源地的纬度存在明显的负相关关系,即随着纬度的增加,南方型美洲黑杨的萌芽展叶期都会相继推迟,路易斯安娜州萌芽展叶最早,其次是密西西比州,最后是田纳西州;而随着纬度的增加,叶黄叶落的时间会相继提前,田纳西州种源最先叶片变色开始落叶,其次是密西西比州种源,最后是路易斯安娜州种源。3个种源相比,路易斯安娜州种源是萌芽展叶最早,变色落叶最晚,生长期最长的种源,相反,田纳西州种源是萌芽展叶最晚,变色落叶最早,生长期最短的种源。
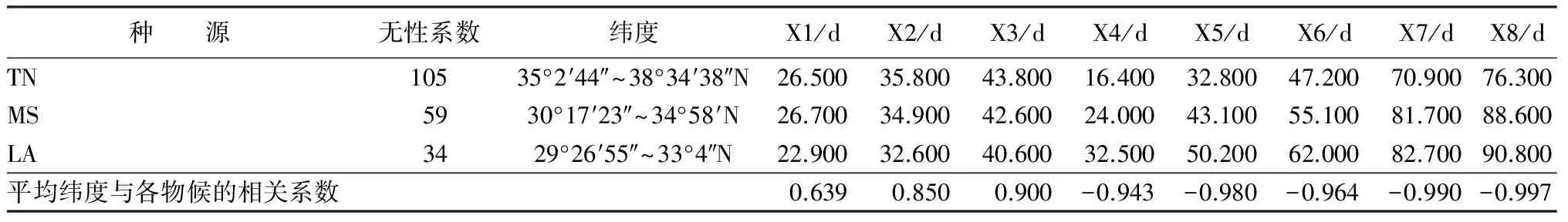
表7 不同种源平均物候期与纬度的相关性
4 结论与讨论
通过对443个南方型美洲黑杨无性系物候期的观测发现,其物候期存在着丰富的变异。总体来说,落叶阶段的物候期变异较大,5个不同物候期的极差跨度为45~64 d,而萌芽展叶阶段的物候期变异相对较小,3个不同物候期的极差跨度为21~24 d。因此,影响各无性系生长期长短的主要因素是生长后期与变色落叶相关的诸物候期的早晚差异。该结论与解荷锋等[19]对49个美洲黑杨无性系物候期的观察研究结论相一致,但与唐洁等[20]对60个美洲黑杨无性系苗期观测所得结论相反,在其研究中指出,与展叶相关的4个物候期的变异系数均大于后4个与落叶相关的物候期,生长期长短的影响因素主要是前期物候期的差异。出现不一致的主要原因在于本研究与解荷锋等[19]的研究对象是成年的美洲黑杨大树,而唐洁等[20]的研究对象是当年的扦插苗。杨树当年扦插苗的物候观测不但受到扦插时间早晚的影响,同时还受到穗条生根等一些关键因素的影响,另外,该试验材料主要是2个来源的无性系,材料背景相对单一,因此出现了结论不一致的现象。
物候期时序组成特征分析表明不同的物候期各有特色,时序特征明显不同,如443个美洲黑杨无性系在展叶始期仅出现4个不同时间点,而在叶变色始期和叶全部变色期则出现了11个不同时间点,而且不同物候期不同时间点出现的无性系比例差异较大,说明该美洲黑杨种质库物候多样性丰富。植物的物候与气温息息相关,特别在植物生长发育期的前期,各种物候期的开始日期与前期的气温之间有显著的相关性[22],本研究叶变色始期,85%的黑杨于9月10日和9月25日集中出现叶片变色,根据当时气象资料显示在这两个集中变色期的前一周左右出现了强烈降温天气,说明叶变色与气温较大降低密切相关。
通过对8个不同物候期的主成分分析,发现第一主成分载荷较高的都是与叶黄叶落有关的因子,第二主成分载荷较高的是与萌芽展叶有关的因子,第三主成分载荷较高的是落叶末期和全部落叶期,其中第一、第二、第三主成分累计贡献率达到了87.29%。该结论与文献中3个对杨树物候研究所得结论基本一致[19-21],都是第一主成分为落叶有关因子,第二主成分为展叶有关因子。
不同种源物候期与纬度的相关性分析结果表明,虽然这些无性系都是南方型美洲黑杨,且处于相同立地条件和管理下,但不同种源地的纬度与萌芽展叶物候期呈正相关,与叶变色落叶物候期呈明显的负相关。在3个种源中,路易斯安娜州种源是展叶最早,落叶最晚,属于展叶早落叶晚类型,相反,田纳西州种源是展叶最晚,落叶最早,属展叶晚落叶早类型,密西西比州种源则介于两者之间。这种不同种源间物候期与纬度存在相关关系的结论,在多种植物研究中也得到印证[23-25]。通过本研究的调查分析,说明不同无性系物候期的差异主要与美洲黑杨自身的遗传特性有关。不同的种质资源在多种性状上都存在差异,除了用形态学、分子标记来研究种质资源的多样性以外,在相同的环境条件下,利用物候期差异也可评价无性系的多样性。对于在调查研究中发现的特殊种质类型,对其加强观测研究作为特殊遗传种质材料进一步评价利用。
[1] 赵天锡.美洲黑杨及其杂种在世界和我国杨树栽培中的地位与作用[J].世界林业研究,1992(1):74-81.
[2] 张绮纹,苏晓华,李金花.美洲黑杨基因资源收存及其遗传评价的研究[J].林业科学,1999,35(2):31-37.
[3] 李世峰,张博,陈英,等.美洲黑杨种质资源遗传多样性的SSR分析[J].南京林业大学学报(自然科学版),2006,30(4):10-14.
[4] 樊孝萍,谯四红,张红,等.南方型美洲黑杨种质资源库的营建[J].湖北林业科技,2016,45(3):79-80.
[5] 解荷锋,于中奎,陈一山,等.黑杨派基因库内无性系生长特性的遗传分析[J].林业科学研究,1995,8(2):226-229.
[6] 管兰华,潘惠新,黄敏仁,等.美洲黑杨×欧美杨F1无性系的多性状联合选择[J].南京林业大学学报(自然科学版),2005,29(2):6-10.
[7] 李火根,黄敏仁,陈道明.美洲黑杨X青杨F1无性系生根性状的遗传变异及C效应[J].东北林业大学学报,1998,26(3):12-15.
[8] 赵丽,樊军锋,高建社,等.美洲黑杨与青杨人工杂交及杂种苗苗期性状研究[J].西北农业学报,2010,19(5):108-112.
[9] 李晓东,樊军锋,邱兴,等.美洲黑杨×青杨派杂种无性系苗期抗寒性的鉴定与筛选[J].西北林学院学报,2015,30(2):100-104.
[10] 黄秦军,黄国伟,丁昌俊,等.美洲黑杨杂种不同生长势无性系光合特征[J].林业科学,2013,49(3):56-62.
[11] 李火根,黄敏仁,潘惠新,等.美洲黑杨新无性系生长遗传稳定性分析[J].东北林业大学学报,1997,25(6):1-5.
[12] 唐洁,汤玉喜,苏晓华,等.美洲黑杨无性系苗期光合生理特性研究[J].中南林业科技大学学报,2014,34(9):12-16.
[13] 黄国伟,苏晓华,黄秦军.美洲黑杨不同生长势无性系生长和生理特征的差异[J].林业科学,2012,48(4):27-34.
[14] 唐镇敏.美洲黑杨南方种源的遗传变异[J].南京林业大学学报,1990,14(3)15-21.
[15] 王玲,段红平,田敏.美洲黑杨遗传转化系统优化的研究[J].云南农业大学学报,2011,26(4):519-523.
[16] 周文才,侯静,郭炜,等.基于SSR标记的美洲黑杨杂交子代的鉴定[J].南京林业大学学报(自然科学版),2015,39(3):45-49.
[17] 黄秦军,袁定昌,赵自成,等.美洲黑杨生长及基因表达差异[J].东北林业大学学报,2010,38(8):1-3.
[18] 龙凤,余传琼,吴承祯,等.不同种源雷公藤叶物候特征比较[J].应用与环境生物学报,2016,22(4):660-666.
[19] 解荷锋,于中奎,陈一山,等.美洲黑杨无性系物候期的观察研究[J].山东林业科技,1995(4):12-16.
[20] 唐洁,汤玉喜,李永进,等.美洲黑杨无性系苗期物候特征差异比较研究[J].中国农学通报,2013,29(31):15-20.
[21] 杨成生,王芳,张亚军,等.10个杨树品种的物候期研究[J].甘肃林业科技,2005,30(2):29-31.
[22] 李荣平,刘晓梅,周广胜.盘锦湿地芦苇物候特征及其对气候变化的响应[J].气象与环境学报,2006,22(4):30-34.
[23] 陈正洪.我国亚热带东部丘陵山区油桐物候特征分析[J].地理科学,1991,11(3):287-293.
[24] 宣继萍,周志芳,刘建秀,等.结缕草属植物物候期的差异分析[J].植物资源与环境学报,2008,17(3):53-57.
[25] 宗俊勤,郭爱桂,刘建秀.中国假俭草种质资源物候期的变异分析[J].中国草地学报,2006,28(6):61-67.
1)“十三五”国家重点研发计划课题(2016YFD0600402);湖北省技术创新专项重大项目(2016ABA111)。
杨杉,男,1979年9月生,湖北省林业科学研究院,副研究员。E-mail:306228796@qq.com。
张新叶,湖北省林业科学研究院,研究员。E-mail:1641135733@qq.com。
2017年3月1日。
S718.43
Phenological Character of Southern Type ofPopulusdeltoidesGermplasm//Yang Shan(Hubei Academy of Forestry, Wuhan 430075, P. R. China); Fan Xiaoping(Shishou Poplar Research Institute, Hubei Academy of Forestry); Su Xiaohua(Research of Forestry, CAF); Huang Guowei, Zhang Xinye(Hubei Academy of Forestry)//Journal of Northeast Forestry University,2017,45(7):7-11,17.
责任编辑:王广建。
We studied the phonological characters with 443Populusdeltoidesgermplasm in Shishou of Hubei Province. There were large differences among different phenophase of clones ofP.deltoides, different characteristics of composition occurred in different phenophase. There was the maximum variation in the phase of leaf color beginning with variation coefficient 52.74%, followed by phase of leaf color totally changed and phase of bud swell beginning. By principal component analysis, there were significantly correlation between phenological phases associated with discolored leaves, and phenological phases associated with bud phenology were also significant-related. The first principal component were factors associated with leaves discolor and fall, the second principal component were factors associated with the sprout and leaves expansion, the third principal component mostly loaded the late phase of leaves fall and the phase of totally leaves fall, of which the total contribution rate of the first, and the second and third principal component was 87.29%. Analysis of correlation between phenophase and latitude from different provenances showed that different provenance latitude was positively correlated with the phenophase of sprout and leaves expansion, and significantly negatively correlated with the phenophase of leaves discolor and fall.
猜你喜欢
特产研究(2022年6期)2023-01-17
农业工程学报(2022年12期)2022-09-09
热带作物学报(2022年6期)2022-07-08
小哥白尼(野生动物)(2021年12期)2021-03-29
辽宁林业科技(2021年1期)2021-03-16
娃娃乐园·综合智能(2019年10期)2019-12-07
中成药(2018年2期)2018-05-09
武夷学院学报(2017年9期)2017-11-29
西南农业学报(2017年5期)2017-06-23
河北林业科技(2016年5期)2016-11-08
