
番茄绿果与橙果间果实颜色及主要色素含量的遗传研究
2017-06-01 09:28赵菁菁罗佳伟李方曼李云洲
中国蔬菜 2017年3期
吴 浪 赵菁菁 罗佳伟 王 晨 李方曼 李云洲 张 颜 梁 燕
(西北农林科技大学园艺学院,陕西杨凌 712100)
番茄绿果与橙果间果实颜色及主要色素含量的遗传研究
吴 浪 赵菁菁 罗佳伟 王 晨 李方曼 李云洲 张 颜 梁 燕*
(西北农林科技大学园艺学院,陕西杨凌 712100)
对番茄组合绿樱(绿果)×金珠1号(橙果)的6个世代遗传群体(P1、P2、F1、BC1、BC2和F2)进行果色性状、番茄红素含量、叶绿素含量和胡萝卜素含量等的遗传规律分析。结果表明:正反交F1的果色性状无明显差异,而色素含量存在显著差异;说明番茄果色性状受核基因控制,而色素含量遗传除受核基因控制外还可能存在胞质效应。采用多世代联合分析法的分析结果表明,番茄绿果与橙果间的果色性状符合2对加性主基因+加性-显性多基因(MX2-A-AD)遗传模型,其BC1、BC2和F2主基因遗传率分别为73.42%、78.25%和61.41%,多基因遗传率分别为22.87%、15.35%和34.94%,即果色性状遗传的主基因遗传力较强;叶绿素含量符合1对负向显性主基因+加性-显性多基因(MX1-AEND-AD)遗传模型,其BC1、BC2和F2主基因遗传率分别为0、1.73%和0.65%,多基因遗传率分别为45.47%、0和37.82%,即主基因遗传力在BC2群体中最高,多基因遗传力在BC1群体中最高;番茄红素含量与胡萝卜素含量均符合2对加性-显性-上位性主基因+加性-显性多基因(MX2-ADI-AD)遗传模型,其BC1、BC2和F2主基因遗传率分别为75.74%、1.79%、84.26%和61.53%、87.21%、81.05%,多基因遗传率分别为20.32%、74.12%、12.68%和0.68%、0、0,表明番茄红素含量和胡萝卜素含量的主基因遗传力较强。
番茄;果色遗传;色素含量遗传;多世代联合分析法
番茄(Lycopersicon esculentum Mill.)果实颜色种类丰富多样,有红色、粉色、黄色、绿色、橙色和紫色等。在对番茄紫色果(阮美颖,2013)、黑色果(林涛 等,2013)和棕色果(肖良军 等,2015)果色的遗传研究中发现,果色在不同颜色种类番茄间的遗传存在差异,即番茄果色性状的遗传具有多样性(孙亚东 等,2009;龚亚菊 等,2013)。同时,果色作为番茄重要的商品品质之一,对番茄产业影响较大。所以关于不同颜色种类番茄间果色性状遗传规律的研究对培育优质特色番茄品种、满足不同偏好消费群体要求,甚至整个番茄产业的发展都具有重要意义。
番茄果实颜色的形成与果实中所含色素种类及其含量存在着直接联系。对不同果色番茄果实中色素组成和含量的研究结果表明,番茄果实颜色的形成主要取决于果实中所含色素的组成、含量和比例(孟凡娟 等,2006;赵润洲和刘鸣韬,2011);此外,果实中色素的含量也历来作为评价番茄营养品质的重要指标之一,特别是番茄红素和胡萝卜素(李晓蕾 等,2010)。对于绿果番茄果实中含量较多的叶绿素,近年来也引起了人们的重视,已有医学研究表明叶绿素具备激活细胞代谢机能、抗氧化、抑制氧自由基对细胞膜的损伤、调节血糖、促进造血机能等药理作用(黄寿吾,2006)。综上可知,色素含量遗传规律的研究对于番茄果色遗传研究及培育优质特色番茄新品种具有重要意义。
截至目前,对番茄果实中番茄红素含量遗传规律的研究已有较多报道(李纪锁 等,2006;曲瑞芳和梁燕,2009;孟昭娟 等,2011),而关于叶绿素含量或胡萝卜素含量的遗传规律研究大多集中在黄瓜(赵建华 等,2005)、辣椒(张宝玺 等,1996)、甜瓜(吕丽华 等,2012)和玉米(王爱玉和张春庆,2008)等作物上,在番茄上还鲜有报道。
本试验以绿果番茄和橙果番茄为试材,在研究番茄绿果与橙果间果实颜色性状遗传规律的同时,对果实中叶绿素含量、番茄红素含量和胡萝卜素含量的遗传规律进行了分析,并对两者的遗传规律进行了初步比较,以期丰富番茄果色遗传和色素含量遗传的基础理论,为培育具备特色果色和特殊营养品质的番茄新品种提供参考。
1 材料与方法
1.1 试验材料
供试番茄材料绿樱(P1)和金珠1号(P2)均为纯系,由西北农林科技大学园艺学院番茄课题组供种,具体性状特征见表1。

表1 试验材料及其相关性状
以绿樱和金珠1号为亲本构建6个世代(P1、P2、F1、BC1、BC2和F2)遗传群体,并同时种植;其中P1、P2、F1各种植25株,BC1种植51株,BC2种植50株,F2种植154株。果实完全成熟期,每株选取第2、第3穗果中发育正常、达到商品成熟度的果实10~15个,进行果色和色素含量的观察、测定。
本试验先后于2013~2014年和2015年在西北农林科技大学新天地试验基地的同一塑料大棚内进行初试验和验证试验。因验证试验与初试验结果基本一致,为节约篇幅,故本文只针对初试验进行了详细论述。
1.2 试验方法
1.2.1 果色性状测定 本试验对果色性状的测定采用了目测和色差仪测定2种方法。目测法:横切经色差仪测定过的番茄果实,结合标准比色卡,目测其果色、果皮色、果肉色和胎座胶状物质颜色,进行孟德尔式分离分析,并进行分离比的卡方检验。色差仪测定法:利用CHROMA METE CR-400全自动式色差仪测定番茄果实表面颜色色差值。操作要点:沿果实赤道面一周均匀取3点进行测定,直接得出L、a、b 3个数值。其中,L值反映颜色的明亮程度,0表示黑色,100表示白色;a值反映红色或绿色物质的浓度,a>0表示颜色偏红,a<0表示颜色偏绿;b值反映橙色或蓝色物质的浓度,b>0表示颜色偏橙,b<0表示颜色偏蓝。每株测定5~6个果,取平均值,计算色光值=2 000× a/L×(a2+b2)1/2,其主要反映物质表面颜色(周蓉 等,2012;吴浪 等,2016)。
1.2.2 色素提取及测定 在果实成熟期,每株选取第2、第3穗果中发育正常、达到商品成熟度的果实10~15个,4 ℃冷库预处理12 h,避光或弱光条件下打成匀浆,称取2.7~2.9 g匀浆2份,分别移入2支10 mL离心管中。一份加入80%丙酮,摇匀,放入超声波(100 W、30 ℃)进行叶绿素和胡萝卜素的提取,充分提取后将提取液转入25 mL棕色容量瓶中,并用80%丙酮定容(马迎杰 等,2005;张秀君 等,2011),然后采用DU-800型分光光度计比色测定其在663、646 nm和470 nm下的吸光值,利用公式法(阮美颖,2013)算出叶绿素含量和胡萝卜素含量,3次重复;另一份经无水甲醇处理后,加入含有2%二氯甲烷的石油醚,放入超声波(100 W、30 ℃)进行番茄红素的提取,充分提取后将提取液转入25 mL棕色容量瓶中,并用含有2%二氯甲烷的石油醚定容(张卫强和邓宇,2002;王燕 等,2003),然后采用DU-800型分光光度计比色测定其在502 nm下的吸光值,利用公式法(阮美颖,2013)算出番茄红素含量,3次重复。
1.2.3 统计分析方法 依照盖钧镒(2005)的多世代联合分析法,同时借助曹锡文等(2013)开发的植物数量性状分离分析Windows软件包SEA对试验数据进行分析,然后根据赤池信息准则(Akaike’s information criterion,AIC)最小原则及模型适合性检验〔包括均匀性检验U12、U22、U32,Smirnov检验(nW2),Kolmogorov检验(Dn)〕,从包括1对主基因、2对主基因、多基因、1对主基因+多基因和2对主基因+多基因等24个模型中选择最优遗传模型,并估计相应一阶遗传参数和二阶遗传参数。采用SPSS 22.0软件(Tukey HSD法)进行差异显著性分析。
2 结果与分析
2.1 各世代果色性状及色素含量表型分析
根据比色卡比对结果(表2),绿樱与金珠1号正反交F1成熟果实颜色性状均与金珠1号相同:果皮黄色,果肉浅橙色,胎座胶状物质浅橙色,果色橙色,说明正反交F1果皮、果肉、胎座胶状物质和果实颜色均为核基因控制,不受细胞质基因的影响。而在BC1、BC2和F2中部分果色性状发生了分离。F2果实颜色,橙色∶红色∶黄色∶绿色= 9∶3∶3∶1,χ2=3.48,P >0.05;果皮颜色,黄色∶透明=3∶1,χ2=1.69,P>0.05;果肉颜色,浅橙∶浅红∶浅黄∶浅绿=9∶3∶3∶1,χ2=3.48,P>0.05;胎座胶状物质颜色,浅绿∶黄绿∶浅橙∶浅黄=9∶3∶3∶1,χ2=47.47,P<0.05;即F2果皮、果肉和果实颜色遵循孟德尔遗传规律,但胎座胶状物质颜色并未呈现出经典的孟德尔式分离比。BC1果实颜色,橙色∶红色∶黄色∶绿色= 1∶1∶1∶1,χ2=2.72,P>0.05;果皮颜色,黄色∶透明=1∶1,χ2=0.08,P>0.05;果肉颜色,浅橙∶浅 红∶浅 黄∶浅 绿=1∶1∶1∶1,χ2=2.72,P>0.05;胎座胶状物质颜色,浅绿∶黄绿∶浅黄= 10∶3∶4,不符合经典的孟德尔式分离比。在BC2中,果皮、果肉和果实颜色均未发生分离,依次分别为黄色、浅橙色和橙色;胎座胶状物质颜色为黄绿∶浅橙=16∶9,不符合经典的孟德尔式分离比。
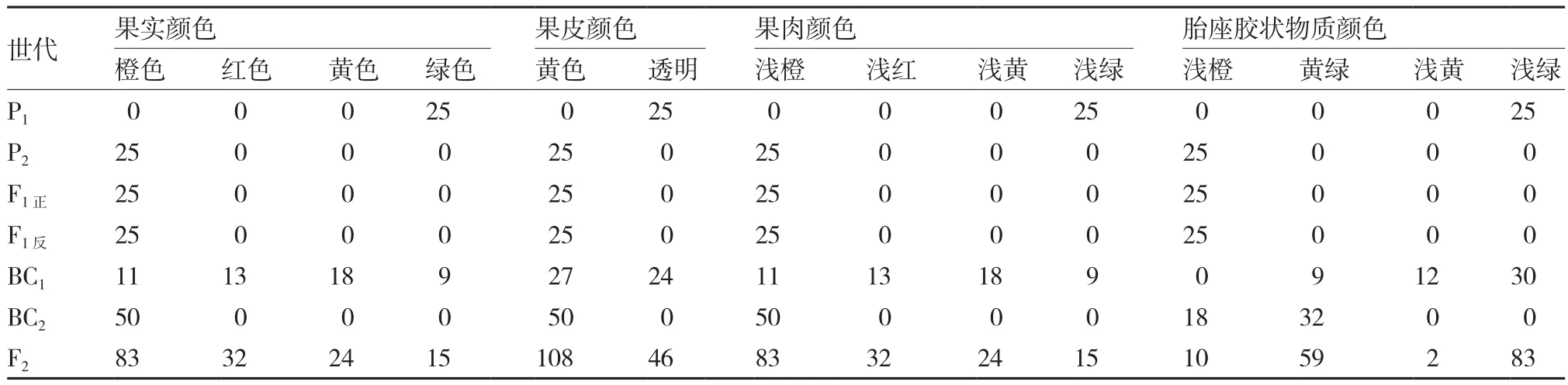
表2 绿樱×金珠1号各世代果实颜色性状
上述结果表明,番茄绿果与橙果间果色的遗传均为2对核基因控制;在各世代中,果肉颜色的分离比与果实颜色的分离比始终相同,表明番茄果实颜色最易受果肉颜色的影响,而受果皮颜色和胎座胶状物质颜色的影响较小。
由表3可知,绿樱与金珠1号F1的色光值正交与反交差异不显著,表明该组合果色性状的遗传由核基因控制;叶绿素含量、番茄红素含量和胡萝卜素含量F1正反交差异显著,说明色素含量的遗传除受核基因控制外还可能存在胞质遗传效应;另外,从亲本与F1的差异显著性分析结果来看,大多数测定指标存在显著性差异,表明该组合果色性状和色素含量的遗传可能存在显性或上位性效应。
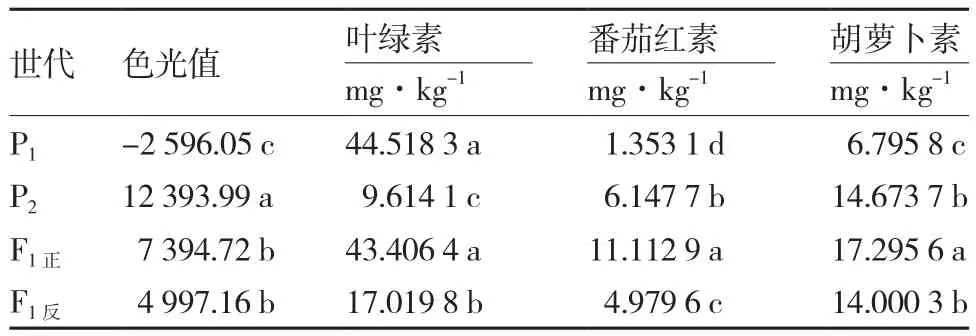
表3 绿樱(P1)与金珠1号(P2)及其正反交F1的果实色光值及色素含量
进一步分析各测定指标表型值在F2中的次数分布(图1),色光值均值为7 539.14,在5 000~10 000间存在的植株数最多;叶绿素含量均值为32.9 mg·kg-1(FW),在25.0~30.0 mg·kg-1间存在的植株数最多;番茄红素含量均值为12.6 mg·kg-1(FW),大多集中于3.0 mg·kg-1;胡萝卜素含量均值为9.6 mg·kg-1(FW),10.0 mg·kg-1处的植株数最多。即在F2中大多数植株的4项指标均偏向高值亲本。从峰值的个数来看,色光值存在2个峰,叶绿素含量存在3个峰,番茄红素含量呈偏峰分布,胡萝卜素含量存在3个峰;表明该组合果色性状和色素含量的遗传呈明显的主基因+多基因遗传特征。
2.2 果色性状的主基因+多基因混合遗传分析
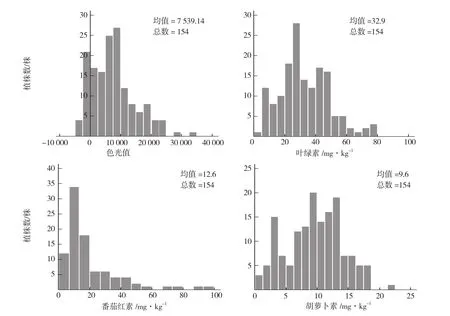
图1 绿樱×金珠1号F2色光值和色素含量的次数分布
2.2.1 最优遗传模型的选择 以绿樱×金珠1号的P1、P2、F1、BC1、BC2和F2的果实表面色光值为基础,运用多世代联合分析法,对这6个世代材料进行分析,得到1对主基因、2对主基因、多基因、1对主基因+多基因和2对主基因+多基因等5类共24个不同遗传模型的AIC值(表4)。根据AIC值最小原则,从中选出3个AIC值较小且差异不大的遗传模型:MX2-ADI-ADI、MX2-A-AD和MX2-EEAD-AD,均为2对主基因+多基因遗传模型。
然后对初选遗传模型进行群体适合性检验,分别统计各模型中达到显著水平的统计量个数。由表5可知,模型MX2-EEAD-AD中达到显著水平的统计量为6个,而模型MX2-ADI-ADI和MX2-AAD均为2个,表明后两者与分布群体不一致的统计量个数较少;又因MX2-A-AD模型的AIC值较MX2-ADI-ADI小,故按照试验群体的适合性检验统计量与模型差异越少越优和AIC值最小的原则,确定该组合果色性状的最优遗传模型为MX2-AAD,即2对加性主基因+加性-显性多基因遗传。
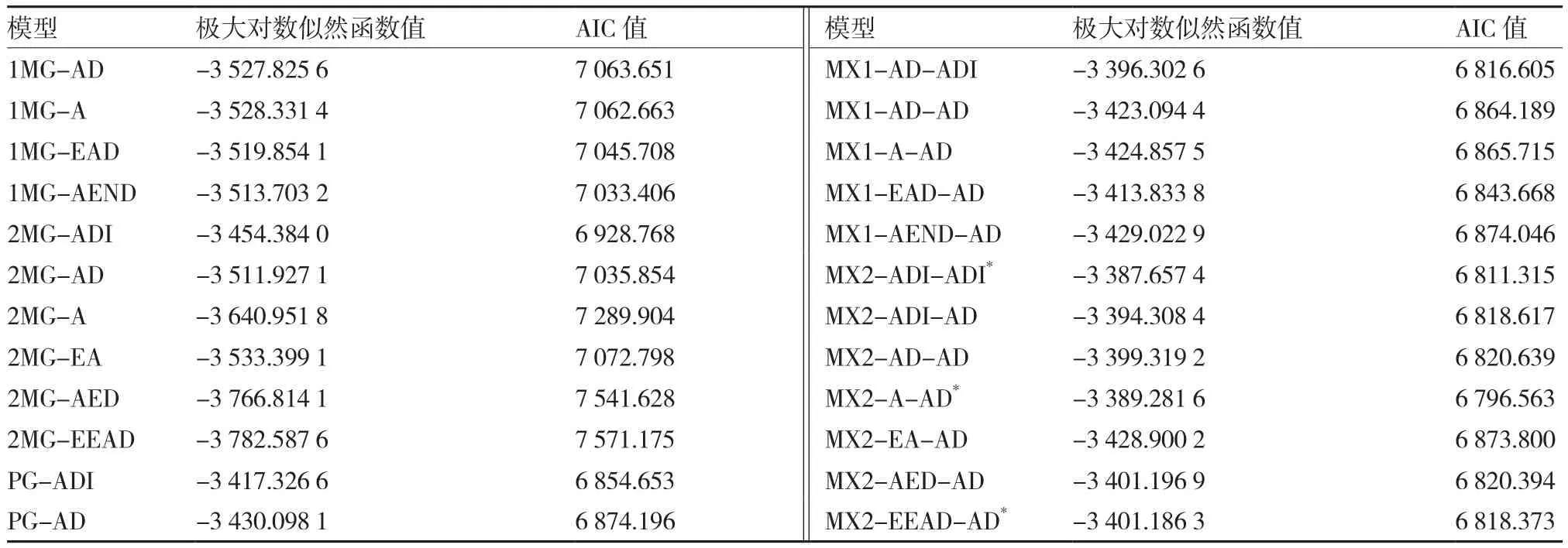
表4 绿樱×金珠1号果色性状不同遗传模型下的极大对数似然函数值和AIC值
2.2.2 最优遗传模型遗传参数的估计 表6为该组合果色性状最优遗传模型MX2-A-AD下的一阶遗传参数和二阶遗传参数估计值,结果显示控制番茄绿果与橙果间果色性状遗传的有2个主效基因:第1主效基因和第2主效基因,均只存在加性效应而无显性效应;它们的加性效应值分别为1 219.076和2 300.925,表明第2主效基因的加性效应较为明显。多基因的加性效应值和显性效应值分别为-12 117.6和-706.056,即多基因加性效应较为明显。遗传率方面,BC1、BC2和F2的主基因遗传率分别为73.42%、78.25%和61.41%,多基因遗传率分别为22.87%、15.35%和34.94%,表明该组合主基因表现的遗传力较高。
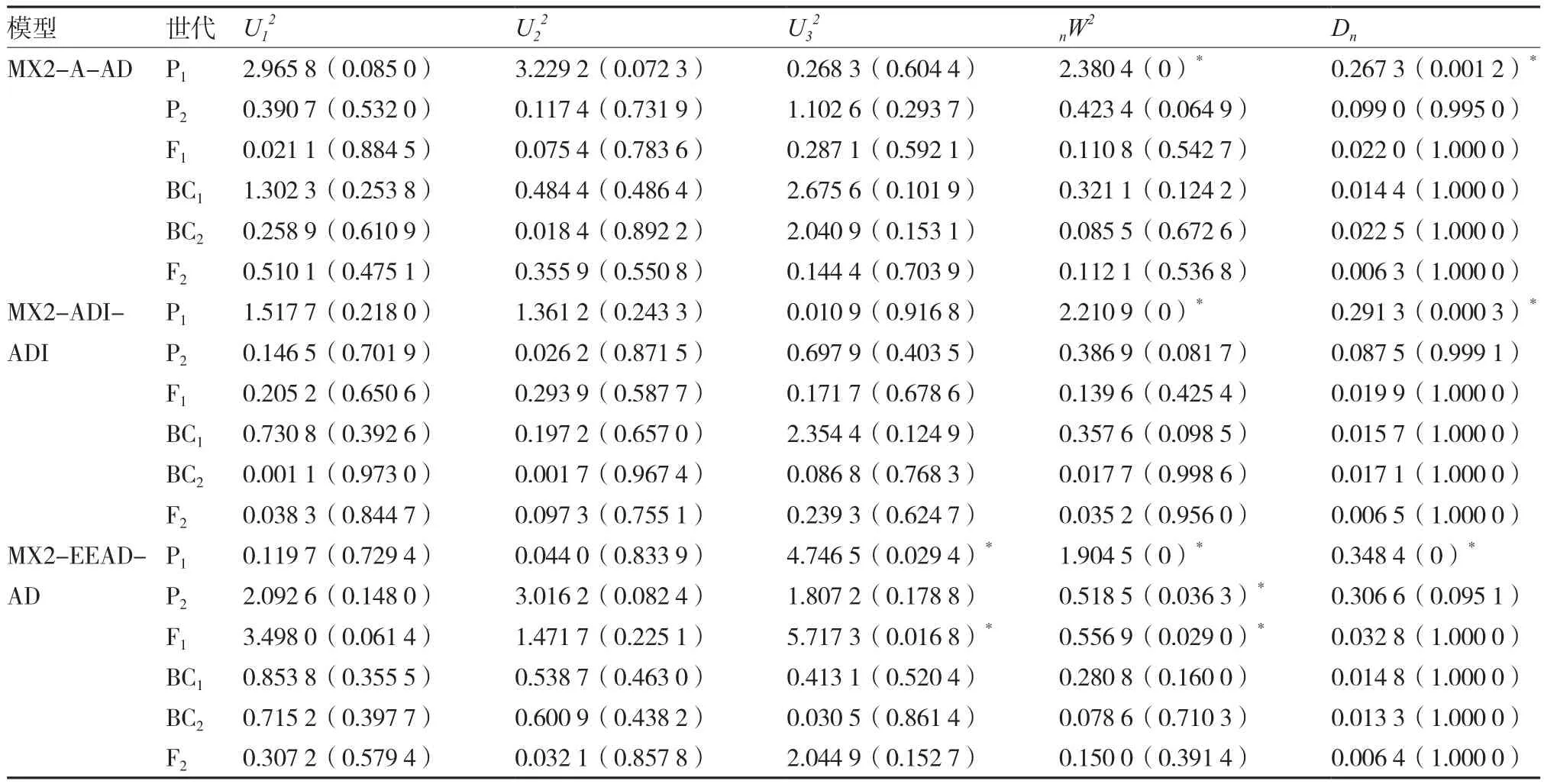
表5 绿樱×金珠1号果色性状初选遗传模型的适合性检验
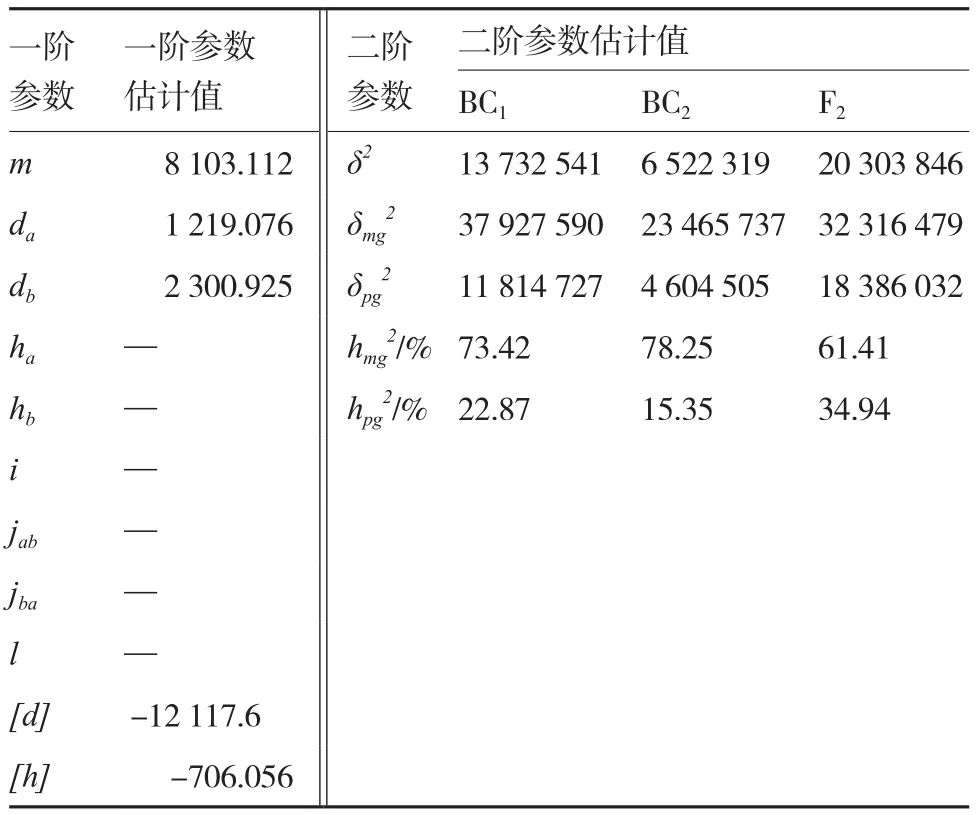
表6 绿樱×金珠1号果色性状的遗传参数估计值
2.3 色素含量的主基因+多基因混合遗传分析
前人研究表明,同一色素在不同颜色番茄果实中的含量不同,同一颜色番茄果实中各色素的含量也存在差异(赵润洲和刘鸣韬;2011),说明番茄果实颜色的形成不是由一种色素决定的而是取决于多种色素的组成、含量和比例。因此,本试验对同一遗传群体番茄果实中的叶绿素含量、番茄红色含量和胡萝卜素含量进行了遗传分析。
2.3.1 最优遗传模型的选择 与果色性状遗传分析类似,运用多世代联合分析法,根据AIC值最小原则,分别初选出3个遗传模型后,对其进行群体适合性检验,按照试验群体的适合性检验统计量与模型差异越少越优的原则,同时结合AIC值确定色素含量的最优遗传模型。如表7所示,叶绿素含量、番茄红素含量和胡萝卜素含量的最优遗传模型分别为MX1-AEND-AD、MX2-ADI-AD和MX2-ADIAD,即番茄红素含量和胡萝卜素含量符合同一遗传模型MX2-ADI-AD,从遗传模型上看其与叶绿素含量遗传的主要区别在于叶绿素含量的遗传为1对负向显性主基因,而番茄红素含量和胡萝卜素含量的遗传为2对加性-显性-上位性主基因。
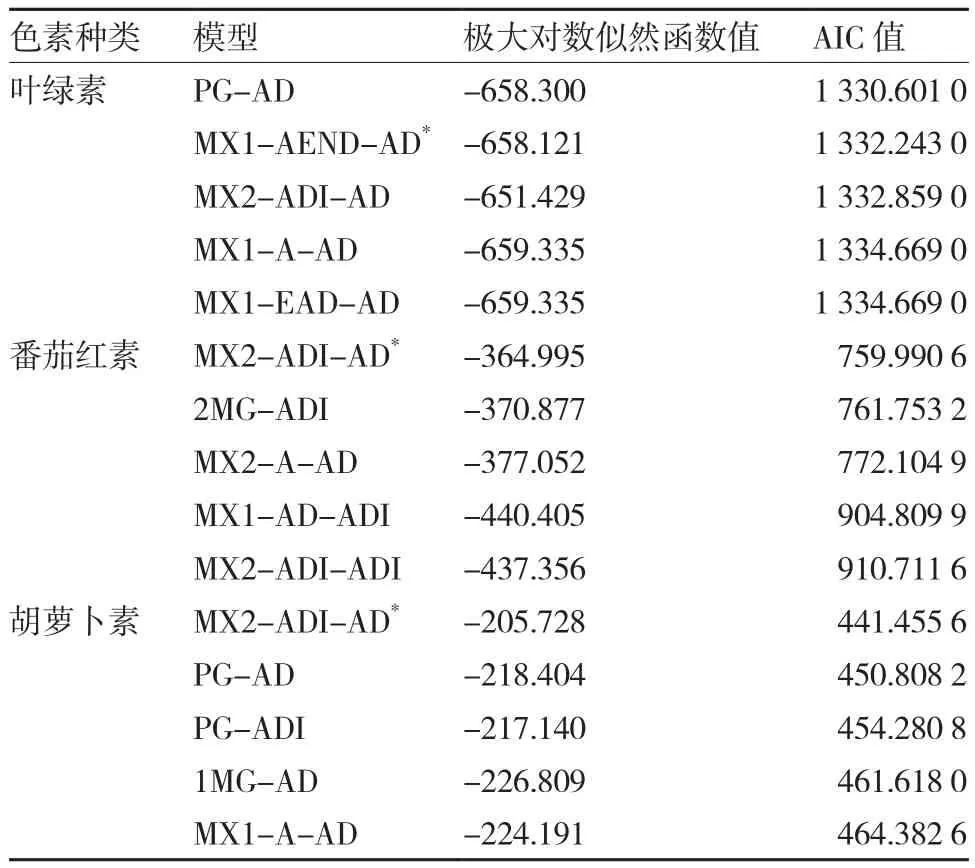
表7 绿樱×金珠1号色素含量初选遗传模型下的极大对数似然函数值和AIC值
2.3.2 最优模型遗传参数的估计 由表8、9可知,该组合叶绿素含量的主基因加性效应值和显性效应值分别为0.008 3和-0.008 3,表明主基因加性效应和显性效应同等重要;多基因加性效应值和显性效应值分别为0.537 1和3.716 3,即多基因显性效应更为明显。从二阶遗传参数估计值来看,BC1、BC2和F2主基因遗传率分别为0、1.73%和0.65%,多基因遗传率分别为45.47%、0和37.82%,表明主基因遗传率较低,多集中在BC2,多基因遗传率较高,多集中在BC1,即叶绿素含量多基因遗传力较主基因遗传力高。
番茄红素含量第1、第2主基因的加性效应值、显性效应值均为负值,表示效应作用的方向趋向于高值亲本;其中第1主基因加性效应值、显性效应值和第2主基因加性效应值、显性效应值依次分别为-0.070 9、-2.119 3、-0.070 9、-3.611 2,多基因加性效应值和显性效应值分别为-0.151 2和0.327 2,表明主基因和多基因的显性效应均较加性效应明显。此外,BC1、BC2和F2主基因遗传率分别为75.74%、1.79%和84.26%,多基因遗传率分别为20.32%、74.12%和12.68%,表明番茄红素含量的主基因遗传力较多基因遗传力强。
胡萝卜素含量的遗传与番茄红素含量较为相似,第1、第2主基因的加性效应值、显性效应值也均为负值,多基因加性效应值和显性效应值分别为0.122 8和0.065 1,即遗传效应以加性效应为主。遗传率方面,BC1、BC2和F2主基因遗传率分别为61.53%、87.21%和81.05%,多基因遗传率分别为0.68%、0和0,表明主基因遗传力较高。
综上,该组合果实中叶绿素含量、番茄红素含量和胡萝卜素含量的遗传均为主基因+多基因的遗传方式;遗传效应以显性效应为主,遗传率方面叶绿素含量是多基因遗传力较强,番茄红素含量和胡萝卜素含量却是主基因遗传力较强。
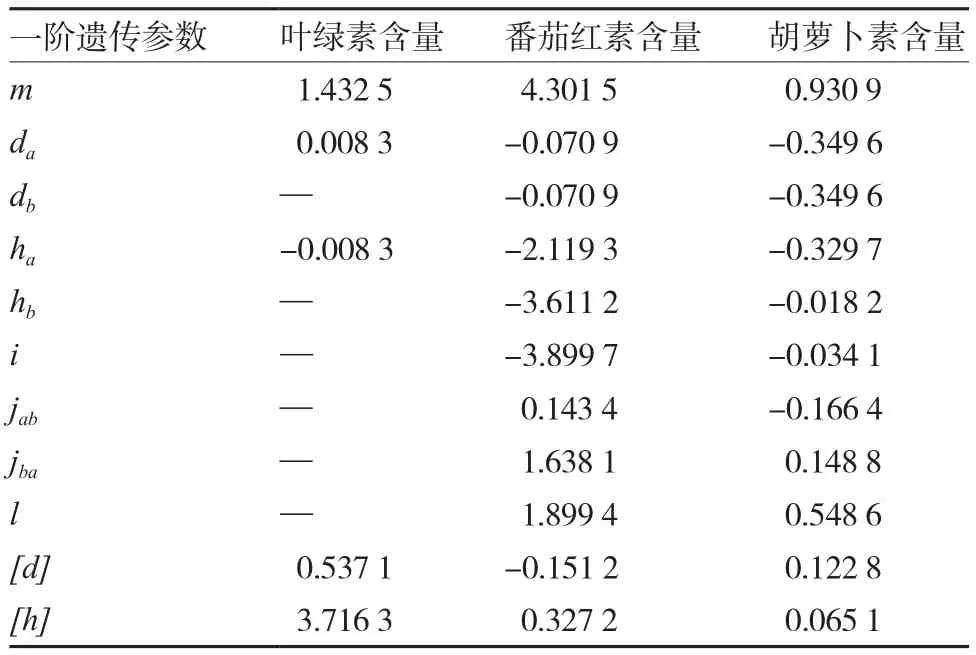
表8 绿樱×金珠1号色素含量的一阶遗传参数估计值
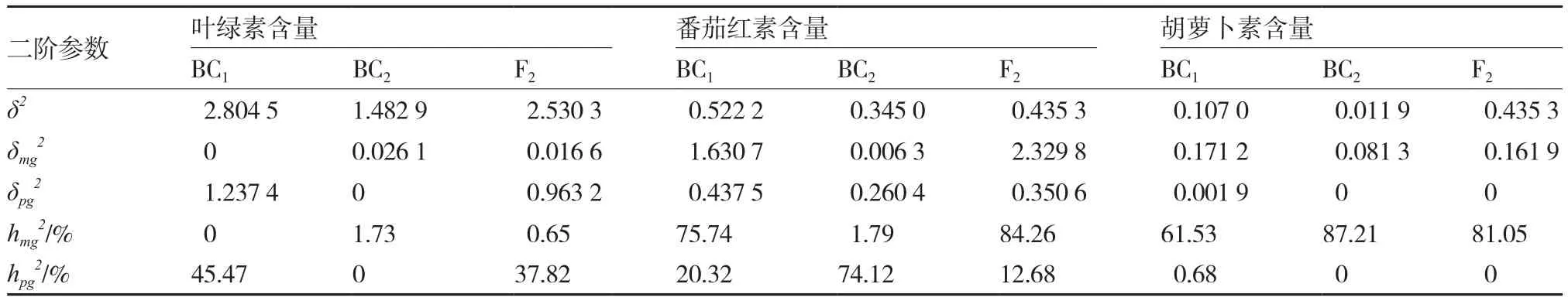
表9 绿樱×金珠1号色素含量的二阶遗传参数估计值
3 结论与讨论
番茄成熟果实颜色种类多样,多被作为质量性状进行研究(吕书文 等,2004;阮美颖,2013;肖良军 等,2015),得到的研究结果也多是质量性状遗传。据Ballester等(2010)报道,番茄果实颜色的形成主要由果皮颜色和果肉颜色共同决定,受胎座胶状物质颜色的影响较小。除了控制果皮颜色的基因Y-y和控制果肉颜色的基因R-r和T-t外,涉及果实颜色的基因还有B、del、dg、hp、ogc、mob等(Kallo,1991;Ronen et al.,2000),这些基因的不同组合使番茄果实表现出了不同的颜色。在本试验中,绿果番茄与橙果番茄间果色性状的遗传存在连续性变异特点,在分离群体F2和BC1中出现了肉眼难辨的中间型过渡色。由此表明,番茄果色性状并不是简单的质量性状。为此,本试验借鉴周蓉等(2012)在定量分析番茄果实中番茄红素含量时将色差仪测定值L、a和b转换为以它们为变量的色光值〔色光值=2 000×a/L×(a2+b2)1/2〕的方法,对番茄果色性状的遗传进行了数量性状遗传分析。结果表明,绿果番茄与橙果番茄间果色性状的遗传符合2对加性主基因+加性-显性多基因(MX2-A-AD)遗传模型,即番茄果色性状的遗传也有数量性状的特点。
除了受基因控制外,番茄果实颜色的形成还直接与果皮中类黄酮、果肉中叶绿素和类胡萝卜素的组分和含量有关(Fray & Grierson,1993;Delgado-Vargas et al.,2000;Rosati et al.,2009;Ballester et al.,2010)。红色番茄果实中番茄红素含量较多,绿色番茄果实中叶绿素含量较多,橙色番茄果实中胡萝卜素含量较多,紫色番茄果实中花青素含量较多,棕色番茄则与类胡萝卜素和叶绿素的累积有关(孟凡娟 等,2006;赵润洲和刘鸣韬,2011),说明番茄果实颜色主要取决于番茄红素、叶绿素、胡萝卜素和花青素等有色色素的含量和相对比例。
目前,关于番茄果实中色素含量遗传规律的研究多集中于番茄红素。刘进生和赵有为(1986)研究表明,番茄红素含量遗传模型以加性效应为主,存在少量上位性效应,而显性效应不显著。李纪锁等(2006)得出番茄红素含量的遗传符合1个加性主基因+加性-显性-上位性多基因模型。曲瑞芳和梁燕(2009)却表示番茄红素含量遗传符合加性-显性模型。孟昭娟等(2011)的研究结论是番茄红素含量的遗传符合1个完全加性的主效基因+加性-显性多基因遗传模型。而本试验采用6个世代联合分析法,发现番茄果实中番茄红素含量的遗传为2对加性-显性-上位性主基因+加性-显性多基因(MX2-ADI-AD)遗传模型,与前人研究结果的主要差异是产生遗传效应的主基因有2对,究其原因可能是试验所选用的品种和调查的生长时期不同,或是生长环境不同。
另外,本试验还对番茄果实中叶绿素含量和胡萝卜素含量的遗传规律进行了分析。结果表明,叶绿素含量的遗传符合1对负向显性主基因+加性-显性多基因(MX1-AEND-AD)遗传模型;而胡萝卜素含量与番茄红素含量的遗传模型相同,均为2对加性-显性-上位性主基因+加性-显性多基因(MX2-ADI-AD)遗传模型。依据色素代谢途径推测,出现这一结果的原因很可能是番茄红素和胡萝卜素同属于类胡萝卜素代谢途径,且二者存在上下游关系。
在比较番茄果色与色素含量遗传规律异同时发现,番茄果色性状、叶绿素含量、番茄红素含量和胡萝卜素含量的遗传均符合主基因+多基因的植物数量性状遗传规律,其最优遗传模型分别是MX2-A-AD、MX1-AEND-AD、MX2-ADI-AD和MX2-ADI-AD。根据最优遗传模型不难看出,它们的主要区别在于果色性状的主基因仅存在加性效应,而色素含量的主基因除了加性效应还有显性效应和上位性效应;相同的是多基因均有加性效应和显性效应;此外,根据一阶遗传参数和二阶遗传参数可知,果色性状在后代中的遗传与番茄红素含量和胡萝卜素含量较为接近,遗传效应均以加性效应为主,且在子代中遗传力均是主基因遗传力较强;而叶绿素含量在后代中的遗传力却是多基因遗传力表现较强。表明果色性状的遗传与番茄红素含量和胡萝卜素含量的遗传更为接近,而与叶绿素含量的遗传存在着较大差异。原因可能是在番茄成熟果实中叶绿素多累积于胎座周围,而番茄红素和胡萝卜素多累积在果皮和果肉中。因此,在今后的研究中可通过番茄果实各部位色素含量的测定来推测果色性状的遗传和色素间的关系。
参考文献
曹锡文,刘兵,章元明.2013.植物数量性状分离分析Windows软件包SEA的研制.南京农业大学学报,36(6):1-6.
盖钧镒.2005.植物数量性状遗传体系的分离分析方法研究.遗传,27(1):130-136.
龚亚菊,吴丽艳,黎志彬,鲍锐,钟利.2013.大果番茄种质资源的遗传多样性和聚类分析.西南农业学报,26(6):2447-2450.
黄寿吾.2006.叶绿素的药理和临床应用.食品与药品,8(4):5-8.
李纪锁,沈火林,石正强.2006.鲜食番茄果实中番茄红素含量的主基因-多基因混合遗传分析.遗传,28(4):458-462.
李晓蕾,李景富,康立功,张贺,许向阳.2010.番茄品质遗传及育种研究进展.中国蔬菜,(14):1-7.
林涛,李锦泉,黄青峰,陈朝文,林金秀.2013.樱桃番茄果色的遗传分析.福建农业学报,28(10):987-992.
刘进生,赵有为.1986.番茄果实内番茄红素含量的遗传.遗传,8(2):9-12.
吕丽华,王怀松,付秋实,朱慧芹,张延国.2012.甜瓜果肉β-胡萝卜素含量的遗传分析.中国蔬菜,(14):45-50.
吕书文,李海涛,孟玲,许文奎,邹庆道.2004.番茄未熟果深绿色的遗传研究.辽宁农业科学,(2):5-6.
马迎杰,周宝利,程孙亮,付亚文.2005.超声波提取番茄果实中类胡萝卜素条件的优化.植物生理学通讯,41(5):110-112.
孟凡娟,许向阳,李景富.2006.番茄果实色素含量和表面颜色相关性研究.东北农业大学学报,37(4):459-462.
孟昭娟,刘姝红,姜晶.2011.鲜食番茄果肉中番茄红素含量性状的遗传分析.中国农学通报,27(5):452-456.
曲瑞芳,梁燕.2009.番茄果实中番茄红素含量的遗传分析.华北农学报,24(s1):46-49.
阮美颖.2013.紫色番茄果实主要色素积累、果色遗传规律及种质创新研究〔硕士论文〕.杭州:浙江师范大学.
孙亚东,梁燕,吴江敏,张飞,闫见敏.2009.番茄种质资源的遗传多样性和聚类分析.西南农业学报,18(5):297-301.
王爱玉,张春庆.2008.玉米叶绿素含量的QTL定位.遗传,30(8):1083-1091.
王燕,胡亚平,张喻,曾意孟.2003.影响番茄红素提取效果的因素的研究.食品科技,(5):62-64.
吴浪,刘婧仪,梁燕.2016.番茄绿果与红果颜色性状遗传的研究.园艺学报,43(4):674-682.
肖良军,陆佳楠,李宁,刘小茜,杨文才.2015.棕果番茄果实颜色遗传及gf位点序列变异分析.园艺学报,42(1):38-46.
张宝玺,郭家珍,杨桂梅,徐光,东惠茹.1996.辣椒绿熟期果色及主要色素含量的遗传.园艺学报,23(1):94-96.
张卫强,邓宇.2002.微波辐射萃取番茄红素的研究.食品工业科技,23(5):36-37.
张秀君,孙钱钱,乔双,朱海,孙丹,江丕文.2011.菠菜叶绿素提取方法的比较研究.作物杂志,(3):57-59.
赵建华,张建农,李浩霞,范瑞霞.2005.瓜类作物叶绿素含量的遗传特性研究.甘肃农业大学学报,40(1):13-16.
赵润洲,刘鸣韬.2011.番茄果实色泽与色素组成的关系.河南农业科学,40(9):98-100.
周蓉,蒋芳玲,梁梅,邹滔,刘小娟,吴震.2012.用色差仪法定量分析番茄果实番茄红素的含量.江西农业学报,24(9):45-48.
Ballester A R,Molthoff J,de Vos R,Hekkert B L,Orzaez D,Fern á ndez-Moreno J P,Tripodi P,Grandillo S,Martin C,Heldens J,Ykema M,Granell A,Bovy A.2010.Biochemical and molecular analysis of pink tomatoes:deregulated expression of the gene encoding transcription factor SlMYB12 leads to pink tomato fruit color.Plant Physiology,152:71-84.
Delgado-Vargas F,Jim é nez A R,Paredes-L ó pez O.2000.Natural pigments:carotenoids,anthocyanins,and betalainscharacteristics,biosynthesis,processing,and stability.Critical Reviews in Food Science and Nutrition,40(3):173-289.
Fray R G,Grierson D.1993.Identification and genetic analysis of normal and mutant phytoene synthase genes of tomato by sequencing,complementation and co-suppression.Plant Molecular Biology,22:589-602.
Kallo G.1991.Genetic improvement of tomato.Theor Appl Genet,14:386-392.
Ronen G,Carmel-Goren L,Zamir D,Hirschberg J.2000.An alternative pathway to β-carotene formation in plant chromoplasts discovered by map-based cloning of Beta and old-gold color mutation in tomato.Proceedings of the National Academy of Sciences,97(20):11102-11107.
Rosati C,Diretto G,Giuliano G.2009.Biosynthesis and engineering of carotenoids and apocarotenoids in plants:state of the art and future prospects.Biotechnology and Genetic Engineering Reviews,26(1):139-162.
Abstract:A genetic analysis was conducted to study fruit color,lycopene content,chlorophyll content and carotene content of the genetic populations(P1,P2,F1,BC1,BC2and F2)generated by crossing green fruit tomato inbred line‘Lyuying’and orange fruit tomato inbred line‘Jinzhu No.1’.The result showed that there was no significant differences in fruit color of F1generation groups.However there were significant differences in pigment content of F1generation groups,indicating fruit color was controlled by nuclear gene,while genetic of pigments content in F1was not only controlled by nuclear gene,but also there existed some cytoplasm effects.The result of analyzing by multiple generation conjoint showed that the genetics of tomato fruit color between green and orange were accord with the pattern of MX2-A-AD.The major gene heritability and the polygene heritability in BC1,BC2and F2was 73.42%,78.25%,61.41% and 22.87%,15.35%,34.94%,respectively.The genetics of chlorophyll content was accord with MX1-AEND-AD.The major gene heritability and the polygene heritability in BC1,BC2and F2was 0,1.73%,0.65% and 45.47%,0,37.82%,respectively.The genetics of lycopene content,as with genetics of carotene content,was accord with MX2-ADI-AD.The major gene heritability of lycopene content and carotene content in BC1,BC2and F2was 75.74%,1.79%,84.26% and 61.53%,87.21%,81.05%,respectively.And the polygene heritability of them was 20.32%,74.12%,12.68% and 0.68%,0,0,respectively.It indicated that the major gene heritability of lycopene content and carotene content was stronger.
Studies on Inheritance of Fruit Color and Major Pigment Contents between Green and Orange of Tomato(Lycopersicon esculentum Mill.)
WU Lang,ZHAO Jing-jing,LUO Jia-wei,WANG Chen,LI Fang-man,LI Yun-zhou,ZHANG Yan,LIANG Yan*
(College of Horticulture,Northwest A&F University,Yangling 712100,Shaanxi,China)
Tomato(Lycopersicon esculentum Mill.);Inheritance of fruit color;Inheritance of pigment content;Multiple generation conjoint analysis
吴浪,男,硕士研究生,专业方向:蔬菜育种与生物技术,E-mail:15291832633@163.com
*通讯作者(Corresponding author):梁燕,教授,博士生导师,专业方向:番茄新品种选育与蔬菜种质资源创新,E-mail:liangyan@nwsuaf. edu.cn
2016-10-13;接受日期:2017-01-07
国家重点研发计划项目(2016YFD0101703,2016YFD01002 04-30),陕西省科技统筹创新工程项目(2015KTTSNY03-01,2015K TCG01-04)
猜你喜欢
娃娃乐园·综合智能(2022年9期)2022-08-16
湖南农业(2022年6期)2022-07-29
食品安全导刊(2021年20期)2021-08-30
科学大众(2021年9期)2021-07-16
临床肝胆病杂志(2020年1期)2020-12-20
中国食品学报(2020年7期)2020-08-03
消费导刊(2017年24期)2018-01-31
中国调味品(2017年2期)2017-03-20
中国科技纵横(2016年15期)2016-12-29
甘肃教育(2016年22期)2016-12-20
