
低氧训练对大鼠骨骼肌谷胱甘肽抗氧化系统的影响
2017-05-08 03:22陈晓彬林文翁锡全
军事体育学报 2017年4期
陈晓彬,林文,翁锡全
(广州体育学院 运动生物化学省重点实验室,广东 广州 510075)
近年来,高原训练作为一种提高有氧耐力辅助训练手段引起国内外体育界重视,国内外许多有氧耐力项目运动员在参加重大比赛前经常采用了高原训练手段。自由基代谢及抗氧化酶与机体的机能状态密切相关,低氧训练对人体自由基抗氧化系统影响一直备受人们关注。谷胱甘肽抗氧化系统是机体内非常重要的抗氧化酶系,最近研究报道,GSH-PX和GSH基因表达是低氧敏感基因之一[1,2]。目前,国内外有关低氧训练的研究结果仍存在许多分歧和争议,有关低氧训练对谷胱甘肽抗氧化系统这方面的研究尚较少,本文通过探讨低氧训练对骨骼肌谷胱甘肽抗氧化系统的影响,为低氧训练方法进一步推广提供理论依据。
1 材料与方法
1.1 实验对象
雄性SD大鼠40只,体重190~210 g,8 wk龄,购自中山大学实验动物中心。分笼饲养,每笼5只,自由饮食、饮水,标准啮齿类饲料,室温(25±2)℃,湿度40%~60%,自然采光,利用空调和排气扇保持通风,每天换一次木屑垫料以保持清洁。
1.2 实验分组
将雄性SD大鼠40只随机分为低住安静组(LC)、低住低练组(LoLo)、低住高练组(LoHi)和高住低练组(HiLo)等4组,每组10只。
1.3 实验方案
实验采用美国Hypoxico公司生产常压低氧舱,低氧浓度控制在15.4%左右。低住安静组大鼠每天在常氧环境生活;低住低练组大鼠每天在常氧环境生活23 h,常氧动物跑台运动1 h;低住高练组大鼠每天在常氧环境生活23 h,低氧动物跑台运动1 h;高住低练组大鼠每天在低氧环境生活23 h,常氧动物跑台运动1 h。各运动组大鼠采用杭州段氏生产的动物跑台进行运动,低住组运动强度为35 m/min、高住组运动强度为30 m/min,60 min/d,5 d/wk,持续4 wk。
1.4 样本采集和处理
第4周末次训练后所有大鼠禁食12 h进行采样,先按大鼠0.3 ml/100 g体重剂量腹腔注射10%水合三氯乙醛溶液进行麻醉、固定,迅速分离大鼠股四头肌,用生理盐水漂洗大鼠股四头肌表面的血迹,采用滤纸吸干,用灭菌锡纸包好后放在液氮罐里面速冻,后移放至-70℃冰箱冻存备用。
1.5 测试方法
谷胱甘肽转硫酶(GST)、谷胱甘肽过氧化物酶(GSH-PX)、还原型谷胱甘肽(GSH)、谷胱甘肽还原酶(GR)、丙二醛(MDA)、总抗(T-AOC)、巯基(-SH)等生化指标测试均采用南京建成生物工程研究所提供试剂盒进行测定,专人负责各生化指标的测试,测试时严格按照说明书要求进行操作。
1.6 主要仪器
S22PC分光光度计、752紫外光分光光度计、内切式组织匀浆机、高速冷冻离心机、恒温箱、三合电热水箱等。
1.7 数据处理
2 实验结果
2.1 各组大鼠股四头肌谷胱甘肽抗氧化系统指标的变化
从表1可见,低住高练组和高住低练组大鼠股四头肌GSH活性显著高于其它各组(P<0.05),但两组间差异不显著(P>0.05),低住低练组大鼠股四头肌GSH活性显著高于低住安静组(P<0.05);低住高练组和高住低练组大鼠股四头肌GSH-PX活性显著高于其它各组(P<0.05),但两组间差异不显著(P>0.05),低住低练组大鼠股四头肌GSH-PX活性显著高于低住安静组(P<0.05);低住安静组大鼠股四头肌GST活性显著低于其它各组(P<0.05),其余各组间大鼠股四头肌GST活性没有显著性差异(P>0.05);低住低练组大鼠股四头肌GR活性显著高于其它各组(P<0.05),其余各组间大鼠股四头肌GR活性没有显著性差异(P>0.05)。
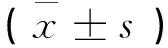
表1 各组大鼠股四头肌谷胱甘肽抗氧化系统指标的变化
2.2 各组大鼠肌四头肌总抗、丙二醛、巯基的变化
从表2可见,低住高练组和高住低练组大鼠股四头肌T-AOC显著高于其它各组(P<0.05),但两组间差异不显著(P>0.05),低住低练组大鼠股四头肌T-AOC显著高于低住安静组(P<0.05);低住高练组和高住低练组大鼠股四头肌MDA显著低于其它各组(P<0.05),但两组间差异不显著(P>0.05),低住低练组大鼠股四头肌MDA显著低于低住安静组(P<0.05);低住高练组、高住低练组、低住低练组各组间大鼠股四头肌-SH的活性比较,差异不显著(P>0.05)。
表2 各组大鼠股四头肌总抗、丙二醛、巯基的变化
3 分析与讨论
谷胱甘肽抗氧化系统主要指谷GSH、GSSG、GSH-PX、GR、GST。GSH主要生理功能是清除羟自由基,防止自由基损伤,同时保护细胞膜中巯基蛋白质和酶的还原状态,也可以作为GSH-PX、GST底物与各种内源性和外源性电子化合物发生反应,生成无毒性或毒性小的硫结合物,而自身被氧化生成GSSG,GSSG在GR催化下,以NADPH为供氢体,又可还原生成GSH。
国内外有关低氧训练对抗氧化酶的研究结果仍存在一些分歧和争议,其中有一些研究文献并不支持低氧训练能提高机体的抗氧化水平,这可能和实验方案中的低氧浓度以及实验方法有关。如Hartney[3]等研究发现低氧训练会导致GSH和GSSG的比值降低,这不利于体内自由基的清除。Rulin Li等[4]对怀孕大鼠进行极低氧浓度(2%~5%O2)的低氧刺激,结果发现与对照组大鼠相比,实验组大鼠GSH显著降低,脂质过氧化水平严重。但也研究报道,进行一定程度的低氧训练能使相关细胞的GSH-PX抗氧化酶基因的表达量增加[5]。Jain研究也表明,进行适度的低氧训练可以提高谷胱甘肽过氧化物酶的活性[6]。Badawi[7](2009)等在研究低氧暴露环境下HIF-1对星形胶质细胞对咳肮贡酸毒性的研究中也发现,进行低氧暴露能导致细胞的GSH水平的升高。
本研究发现,低住高练组和高住低练组大鼠股四头肌除GR活性无明显变化外,GSH、GSH-Px、GST均得到了显著的改善,如低住高练组和高住低练组大鼠股四头肌GSH、GSH-PX显著高于其它各组,且低住低练组显著高于低住安静组。这表明进行低住高练和低住高练均能使大鼠股四头肌谷胱甘肽抗氧化能力得到改善。分析其内在机制,这可能是当机体处于适度的低氧刺激时,能在避免引起机体损伤的前提下通过生物体的适应能力,增强机体的抗氧化能力。而当低氧浓度极低甚至可以说时亚弱的条件下会造成机体不良反应,使得机体产生大量ROS,易引起机体损伤的应激[8,9]。而机体产生大量的ROS和其它对机体产生损害的自由基的同时,一方面,机体GSH含量却显著下降,GSSG含量升高,使得机体整体的抗氧化能力大大降低;另一方面,与谷胱甘肽抗氧化能力相关的酶如GSH-PX、GST、GSH的总体变化趋势也会朝着不利于机体抗氧化能力的方面转变。在本研究中,低住高练与高住低练组的大鼠骨四头肌GST、GR与低住低练组无明显差异,但是低住安静组要显著低于低住高练与高住低练组,而低住高练与高住低练组的巯基要显著高于低住安静组与低住低练组,这可能和相关的酶对GSH和GSSG之间浓度变化的适应性提高有关。
正常情况下ROS的产生和清除处于动态平衡之中,当ROS产生大于清除时,即会攻击机体,即产生所谓氧化应激。ROS能够与生物膜脂质、蛋白质、糖类等物质发生反应,组织的脂质过氧化是ROS过量引起氧化损伤的重要标志。ROS通过其强烈的氧化作用,对不饱和脂肪酸产生氧化和过氧化作用,生成脂质过氧化物,并降解成MDA,MDA的寿命比羟基自由基长,具有很强的毒性,极易与磷脂蛋白发生反应,改变细胞膜的通透性,而造成组织细胞的氧化损伤,可以由生成部位扩散到其他部位而产生毒性作用,也可以引起其它物质产生过氧化作用的连锁反应,所以MDA是反映LPO水平的有效指标。
本实验研究发现,低住低练组大鼠股四头肌MDA显著低于低住安静组,低住低练组大鼠股四头肌T-AOC显著高于低住安静组,说明4 wk的跑台运动可以使脂质过氧化物生成减少,降低组织细胞损伤。大量的研究表明,低氧训练提高了机体对缺氧的适应能力,减少MDA生成,增强机体的抗氧化能力[10,11]。本实验也发现相类似的结果,低住高练组和高住低练组大鼠股四头肌MDA显著低于其它各组,低住高练组和高住低练组大鼠股四头肌T-AOC显著高于其它各组,这表明低氧训练可以减少脂质过氧化物的生成,降低对组织细胞的损伤,同时也说明机体对ROS产生了适应,加上适宜的运动训练更有利于机体抗缺氧耐受性的形成。但也有实验结果发现低氧下自由基过量产生,自由基引起的脂质过氧化使细胞膜结构和功能都受到损伤。Sarada研究发现低氧暴露导致血浆中MDA水平增加,同时血液中GSH,GSH-PX减少[12]。造成这两种截然不同的结果可能是低氧刺激的浓度、低氧刺激时间、以及不同组织器官等。
4 结论
运动、低住高练和高住低练均能提高大鼠股四头肌GSH、GSH-PX、T-AOC、减少MDA的生成,这表明低住高练与高住低练这两种训练方法比单一的常氧运动更能改善机体的抗氧化能力,降低脂质过氧化水平。
参考文献:
[1] Ornoy A, Livshitz A, Ergaz Z,et al. Hyperglycemia, hypoxia and their combination exert oxidative stress and changes in antioxidant gene expression: studies on cultured rat embryos[J].Birth Defects Res B Dev Reprod Toxicol,2011,92(3):231—239.
[2] Stefan Schülkea, Daniel Dreidaxa, Assaf Malik,et al. Living with stress: Regulation of antioxidant defense genes in the subterranean, hypoxia-tolerant mole rat, Spalax[J].Gene,2012,500(2):199—206.
[3] Tanya Hartney, Rahul Birari, Sujatha Venkataraman,et al. Xanthine Oxidase-Derived ROS Upregulate Egr-1 via ERK1/2 in PA Smooth Muscle Cells; Model to Test Impact of Extracellular ROS in Chronic Hypoxia[J].PLoS ONE,2011,6(11):e27531—e27543.
[4] Rulin Li,Martha Chase,Sung-Kwon Jung.Hypoxic stress in diabetic pregnancy contributes to impaired embryo gene expression and defective development by inducing oxidative stress[J].Am J Physiol Endocrinol Metab.2005,289:591—559.
[5] Ainiwaer Aikemu,Abdiryim Yusup, Anwar Umar,et al.The impact of the Uighur medicine abnormal savda munziq on antitumor and antioxidant activity in a S180 and Ehrlich ascites carcinoma mouse tumor model[J]. Pharmacogn Mag.2012,8(30):141—148.
[6] Vishal Jain,Iswar Baitharu,Kalpana Barhwal.Enriched Environment Prevents Hypobaric Hypoxia Induced Neurodegeneration and is Independent of Antioxidant Signaling[J].Cell Mol Neurobiol,2012,32:599—611.
[7] Yomna Badawi, Prabhu Ramamoorthy,Honglian Shi.Hypoxia-inducible factor 1 protects hypoxic astrocytes against glutamate toxicity[J].ASN NEURO.2012,4(4):231—241.
[8] Guofei Zhou,Laura A. Dada,Minghua Wu,et al.Hypoxia-induced alveolar epithelial mesenchymal transition requires mitochondrial ROS and hypoxia-inducible factor 1[J].Am J Physiol Lung Cell Mol Physiol,2009,297:L1120—L1130.
[9] Annie Y. Chi, Gregory B. Waypa, Paul T,et al.Prolonged Hypoxia Increases ROS Signaling and RhoA Activation in Pulmonary Artery Smooth Muscle and Endothelial Cells[J].Antioxid. Redox Signal,2010,12(5):603—610.
[10] Cell BR. Is pulmonary rehabilitation an effective treatment for chronic obstructive pulmonary disease Am J Respire Cirt Care, 1997, 155(5):781—783.
[12] Sarada, S.K.S. sarada, Sairam, M, Dipti, P et al. Role of selenium in reducing hypoxia-induced oxidative stress: an in vivo study. Biomedicine & Pharmacotherapy; 2002, 56(4):173—176.
猜你喜欢
山东第一医科大学(山东省医学科学院)学报(2022年8期)2022-12-07
浙江大学学报(农业与生命科学版)(2022年4期)2022-09-07
中国油脂(2022年5期)2022-05-31
现代临床医学(2021年6期)2021-11-20
现代养生·上半月(2021年10期)2021-09-24
中国畜牧杂志(2021年6期)2021-06-19
体育科学(2021年12期)2021-03-23
昆明医科大学学报(2021年1期)2021-02-07
分析化学(2018年4期)2018-11-02
安徽化工(2015年3期)2015-12-12
