
115份高羊茅种质资源农艺性状特征分析
2017-03-28 12:15娄燕宏宋付朋诸葛玉平徐庆国
草原与草坪 2017年1期
娄燕宏,王 会,宋付朋,诸葛玉平,徐庆国
(1.山东农业大学 资源与环境学院,山东 泰安 271018; 2.湖南农业大学 农学院,湖南 长沙 410128)
115份高羊茅种质资源农艺性状特征分析
娄燕宏1,王 会1,宋付朋1,诸葛玉平1,徐庆国2
(1.山东农业大学 资源与环境学院,山东 泰安 271018; 2.湖南农业大学 农学院,湖南 长沙 410128)
对不同地理来源的115份高羊茅种质资源的株高、穗长、叶枕距、每穗小穗数、主穗分枝数、单株穗数、穗重等7个农艺性状进行了主成分和聚类分析。结果表明:前3个主成分代表了115份高羊茅种质7个农艺性状的82.82%的信息,且对3个主成分作用较大的性状主要有穗长、单株穗数、主穗分枝数。应用系统聚类分析,将115份高羊茅种质分为5个类群,其中,第Ⅱ类群又可分为3个亚类群,其中Ⅰ-Ⅱ亚类群和Ⅳ类群的每穗小穗数和穗重较大,综合表现较好。同时,地理距离和农艺性状间未存在严格的一致性。
高羊茅;农艺性状;主成分分析;聚类分析
高羊茅(Festucaarundinacea),又称苇状羊茅,不仅是一种常用的草坪草,而且是一种重要的牧草[1]。高羊茅具有抗旱、耐湿、抗寒、耐热、耐贫瘠等特性,广泛分布于欧洲和北非、中东、中亚和西伯利亚的地中海地带,并延伸分布于西伯利亚、东非及马达加斯加地区,在我国新疆、东北、贵州、四川、广西等湿润地区亦有分布[2]。
1 材料和方法
1.1 试验地概况
试验地位于湖北省武汉市武昌区东湖磨山中国科学院武汉植物园草坪种质资源圃内,土壤为黄棕壤土,质地为粘土,土壤肥力和水分状况良好,雨量充沛,日照充足,夏季酷热,冬季湿冷,属亚热带过渡的湿润季风性气候。年平均气温15.8~17.5℃,最冷月(1月)均温3.0℃;最热月(7月)均温29.3℃;极端最高气温44.5℃,活动积温在5 000℃~5 300℃;年降水量1 205 mm,年蒸发量1 455 mm,年日照时数2 000 h,无霜期达240 d,试验期间无极端异常天气出现。
1.2 供试材料
供试材料选用来自世界不同国家和地区的115份高羊茅种质材料,其中25份为商业品种,均来自于美国,其余90份野生型高羊茅种质材料由美国农业部(United States of Agriculture-Agricultural Research Service,USDA-ARS)提供,依据其不同地理来源划分为4个群体(美洲群体、欧洲群体、非洲群体、亚洲群体)(表1)。
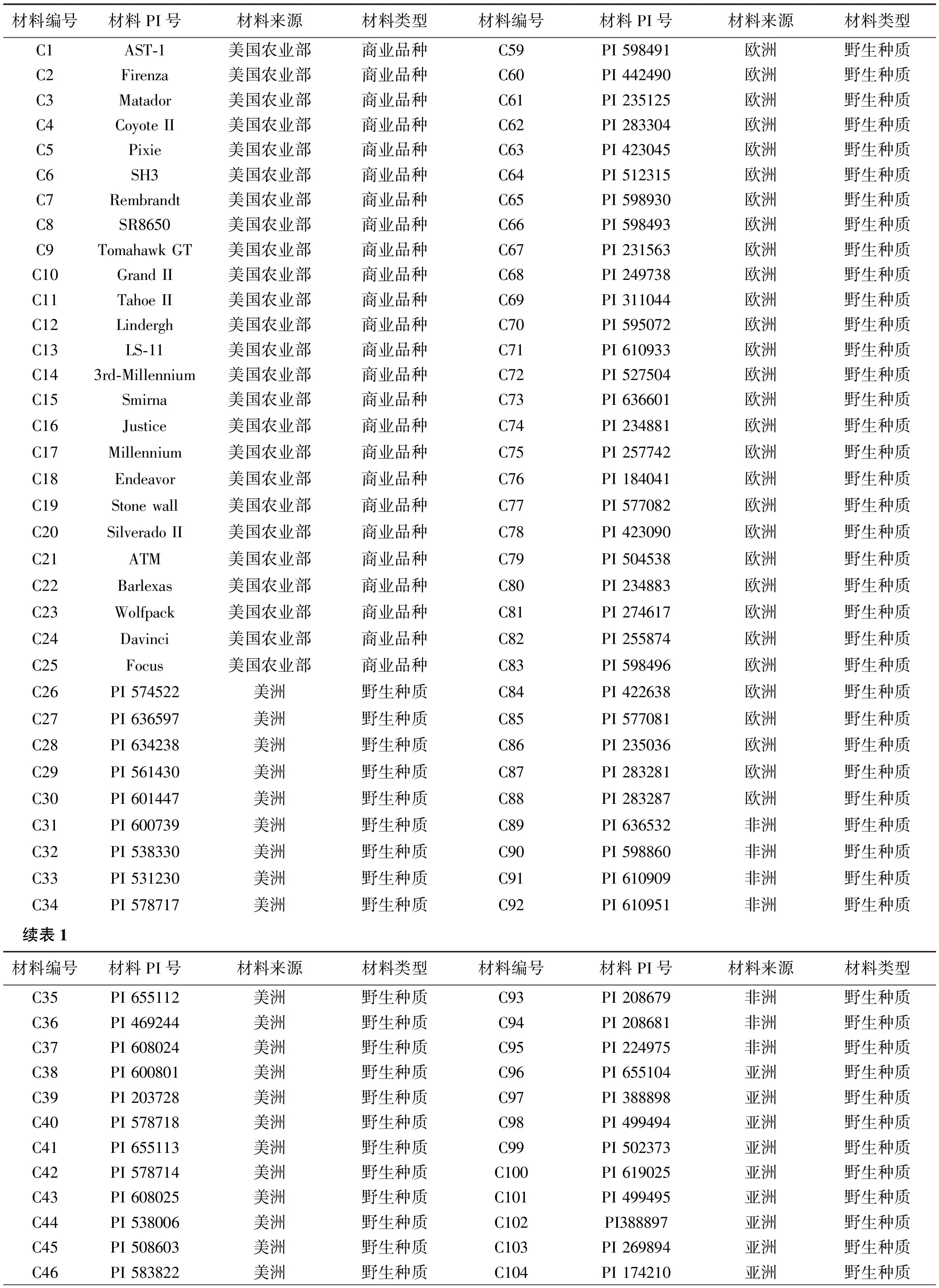
表1 供试115份高羊茅种质资源的编号及来源
1.3 田间试验方法
2012年5月,从每份高羊茅种质中选取10粒饱满的种子,置于铺有湿滤纸的培养皿中,将培养皿于20℃下避光培养,每天补充水分以保证滤纸湿润,以提供种子萌发所需要的水分。待种子发芽后,提供自然光照,光强为300~500 μmol photons/(m2·s),光周期为14 h。培养两周后,所有种质随机选择优良单株转移至装有有机营养土与沙子(v/v=1∶1)混合栽培基质的塑料花盆(直径14 cm,高度15 cm)中进行分蘖增殖,多次进行分蘖增殖以保证材料的遗传一致性。所有试验材料置于中国科学院武汉植物园可控温室中培养,温度设定为白天24℃,夜间20℃,光周期14 h,光强 300 μmol photons/(m2·s)。每天浇水以保证高羊茅生长的水分需求,每周浇1/2 Hoagland营养液(所有营养成分均用去离子水制备)1次以保证高羊茅生长的养分供应。每周剪草1次,剪草高度为7 cm。
2012年9月,试验地喷施灭生性除草剂草甘膦杀死杂草并将杂草清理干净,撒施底肥N 49 kg/hm2,P 98 kg/hm2,K 98 kg/hm2,覆盖2 cm厚有机土,然后深翻土壤,耙平,最后覆沙2 cm。2012年10月,将115份高羊茅种质材料转移至田间,小区面积为1.5 m×1.5 m,小区间隔0.5 m,随机区组排列,3次重复。2013年3月施用复合肥(N∶P∶K=21∶6∶13)1次,用量以N 49 kg/hm2计。2013年4~11月,每月追肥1次,复合肥与尿素交替使用。2013年6月13日数据采集完成后,剪草至草高10 cm,在整个试验期间其他时间不再进行剪草。2014年,分别于4月、5月各施肥1次,4月施用复合肥,5月施用尿素,施用量以N 49 kg/hm2计。2014年5月28日~6月7日田间采集数据。整个试验期间保持充足的水分供应,灌溉量以湿润整个根系为宜。
1.4 测定项目及方法
不同开口形状与开口不同的加强形式的应力集中程度如图7所示,开口位置的主要载荷形式为拉压载荷,说明风机塔筒结构主要受弯曲载荷作用。从图7可以看出,在孔边缘焊接加强结构会大幅度降低开孔边缘的应力峰值,但是对开口结构的疲劳强度却未必有改善。不同开口的应力集中情况与疲劳损伤大小如表5所示,从表中可以看出,虽然增加套筒降低了开孔边缘的应力集中,但是疲劳寿命却降低了,主要原因是引入了焊接,增加了开口结构疲劳开裂的风险。所以一旦出现疲劳强度不足时,除非考虑优化开口形式,很难找到其他更加经济的方法,所以在单桩基础上开口,需要对疲劳强度做细致的分析,确保开口结构具有足够的疲劳强度。
分别于2013年6月4日~13日,2014年5月28~6月7日,每个小区随机选择10株高羊茅,测量最高部位至地面的绝对高度,取平均值即为株高;测量穗顶端至旗叶枕的距离,取平均值即为(旗叶)叶枕距;测量穗顶端至穗颈节的长度,取平均值即为穗长。对选取的10株高羊茅进行考种,记录每株的穗数、每穗的分枝数、每穗的小穗数和每穗重量,取平均值即为单株穗数、主穗分枝数、每穗小穗数和穗重。
1.5 数据统计与分析
各高羊茅种质农艺性状的数据采用两年调查数据的平均值,用Excel 2003进行数据统计,采用SPSS 13.0软件进行主成分分析和聚类分析,根据农艺性状间的遗传相关系数矩阵计算特征根、特征向量;聚类分析采用欧式距离法。
2 结果与分析
2.1 主成分分析
通过对不同地理来源的115份高羊茅种质资源的主要农艺性状进行主成分分析,结果表明,试验所选用的7个农艺性状中的前3个主成分累计贡献率达到82.82%,能概括7个主要农艺性状的绝大部分信息,并能基于此信息对这些种质资源进行综合评价(表2)。
第1主成分的特征值为3.106,贡献率为44.366%,其中穗长对应的特征向量最高;其次为叶枕距和株高,主要反映穗长、叶枕距和株高的影响。
第2主成分的特征值为1.601,贡献率为22.872%,其中单株穗数对应的特征向量最高;其次为穗重,主要反映单株穗数和穗重的影响。
第3主成分的特征值为1.090,贡献率为15.578%,主穗分枝数对应的特征向量最高;其次为每穗小穗数、穗重,主要反映主穗分枝数、每穗小穗数、穗重的影响。
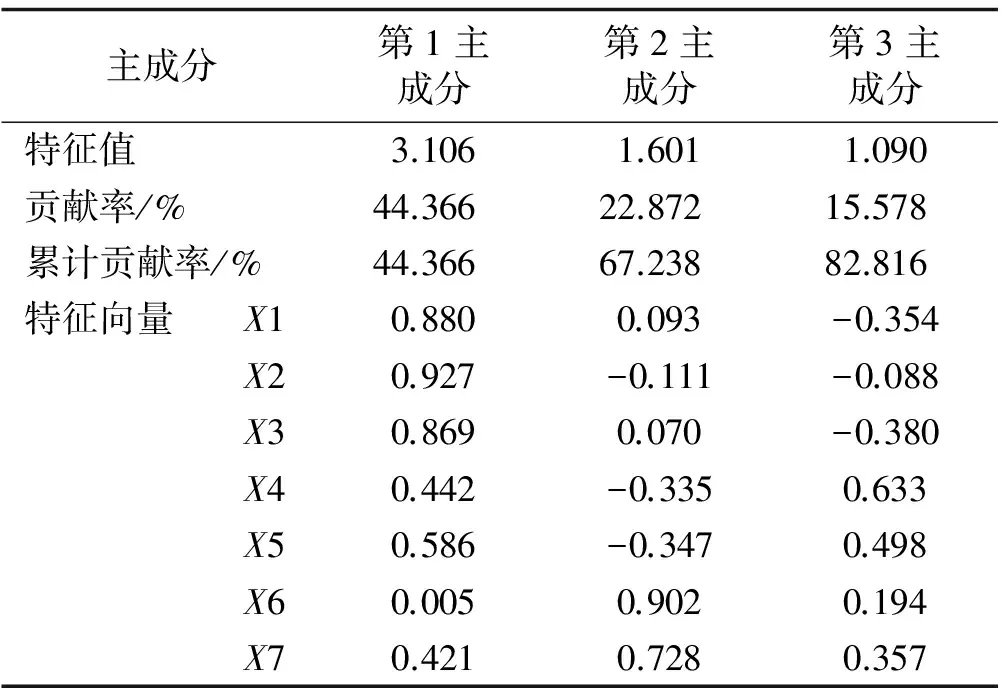
表2 不同地理来源高羊茅种质主要农艺性状的主成分分析
注:X1 株高;X2 穗长;X3 叶枕距;X4 每穗小穗数;X5 主穗分枝数;X6 单株穗数;X7 穗重
2.2 聚类分析
运用软件DPS 3.01系统进行聚类分析,以欧式距离作为各高羊茅种质间距离,以类平均法为聚类方法,以各高羊茅株高、穗长、叶枕距、每穗小穗数、单株穗数、主穗分枝数、穗重等7个主要农艺性状为指标,对115份高羊茅种质资源进行了聚类分析,建立种质资源聚类树状图,115份高羊茅种质资源可分为5大类群(图1)。
第Ⅰ类群包括26份种质资源(10份商业品种,5份材料来自于美洲群体,7份材料来自于欧洲群体,4份材料来自于亚洲群体)。他们特点是株高在69.6~127.6 cm;穗长在13.0~34.6 cm;叶枕距在29.5~68.1 cm;每穗小穗数在42.2~108.4粒/穗;主穗分枝数在9.0~16.3个/穗;单株穗数在6.0~44.5穗/株;穗重在3.9~30.3 g/穗。其类群材料具有单株穗数和穗重较小的特征。
第Ⅱ类群包括53份种质资源(3份商业品种,18份材料来自于美洲群体,17份材料来自于欧洲群体,6份材料来自于非洲群体,9份材料来自于亚洲群体),又可分为3个亚类群,第Ⅰ-Ⅱ亚类群包含16份种质资源(3份商业品种,8份材料来自于美洲群体,3份材料来自于欧洲群体,2份材料来自于非洲群体)。主要特点是株高在114.1~158.8 cm,穗长在23.2~38.0 cm;叶枕距在57.1~83.4 cm,每穗小穗数在98.8~140.8粒/穗,主穗分枝数在11.8~16.6个/穗,单株穗数在40.2~78.3穗/株,穗重在22.2~73.6 g/穗。本类群材料具有株高、叶枕距、每穗小穗数和穗重较大的特征。第Ⅱ-Ⅱ亚类群包含26份种质资源(9份材料来自于美洲群体,6份材料来自于欧洲群体,4份材料来自于非洲群体,7份材料来自于亚洲群体)。主要特点是株高在129.8~176.1 cm,穗长在26.5~40.3 cm,叶枕距在58.5~88.8 cm,每穗小穗数在57.2~112.8粒/穗,主穗分枝数在10.7~14.0个/穗,单株穗数在11.2~66.8穗/株,穗重在13.9~45.4 g/穗。本类群材料具有株高、穗长和叶枕距较大,单株穗数较小的特征。第Ⅱ-Ⅲ亚类群包含11份种质资源(自于美洲群体的C40,来自亚洲群体的C96和C108,8份材料来自于欧洲群体)。主要特点是株高在107.4~145.5 cm,穗长在29.2~44.2 cm,叶枕距在48.7~77.6 cm,每穗小穗数在88.2~123.4粒/穗,主穗分枝在11.4~16.6个/穗,单株穗数在16.0~37.0穗/株,穗重在8.3~49.0 g/穗。本类群材料具有穗长、叶枕距和主穗分枝数较大,单株穗数较小的特征。
第Ⅲ类群包括25份种质资源(11份商业品种,7份材料来自于美洲群体,6份材料来自于欧洲群体,1份材料来自于非洲群体)。主要特点是株高在95.5~169.0 cm,穗长在20.2~37.0 cm,叶枕距在44.6~76.1 cm,每穗小穗数在36.9~98.3粒/穗,主穗分枝数9.0~15.6个/穗,单株穗数在55.7~137.2穗/株,穗重在21.3~76.5 g/穗。本类群材料具有叶枕距、单株穗数和穗重较大,每穗小穗数和主穗分枝数较小的特征。
第Ⅳ类群仅有3份种质材料,分别是来自欧洲群体的C69,来自亚洲群体的C97和C100。主要特点是株高在115.2~129.6 cm,穗长在31.3~39.2 cm,叶枕距在40.7~69.4 cm,每穗小穗数在143.5~183.6粒/穗,主穗分枝数在12.4~18.8个/穗,单株穗数在9.7~40.0穗/株,穗重在15.1~26.0 g/穗。本类群材料具有穗长、每穗小穗数和主穗分枝数较大,单株穗数较小的特征。
第Ⅴ类群包括8份种质资源(商业品种C9,来自美洲群体的C39,来自欧洲群体的C61,还有5份种质材料自来于亚洲群体)。主要特点是株高在34.3~131.1 cm,穗长在21.6~37.3 cm,叶枕距在24.0~61.5 cm,本类群具有穗长较大的特征。
将各高羊茅种质聚类分析结果与高羊茅的地理来源相比较,分析看出各高羊茅种质商业品种主要分布于第I类群(10份种质材料,占商业品种材料总数的40.0%)和第Ⅲ类群(11份种质材料,占商业品种材料总数的44.0%),只有3份高羊茅商业品种种质分布于第Ⅱ类群,仅有1份高羊茅商业品种种质分布于第Ⅴ类群;高羊茅来自于美洲的种质主要分布于第Ⅱ类群(18份材料,占美洲群体材料总数的58.1%)、第Ⅲ类群(7份材料)和第I类群(5份材料),仅有1份高羊茅美洲群体的材料分布于第Ⅴ群体;高羊茅来自于欧洲的种质主要分布于第Ⅱ类群(17份材料,占欧洲群体材料总数的53.1%)、第I类群(7份材料,占欧洲群体材料总数的21.9%)和第III类群(6份材料,占欧洲群体材料总数的18.8%),另外各有1份高羊茅欧洲群体材料分布于第Ⅳ类群和第V类群;高羊茅来自于非洲的种质主要分布于第Ⅱ类群(6份材料,占非洲群体材料总数的85.7%)和第Ⅲ类群(1份种质材料);高羊茅来自于亚洲的种质主要分布在第II类群(9份材料,占亚洲群体材料总数的45.0%)、第Ⅴ类群(5份材料,占亚洲群体材料总数的25.0%)和第I群体(4份材料,占亚洲群体材料总数的20.0%),另外2份高羊茅亚洲群体材料分布于第Ⅳ群体。因此,地理距离和农艺性状间未存在严格的一致性。
3 讨论与结论
种质资源是作物遗传改良的重要基因来源,种质资源的遗传多样性研究可为作物遗传改良提供理论依据,作物育种史上的重大突破均与优异种质的发现和利用相关[14-16]。因此,对高羊茅种质资源农艺性状的研究,发掘其优异的基因资源,对研究高羊茅的遗传多样性、种质资源、遗传改良均具有重要意义。研究选用的115份高羊茅种质具有不同的地理来源,其中90份为野生种质,这些种质具有丰富的优异基因,不管是在株型育种、远缘杂种优势利用,还是抗性育种等方面都具有巨大潜力[17]。研究表明,115份高羊茅种质存在广泛的遗传多样性,主要农艺性状的变异系数由大到小排列为单株穗数>穗重>每穗小穗数>叶枕距>株高>穗长>主穗分支数。高羊茅单株穗数和穗重的变异系数最大,分别为55.68%和48.28%,其遗传变异极其丰富,可选择范围大,农艺性状遗传所控制的变异潜力大,可根据其表型进行有效选择;而其主穗分枝数的变异系数较小,遗传变异较窄,遗传选择效果不大[18]。
作物种质资源的综合评价中,对多个性状的综合考察因性状间存在的相关性,造成一定信息的重复,进而降低了综合评价法的准确性[19]。主成分分析是一个降维过程,避免信息重叠,为资源的评价和选择提供了更为科学的依据[20]。通过对不同地理来源的高羊茅种质资源的主要农艺性状的主成分分析表明,前3个主成分代表了115份高羊茅种质7个农艺性状的82.82%的信息。对3个主成分作用较大的性状主要有穗长、单株穗数、主穗分枝数,表明这3个性状可能是影响高羊茅表型的主要农艺性状。同时,采用该研究确定的3个综合因子替代原有的7个农艺性状变量,简化了高羊茅种质资源综合评价的数据,并且还揭示了高羊茅不同农艺性状变量间的关系,为高羊茅育种的亲本选配提供了有力的依据[17]。

图1 高羊茅种质资源各农艺性状的聚类图Fig.1 Cluster analysis of agronomic traits of tested accessions
聚类分析作为一种研究作物种质资源差异分类的切实可行的分析方法,能粗略地反映出种质间的亲缘关系[21]。对115份高羊茅种质材料的聚类分析结果表明,不同高羊茅种质可划分为5个类群,第I类群26份种质的单株穗数和穗重较小。第Ⅱ类群包括53份种质又可分为3个亚类群,第Ⅰ-Ⅱ亚类群16份种质的株高、叶枕距、每穗小穗数和穗重较大;第Ⅱ-Ⅱ亚类群26份种质的株高、穗长和叶枕距较大,单株穗数较小;第Ⅱ-Ⅲ亚类群11份种质的穗长、叶枕距和主穗分枝数较大,单株穗数较小。第III类群25份种质的叶枕距、单株穗数和穗重较大,每穗小穗数和主穗分枝数较小。第Ⅳ类群3份种质的穗长、每穗小穗数和主穗分枝数较大,单株穗数较小。第Ⅴ类群8份种质的穗长较大。因此,可依据高羊茅育种目标并根据不同高羊茅种质类群农艺性状的遗传变异特点,选配适当的高羊茅种质类群亲本材料。
此次研究仅对不同地理来源的115份高羊茅种质资源的主要农艺性状进行了分析评价,若要全面、客观的评价高羊茅种质资源,还需进一步开展品质分析、抗逆性鉴定,同时研究其遗传特性。
[1] 倪星,秦楚,平玲,等.转SOS基因高羊茅的耐盐性鉴定[J].草原与草坪,2016,36(1):43-47.
[2] 周寿荣.草坪地被与人类环境[M].成都:四川科学技术出版社,1996.
[3] 王建丽,申忠宝,潘多峰,等.红三叶种质资源农艺性状的综合评价及聚类分析[J].草原与草坪,2013,33(2):33-37.
[4] 王林海,王晓伟,詹克慧,等.黄淮麦区部分小麦种质资源农艺性状的聚类分析[J].农艺科学,2008,24(4):186-191.
[5] 郝峰,徐柱,李平,等.无芒雀麦农艺性状遗传多样性研究[J].草业科学,2011,28(5):769-776.
[6] 胡兴雨,陆平,贺建波,等.黍稷农艺性状的主成分分析与聚类分析[J].植物遗传资源学报,2008,9(4):492-496.
[7] Zarrough K M,Nelson C J,Coutts J H.Relationship between tillering and forage yield of tall fescue.I.Yield[J].Crop Science,1983,23(2):333-337.
[8] Nguyen H T,Sleper D A.Genetic variability of seed yield and reproductive characters in tall fescue[J].Crop Sci,1983,23(4):621-626.
[9] 李德颖,孙阿琦.高羊茅生物学性状分析在草坪品种选育中的应用[J].草业学报,1997,6(2):28-35.
[10] Hopkins A A,Young C A,Panaccione D G,etal.Agronomic performance and lamb health among several tall fescue novel endophyte combinations in the south-central USA[J].Crop Sci,2010,50(4):1552-1561.
[11] 吴佳海,尚以顺,牟琼.高羊茅种子产量与主要农艺性状的通经分析[J].种子,2000(5):64-65.
[12] Wang Q H,Dooner H.Remarkable variation in maize genome structure inferred from haplotype diversity at the bz locus[J].Proc Natl Acad Sci USA,1996,103(47):17644-17649.
[13] 左相兵,付薇,杨正德,等.贵州饲用燕麦种质资源农艺性状的遗传多样性分析[J].贵州农业科学,2012,40(6):9-13.
[14] Walsh J.Genetic vulnerability down on the farm[J].Science,1981,214(4517):161-164
[15] 张新全,张锦华,杨春华,等.四川省牧草种质资源现状及育种利用[J].四川草原,2002(1):6-15.
[16] 李春花,王艳青,卢文洁,等.云南薏苡种质资源农艺性状的主成分和聚类分析[J].植物遗传资源学报,2015,16(2):277-281.
[17] 董攀,李伟,郑有良.波兰小麦主要农艺性状分析[J].麦类作物学报,2007,27(2):216-222.
[18] Lou Y H,Chen L,Xu Q G,etal.Genotypic Variation of Morphological Traits in Tall Fescue(FestucaarundinaceaSchreb.) Accessions[J].HortScience.2015,50(4):512-516.
[19] 李新峥,杜晓华,张振伟.中国南瓜经济性状遗传初探[J].西北农业学报,2009,18(4):319-323
[20] 杜晓华,李新峥,刘海妮.南瓜前期农艺性状的主成分分析[J].西北农业学报,2010,19(2):168-171.
[21] 王绘艳,史雨刚,马昊翔,等.30份春小麦品系主要农艺性状的聚类分析[J].中国农学通报,2014,30(18):60-64.
Agronomical traits of 115 tall fescue (Festucaarundinacea) accessions
LOU Yan-hong1,WANG Hui1,SONG Fu-peng1,ZHGE Yu-ping1,XU Qing-guo2
(1.CollegeofResourcesandEnvironment,ShandongAgriculturalUniversity,Taian410128,China;2.CollegeofAgronomy,HunanAgriculturalUniversity,Changsha410128,China)
The agronomical traits (plant height,spike length,pulvinus distance,spikelet count,branch count per spike,spike count per plant,and spike weight) of 115 tall fescue accessions were observed and analyzed through principle component and cluster analysis in order to identify the diversity of tall fescue germplasm.The results showed that 7 traits could be simplified into 3 principle components (over 82.82% contribution),and the contributions of spike length,branch count per spike and spike count per plant were higher.The results of cluster analysis indicated that all accessions could be classified into 5 clusters,and 3 sub-clusters were divided in Ⅱ cluster.The higher spikelet count and spike weight were observed in Ⅱ-Ⅰ and Ⅳ cluster.Clustering analysis showed that no significant correlation between geographic distance and agronomic traits.
tall fescue;agronomical traits;principle component analysis;cluster analysis
2016-05-23;
2016-06-06
山东省自主创新及成果转化专项(2014 ZZCX07402);湖南省教育厅项目(KC2011B023)资助
娄燕宏(1985-),山东蒙阴人,博士,讲师,研究方向为草类植物逆境生理及遗传育种。 E-mail:yanhonglou@sdau.edu.cn 诸葛玉平,徐庆国为通讯作者。
S 543
A
1009-5500(2017)01-0031-08
猜你喜欢
新农民(2021年15期)2021-09-16
湖南农业大学学报(自然科学版)(2021年2期)2021-05-06
江苏农业科学(2020年22期)2020-03-03
中国种业(2020年2期)2020-02-26
农业与技术(2020年2期)2020-02-05
江苏农业科学(2019年5期)2019-09-02
山西农经(2019年5期)2019-02-22
现代农业科技(2018年14期)2018-10-31
广东农业科学(2017年5期)2017-08-29
应用海洋学学报(2014年1期)2014-11-22
