
大脑皮质神经元及其网络的兴奋
2017-03-14 01:47:31邓穗馨舒友生
中国药理学与毒理学杂志 2017年11期
邓穗馨,舒友生
(1.北京师范大学认知神经科学与学习国家重点实验室,2.IDG/麦戈文脑科学研究院,北京 100875)
大脑皮质中数量庞大的神经元作为神经系统基本单元,通过突触彼此连接,形成了能实现感觉、运动、学习、语言和决策等各种功能的复杂神经网络[1]。皮质的网络活动主要由释放谷氨酸的兴奋性神经元和释放γ-氨基丁酸(gamma aminobutyric acid,GABA)的抑制性神经元共同介导。这些神经元介导的兴奋性和抑制性信号相辅相成,共同维持大脑功能正常行使所必需的兴奋和抑制平衡。无论是在清醒状态下感觉引发的皮质网络活动中,亦或是在睡眠或麻醉状态下自发的网络振荡中,兴奋和抑制呈比例变化[2]。因此,研究兴奋性神经元和抑制性神经元产生电信号的基本原理、突触传递特性、基本环路结构以及兴奋-抑制平衡网络的调控机制,对于理解认知加工的过程十分重要。此外,皮质功能的紊乱、兴奋抑制的失衡常与神经性或精神性疾病有关,这些研究对了解脑疾病的发生机制也有重要意义[3-4]。本综述将围绕神经网络的兴奋-抑制平衡机制,分别从单细胞兴奋性、突触传递特性、抑制性微环路特异性及其动态变化特征等进行阐述。
1 皮质神经元的分类
大脑皮质的主体神经元是投射型谷氨酸能锥体细胞(pyramid cells,PC),占皮质神经元总数的70%~80%[5]。另一类神经元是非投射型谷氨酸能神经元多棘星型细胞,其树突较短呈星形放射状[6]。兴奋性神经元在皮质排列井然有序,分层聚集,根据其形态结构和电生理特征可分为最终投射终结于端脑内侧束神经元,具有离皮质投射能力投向脑干、脊髓和基底神经节等皮质下脑组织的锥体束神经元和特异性投向丘脑的皮质丘脑神经元[7-8]。皮质的抑制性神经元种类多样,一般仅在局部网络形成突触连接,因此常被称为中间神经元。可根据其形态结构、电生理学特征或特异性表达的分子等进行分类[5],如小清蛋白(parvalbumin,PV)和生长抑素(somatostatin,SST)阳性神经元占中间神经元总数的约70%[9]。特定类型中间神经元按照不同分类方式有较高的重合率。例如,PV神经元常为胞体和近端树突投射型中间神经元,其轴突一般呈篮状包裹PC胞体,电生理特征常呈快速放电模式[5,9];吊灯细胞是一类特殊的大多表达PV的神经元,其轴突主要支配PC的轴突始段[10]。又如,SST神经元多是远端树突投射型中间神经元,又称Martinotti细胞,且电生理特征常呈低阈值放电模式[5,9,11]。
灵长类大脑皮质中存在一些独特的神经元类型,如梭形神经元和持续放电神经元。梭形神经元是一类胞体较大且呈纺锤形的投射型兴奋性神经元,主要存在于前扣带回和前脑岛皮质的5b层[12],其异常被认为与孤独症[13]、精神分裂症[14]和额颞叶痴呆[15]相关。持续放电神经元在人脑皮质的非锥体细胞中所占比例约为9%,诱发单个动作电位(action potential,AP)的短暂刺激可引起持续去极化平台电位,并导致持续AP的发放;诱发高频AP发放的较强刺激则会抑制这种平台电位和AP的持续发放[16]。丰富的神经元类型及独特的突触连接方式可能对灵长类大脑皮质高级认知功能的实现具有重要贡献。
2 神经元的兴奋性
神经元通过产生AP对信息进行编码,参与大脑皮质的信息处理[17],同时产生AP的能力也反映了神经元的兴奋性。因此,对皮质网络的兴奋和抑制平衡机制的探讨,首先需要对网络中单个神经元的兴奋性决定机制进行研究。神经元去极化到特定的阈电位是AP爆发的关键,一旦去极化达到阈电位,神经元将不可逆地在轴突始段爆发AP,膜电位超射至约30 mV,此时更大的阈上电流刺激也不能使AP超射幅值发生太大变化;而阈下膜电位的波动将在轴突上产生电紧张性扩佈,最终衰减而无法传导至下一级神经元。因此,AP是“全或无”的信息编码模式,即1和0的数字信号,并通过时间和频率编码来主导大脑中神经信号的传导。近期研究显示,神经元阈下膜电位的波动通过影响AP介导突触传递[18-19]。具体表现为突触前阈下膜电位去极化程度越高,AP触发的突触后电位幅值越大。突触前这种膜电位波动不是“全或无”的,而是连续的,即模拟信号编码模式。因此,在大脑皮质中,神经元以数字信号与模拟信号相结合的方式共同编码神经信息,并决定下级神经元的兴奋。
2.1 谷氨酸能神经元兴奋性的决定机制及其调节
关于中枢神经系统中神经元兴奋性的早期研究,主要集中在各脑区的主体细胞。在锥体细胞上,一般认为轴突始段拥有的高密度的钠通道是决定AP首先在此爆发的原因[20]。更细致的工作发现,钠通道在PC的轴突始段胞体近端和远端虽均高密度分布,但AP首先在距离胞体约40 μm的远端爆发[21]。因此,除了高密度的钠通道,AP偏好于轴突始段远端爆发应该还存在其他机制。
AP发生机制的探讨着重于轴突膜上离子通道的生物物理性质的研究。由于大部分中枢神经元的轴突直径<1 μm,以往对轴突的记录仅局限于某些特殊的巨大轴突终末,如脑干花萼突触[22]和海马苔藓纤维突触节[23]。近期,一种新颖的轴突泡记录方法突破了以往的技术限制[19,24-26]。该方法是在脑薄片上用比正常更细的膜片钳电极直接记录轴突小泡,这一泡状结构是在制备脑薄片的过程中由轴突切断处的细胞膜重新愈合而形成的。通过对轴突小泡进行记录或对胞体和轴突小泡同时进行双电极膜片钳记录,可测量轴突上特定离子通道的密度、通道亚型电生理特性、AP波形及传导速度等。利用这种技术并结合改良的免疫荧光染色方法发现,在PC的轴突始段远端大量聚集的低阈值电位开放的Nav 1.6决定了AP首先在此爆发;而在轴突始段近端大量聚集的高阈值电位激活的Nav1.2则可进一步开放并促进AP回传至胞体,并决定胞体-树突区域AP爆发的阈值[27]。在人脑皮质的PC上,免疫荧光显示出相似的钠通道亚型分布模式[28],提示具有相似的AP产生机制。
神经调质或递质可调控轴突上的AP发生和传导。有文献报道,5-羟色胺1A受体(5-hydroxy tryptamine 1A receptor,5-HT1AR)在轴突始段的近端高密度分布[29]。激活5-HT1AR不影响轴突始段上Nav 1.6介导的钠电流,但会强烈抑制Nav1.2介导的钠电流,并阻碍AP向胞体的回传。这种对Nav1.2亚型的选择性调节,可能在神经元和神经环路信号处理以及相关脑区功能的调控方面具重要意义[30]。PC轴突上另一种重要的电压门控型离子通道——钾通道,可受到多巴胺受体D1和D2的调节,D1受体的激活可下调轴突上的钾电流,反之D2受体的激活则上调钾电流。因此,神经调质多巴胺可通过调节轴突钾电流来影响AP波形进而影响神经信号的传递[31]。另外,PC轴突上还存在GABAA受体,其开放主要使得轴突膜电位超极化,同时可能通过增加轴突局部的电导产生分流抑制效应,从而影响AP的幅度、宽度和AP引起的细胞内钙水平[32]。在人脑皮质组织中,PV阳性的吊灯细胞突触终末选择性地支配富含Nav 1.6的轴突始段远端,而在Nav1.2分布的轴突始近端上形成的突触较少,提示这类吊灯细胞释放的GABA作用于轴突受体,对PC的兴奋性发挥重要的调节作用[28]。
2.2 皮质抑制性神经元兴奋性的决定机制及其调节
抑制性中间神经元的电活动对皮质兴奋-抑制平衡以及信息处理具有十分关键的意义。抑制性环路功能的下调或中间神经元数目的减少常与各种脑疾病相关[33]。由于中枢神经系统中抑制性神经元的轴突比PC的更细,早期限于轴突记录技术难度,人们对AP在各类中间神经元上的起始和传播特征所知甚少。轴突小泡记录方法在抑制性神经元的研究中仍具巨大优势[24]。利用这一方法并结合免疫荧光染色技术研究发现,PV和SST神经元与PC相似,AP起始点也在富含Nav 1.6的轴突始段远端;但不同的是两类抑制性神经元的轴突始段均表达Nav1.1[34]。进一步实验发现,相较于SST神经元,PV神经元轴突钠通道的激活和失活曲线均向超极化方向移动了近7 mV,与两者的AP电压阈值差异一致。免疫荧光染色显示,这种差异可能源于SST神经元轴突始段上的表达更多的高阈值钠通道(Nav 1.1和Nav1.2)[34]。后续药理学实验表明,抑制Nav1.2电流可阻断SST神经元AP的自主发放,降低其对所介导的网络环路的抑制,从而提高癫痫样放电的频率[34]。
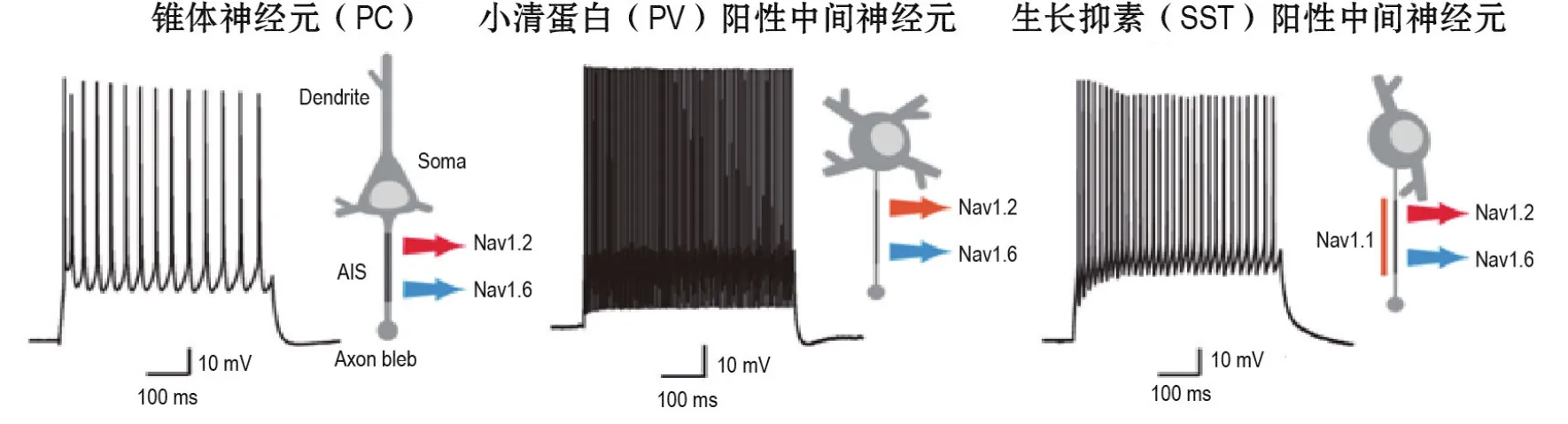
图1 大脑皮质各类神经元的电生理学特征及轴突始段钠通道表达模式.图片修改自Hu等[27]和Tian等[28].
以上研究结果阐明了皮质PC和2类抑制性中间神经元上轴突钠通道的组成(图1)以及各自兴奋性的决定机制,和各种神经调质或递质在控制AP爆发和传播中的重要作用。这些发现有助于了解离子通道疾病的病理基础,为控制细胞兴奋性的药物研发提供分子靶标。
3 突触传递模式
AP的爆发是细胞内在的生物物理特性与突触活动相互作用的结果。化学突触的突触间隙一般约20 nm,其间神经递质的浓度在突触活动中最大可达毫摩尔级别[35],从而实现2神经元之间特异性的突触传递。化学突触中,依赖突触前AP爆发的信号传递主要有2种模式,即同步化释放和非同步化释放[36]。
3.1 同步化递质释放
在多数情况下,AP爆发后沿轴突传导至轴突终末,激活电压门控钙离子通道并引发大量钙离子内流,促使囊泡内神经递质释放。此过程受到众多分子的复杂调控[37]。但从时间上来讲,突触前AP的爆发到突触后反应开始的时间间隔非常短暂(1~2 ms),在此极短时间窗内,大量囊泡释放,即递质的同步化释放[36]。突触结合蛋白(synaptotagmin,Syt)是突触前膜内最常见的钙感受器,敲除其亚型Syt-1基因将严重影响突触传递中的同步化递质释放[38]。
不同类型的突触由于其相关突触蛋白表达量或种类不同,其同步化释放有一定的短时可塑性。当突触前神经元受到一串刺激时,突触后反应大小会发生短时程的改变。第一个刺激引起的囊泡释放率被用来表征不同类型突触对AP信号的敏感性。囊泡释放率往往与双脉冲比率呈负相关。在锥体细胞分别与SST和PV神经元形成的突触上,囊泡释放率差异较大。表现为PC-SST突触拥有较低的囊泡释放率,而PC-PV突触拥有较高的囊泡释放率[39]。因此,当给予PC一串刺激后,在SST神经元上记录到的是随刺激数目增加而逐渐增大的短时程易化突触反应,而在PV神经元上得到逐渐减小的短时程抑制突触反应[40-41]。
PC不同突触的短时程可塑性差异可能与其突触相关蛋白表达的特异性有关。但即使突触前神经元是同一个锥体细胞,在PC对SST和PV神经元分别形成的突触中也能检测到不同的短时程可塑性[42],提示其突触相关蛋白表达的特异性只局部体现在其不同轴突终末上。因此,这种局部特异性的突触相关蛋白的表达,可能依赖于突触后神经元特异性的逆向跨突触分子信号。Elfn1是其中一类可能具逆向跨突触调控作用的蛋白,这种蛋白被发现特异性表达在SST神经元上。当敲除SST神经元中此蛋白的编码基因,可发现PC-SST突触的短时程易化特征受到抑制;而在无这种蛋白表达的PV神经元上外源过表达此蛋白,可导致原先突触前PC-PV神经元的短时程抑制转化成易化模式[43]。此外,SST神经元还特异性地表达代谢型谷氨酸受体1(metabotropic glutamate receptor 1,mGluR1),与其相对应的突触前神经元则特异性表达mGluR7[44]。研究发现,含有mGluR7蛋白的突触前神经元与含有Elfn1蛋白的突触后在空间上的耦合度非常高,敲除Elfn1将导致其突触前神经元mGluR7的聚集程度下降,并且敲除Elfn1的小鼠与敲除mGluR7的小鼠拥有较为相似的疾病表型,提示SST神经元表达的Elfn1可能提供了某种逆向跨突触信号,来调控其突触前mGluR7蛋白的表达与分布[45]。此外,近期研究结果表明,钙感受器Syt-7在调节囊泡释放模式方面有较强作用,在具有短时程易化可塑性的突触中特异性地敲除Syt-7,可检测到短时程易化的消失[46]。
3.2 非同步化递质释放
如上所述,一般AP诱导的突触前囊泡递质释放与AP爆发时间耦合较好,并能在2 ms时间窗内记录到相应的突触后电位。在某些特定的突触中,当AP结束后较长的时间窗(几十至几百毫秒)内仍能检测到连续不断的递质释放事件,这种时间上比AP发生延迟许多的递质释放称为非同步化释放[47]。研究认为,AP强烈发放后,轴突终末累积的残余钙信号触发了囊泡的非同步化释放,因此非同步化释放水平一般与刺激强度(AP频率或数目)呈正相关[36,47]。
中枢神经系统内的非同步化递质释放模式,主要存在于抑制性神经元快速放电的GABA释放中[48-50]。这种GABA的非同步化释放,随着动物发育至成年有逐渐减少的趋势,提示环路对这种非同步化GABA释放产生的抑制的需求随发育而发生变化[50]。关于非同步化GABA释放机制的探讨,则在海马中更为具体。海马齿状回中,非同步化GABA释放也在一类抑制性神经元的输出突触中存在,但不是PV神经元,而是另一种分子标签为胆囊收缩素的中间神经元[47]。胆囊收缩素神经元和PV神经元的轴突虽都偏好于呈篮状包裹PC胞体[51],但它们的轴突终末蛋白在表达和功能上有很大区别,可能引发其非同步化释放水平上的差异。首先,其突触终末介导钙离子内流电压门控钙离子通道的亚型不同,胆囊收缩素神经元表达N型,而PV神经元则表达P/Q型。不同亚型电压门控钙离子通道与钙感受器之间的距离可能导致囊泡释放速率的差异。实验中,用慢钙螯合剂乙二醇双(2-氨基乙基醚)四乙酸(EGTA)可阻断胆囊收缩素神经元的GABA释放,提示电压门控钙离子通道与钙感受器的距离较远,属于微米级偶联(距离>100 nm),即钙离子需要扩散较远的距离才能抵达钙感受器,并导致囊泡释放在时间上的延迟;而PV神经元GABA的释放需要用快钙螯合剂BAPTA才能阻断,提示轴突终末上两者的距离较近,为纳米级偶联(<100 nm),因此囊泡释放时间较为精准[47]。其次,胆囊收缩素神经元和PV神经元的突触后膜表达的受体类型不一致。胆囊收缩素神经元的突触末端表达大麻素1型受体,而PV神经元的突触末端表达乙酰胆碱和脑啡肽受体,这使得PV神经元在突触活动中易受到其配体的特异性调控[51]。第三,分子标志物PV是一种强有效的钙结合蛋白,可与钙离子结合并影响轴突终末的游离钙浓度,从而调节突触活动,如减弱突触囊泡的非同步化释放水平或降低同步化释放的短时程抑制程度[48]。
此外,上文提到的支撑短时程易化的钙感受器Syt-7,在早期斑马鱼神经肌肉接头的研究中被认为是特异性的非同步化释放的钙感受器[52]。后续在啮齿类动物神经元上的实验发现,只有在敲除了介导同步化释放的钙感受器Syt-1的前提下,Syt-7才能体现出其在非同步化释放过程中的重要作用,而仅仅单敲Syt-7对突触传递影响不大[53-54]。进一步研究发现,单敲Syt-7虽无法检测到明显的突触传递变化,但同步化释放过程中的一种较慢的突触后基电流大大降低,最终可导致突触后神经元上AP发放的减少与发放时间精准性的下调[55]。可见,Syt-7虽不如Syt-1对突触传递的贡献大,但其支撑的非同步化释放对突触后神经元的兴奋性仍有较强调控作用,从而可能影响整个网络的电活动。
4 皮质交互式微环路的特征
神经元的突触输入主要来自外界刺激和交互式网络活动。在感受外界刺激时,大脑皮质主要接收并加工来自丘脑的信息输入。视皮质的研究发现,丘脑的输入可在视觉刺激40 ms后激活皮质内部的交互式微环路,而此微环路的兴奋性强度远超于从丘脑直接输入的刺激兴奋;但若切断丘脑输入,此交互式兴奋将迅速消失,说明丘脑输入的信号是维持视觉刺激诱发的皮质兴奋的必要条件,而此丘脑信号在皮质内可被局部交互式微环路的兴奋放大[56]。大脑皮质中各种抑制性神经元也参与到功能相关的交互式网络构建中,并与兴奋性神经元通过突触连接形成特定的交互抑制微环路,从而调控网络活动的输入和输出。交互抑制微环路除了在动物清醒时通过丘脑的信号输入而被激活,在麻醉或睡眠状态下也可被自发网络振荡活动所驱动。
不同抑制性神经元在接受谷氨酸递质同步化释放所展现出的短时程可塑性差异,决定了它们所介导的抑制性信号在交互微环路中功能的特异性[40]。在PC-快速放电神经元-PC组成的皮质交互抑制微环路中,快速放电神经元一般仅在其突触前PC刺激串输入的前期发放AP,并在其突触后PC上产生一个快相的抑制;而在PC-低阈值放电神经元-PC组成的微环路中,低阈值放电神经元一般在刺激串输入的后期才发放AP,随后在其突触后PC上产生一个慢相的抑制[40]。此外,皮质中血管活性肠肽阳性中间神经元,易受到其他高级脑区自上而下信息的调节,或皮质下结构投射来的神经调质纤维的作用,如乙酰胆碱[57]。这类细胞的激活主要通过抑制SST神经元从而在邻近的PC上引起去抑制效应,对网络活动的产生具有重要的调控作用[57-58]。
4.1 快相交互抑制微环路
双电极膜片钳记录发现,快速放电神经元易与邻近PC形成双向突触;而当同时记录2个PC并对其中一个进行串刺激时,却很少能在另一个PC上检测到快相的跨双突触抑制反应(<1%)[59-60]。此现象提示2种可能:①由快速放电神经元所介导的快相交互抑制微环路主要由有双向突触联系的PC和快速放电神经元组成,只负责反馈抑制;②快速放电神经元也介导侧抑制(surround suppression),但在双电极膜片钳记录条件限制下,仅单个突触前PC的活动并不足以激活快速放电中间神经元介导的快相交互抑制微环路。实验发现,快速放电神经元常被报道参与同步化的网络活动中[61],即网络中大量PC同步化爆发的AP才能更有效地激活快相交互抑制微环路。因此,快速放电神经元又常被认为是环路中的“同步探测器”[62]。在环路功能上,这样的抑制信号能维持网络活动中的兴奋-抑制平衡,保护网络不致过于兴奋而引发癫痫或其他脑疾病[63-64]。此外,快相交互抑制微环路还大量被招募到前馈式抑制活动中[65]。在皮质的第4层,当兴奋性神经元接受大量来自丘脑的输入时,这份兴奋也同样被抄送给了快速放电神经元,由快速放电神经元所介导的前馈抑制使得兴奋性神经元接受丘脑的输入后立刻受到抑制。这样的前馈抑制,即快速放电所介导的快相交互抑制能缩小目标兴奋性神经元的兴奋性整合的时间窗,压缩目标兴奋性神经元群发放AP的时间窗,并提高发放时间的精确性,促进兴奋性神经元群的同步化活动。同样的前馈抑制还发生在第4层兴奋性神经元向第2/3层投射的过程中,并且第4层的兴奋性神经元偏好性地投射给2/3层中互相投射的PC和PV神经元,从而进一步加强交互式环路中的反馈抑制[66]。2/3层的PV神经元是保持网络中兴奋-抑制平衡的重要组成,通过动态调整PC的兴奋性,从而改变其突触前PV神经元的抑制性输入大小[67]。可见快速放电神经元介导的快相交互抑制微环路,不论是产生反馈式抑制还是前馈式抑制,均通过随网络兴奋增高而抑制增强的模式,贡献于正常生理活动中的兴奋-抑制平衡,保护神经系统不至于过兴奋。此外,快速放电神经元特异性投射到PC的胞体或近端树突上,因而能更加快速而有效地保证其行使抑制功能[51]。
4.2 慢相交互抑制微环路
较之快相交互抑制微环路,当给予同样的串刺激条件,PC-低阈值放电神经元-锥体细胞这种慢相交互抑制微环路所介导的跨突触抑制反应在双PC膜片钳记录中较易得到;PC-低阈值放电神经元的单突触联系中,低阈值放电神经元上能得到AP的概率高达27%[59-60]。这说明低阈值放电中间神经元较易被单个PC所激活,从而产生AP并形成对下一级PC的抑制。但由于PC对低阈值放电神经元突触输入的短时程易化效应,单PC的低强度输入并不能有效激活慢相交互抑制微环路。实验发现,低阈值放电神经元介导的跨突触抑制效果随突触前刺激频率和数目增加而增强[59],因此低阈值放电神经元又可被称作“簇状发放探测器”。另外,3个PC膜片钳记录发现,同时激活2个PC在第3个PC上得到的跨突触抑制反应比分别单独激活得到的反应的数学加和要大得多。这种跨突触抑制反应的超线性增强的现象表明,少量PC的簇状发放即可强烈激活由低阈值放电介导的慢相交互抑制微环路。因此,在某些特定网络中,只需激活极少量的PC就能达到对整个网络活动的调控[68]。在多个SST神经元上分别用解笼锁谷氨酸(uncage glutamate)进行刺激,在同一个突触后PC上可高概率检测到抑制性反应,并且在邻近的其他PC上有相似的结果[69]。可见这种SST神经元抑制性的投射缺乏选择性。综上,1个PC可在大范围内接受多个突触前SST神经元的密集投射,同时,突触前的SST神经元也可能同时投射给其他PC;1个SST神经元能接受多个突触前PC兴奋性的输入,同时又给多个下游PC提供抑制性。此外,PC与SST神经元还存在相互投射,从而形成一庞大而复杂的慢相交互抑制大网络,来提供大范围内相对长时程的皮质交互性抑制,特别是侧抑制[66],从而调控整个皮质的兴奋-抑制平衡。
实际上,在高等动物或正常清醒状态的低等动物中,这种交互式抑制在感觉信息处理过程中的作用更为强大。近年来,人们提出一种新型神经网络动态兴奋的理论模型抑制稳定网络(inhibition stabilized network,ISN)[70-71]来解析抑制性在其中的重要作用。由于这个ISN模型需要非常强大的交互式网络存在,所以最先在猴、猫等有明显功能柱结构的视皮质被证明,近期才在清醒小鼠的听、视皮质被相应证实[72-73]。这些研究表明,ISN的主要贡献者是SST神经元,它们可通过其不论水平还是垂直方向均广泛的侧抑制[66,72-73]来主导这个网络的稳态。这种大范围抑制的产生可能得益于SST神经元聚集于第一层丰富的轴突分支。SST神经元通过其分支广泛的轴突、无选择性地抑制大量PC的顶树突,减弱PC树突钙电位的发生,从而抑制PC的AP发放[74-75]。此外,这种对顶树突的抑制还主要体现在减弱PC的簇状发放上[76-77],对感觉信息的编码与传递、知觉的形成等有重要调节作用。
5 抑制性微环路的功能与动态变化
皮质神经元组成了复杂的动态变化的交互型神经网络,并通过发放AP来参与不同的生理功能或病理活动[78]。在大部分网络活动中,交互式兴奋和抑制往往是相辅相成的,两者之间维持着适当的兴奋-抑制平衡[79-81]。抑制性微环路在平衡的维持中发挥重要作用,不同类型抑制性中间神经元在时间和空间上,可能贡献于不同的动态网络信息编码过程,从而精细调控不同的生理或病理过程[82]。
5.1 网络电活动与膜电位依赖的交互抑制调控
大脑皮质行使正常生理功能时,皮质神经元的膜电位由于受到突触前各种兴奋性、抑制性输入的影响,除了爆发AP,还常常产生呈一定规律波动的膜电位振荡[79,83]。在麻醉或睡眠动物的胞外或胞内电生理记录过程中,均能记录到神经元交互呈现的两种不同但相对稳定的膜电位状态。一种是平均10~20 mV的去极化膜电位,常伴有AP发生,称为高电位状态(up state);一种是膜电位接近于静息膜电位,几乎无突触输入和AP产生,称为低电位状态(down state)[79,84]。此外,若在改良的人工脑脊液(钙、镁离子浓度降至1~1.2 mmol·L-1或给予乙酰胆碱受体激动剂)中对脑片进行记录,也常得到这种高电位状态和低电位状态[79,83,85]。在神经元处于高电位状态时,可明显检测到其兴奋性的增加,以及对微小输入敏感性的增强,从而引发AP发放增多并缩短AP爆发的延迟时间,提示这种高电位状态的膜电位对信息的编码有重要作用[79,86]。这种交互式网络振荡机制和调节方式迄今尚不清楚,有研究表示抑制性神经元在其中发挥重要作用[83]。
越来越多的工作表明,突触前细胞膜电位依赖的突触传递是AP编码信息的另一种模式,即模拟信号编码模式[19,60]。此模式主要表现为,在突触前神经元处于去极化状态时给予刺激使其爆发AP,其递质释放量将增多,从而导致突触后神经元上突触反应的增大。因此,这种模式可导致突触后神经元更易达到AP的阈值,根据突触后神经元类型及特性来调控交互式神经环路中信号的传递。研究表明,不论是快相交互式抑制还是慢相交互式抑制,在突触前膜电位去极化状态下都被增强[60](图2)。这种膜电位依赖的突触调控主要依靠突触前PC轴突上电压门控钾离子通道Kv1的参与[26,60]。此外,研究发现利用神经元胞体和轴突泡同时记录的方法,当神经元膜电位处于去极化15~20 mV的状态时,从轴突始段传导至远端轴突的AP波形比在静息膜电位状态下更宽,但在胞体处记录到的波形变化不明显[19]。除了AP沿着轴突传导外,胞体的膜电位波动也会通过电紧张扩佈方式传导至轴突远端。在前额叶皮质第5层PC轴突上,扩佈的长度常数>400 μm[19]。因此,对于一个特定的突触来说,胞体去极化可能通过传播到达突触前膜并导致Kv1通道的失活、AP宽度和钙内流的增加,引起递质释放量的增多和突触后反应的增强,最终导致突触后神经元AP发放概率的上升和数目的增多。这些研究均指示,模拟信号对皮质微环路的功能具有重要的调控功能。
5.2 刺激强度依赖的非同步化GABA释放与癫痫
如上所述,皮质快速放电神经元的输出突触上存在GABA的同步化释放和非同步化释放。后者的强度依赖快速放电神经元上AP产生的频率和数目,当刺激强度增强产生高频簇状AP时,非同步化释放增多。因此,皮质网络可能利用此机制动态调控网络的兴奋-抑制的平衡。研究发现,在癫痫患者的脑片中,快速放电神经元的非同步化递质释放较非癫痫患者的更为强烈(图3)。这种增强的非同步化GABA释放,不论是在快速放电神经元的自突触还是在其投向PC的抑制性突触中均存在,但其GABA的同步化释放水平却无太大差异[49]。因此,快速放电神经元的GABA非同步化释放将在突触后神经元上产生一个长时间的抑制,在不同突触中,这种抑制会影响不同的突触后神经元AP的发放率与发放时间精确性[48],从而改变神经网络中的兴奋-抑制平衡,可能使得神经网络活动去同步化来调节癫痫的爆发和维持[49]。综上,快速放电神经元的非同步化GABA释放的改变可能调控癫痫的发作和维持,这些研究可为抗癫痫药物的研发提供新的思路。
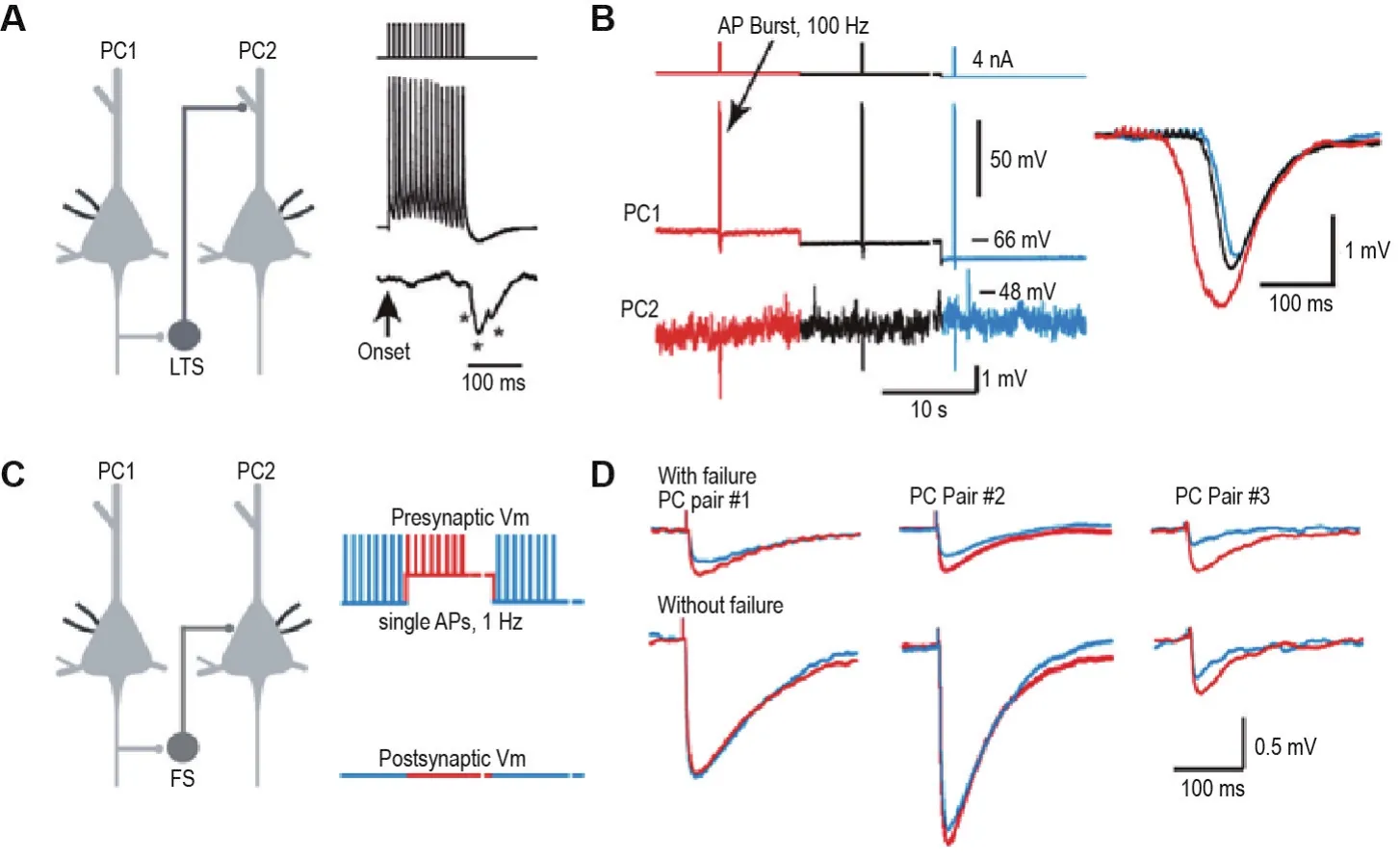
图2 膜电位依赖的交互抑制调控.A:利用双电极膜片钳技术同时记录2个PC,当给予PC1一串刺激(100 Hz,15个AP),可在PC2上检测到由低阈值放电神经元介导的慢相跨双突触抑制性电位;B:PC1膜电位的去极化水平与PC2记录得到的跨双突触抑制性电位大小紧密相关;C:双电极膜片钳技术同时记录2个PC,将PC1钳制在不同膜电位水平并给予持续的1 Hz刺激产生AP;D:3例展示检测到的快相跨双突触抑制性电位在PC1膜电位去极化状态下更大(红色指示PC1膜电位处于去极化状态下的记录;蓝色指示静息膜电位状态下的记录).AP:动作电位.图片修改自Zhu等[60].
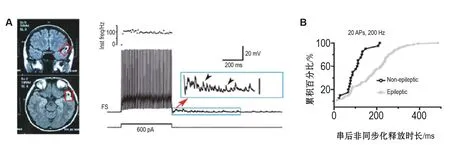
图3 皮质快速放电中间神经元的非同步化GABA释放与癫痫.A:来自神经胶质瘤患者的脑部核磁共振成像图片,此患者伴有严重的癫痫症状,红框指示其位于大脑颞叶的胶质瘤病灶区.利用膜片钳技术得到的胶质瘤病灶旁组织的脑切片中的神经元电生理记录个例.放电频率分析指示其为典型的快速放电中间神经元,并且在刺激结束后还能检测GABA非同步化释放所引起的突触后反应(蓝框).B:癫痫患者和非癫痫患者脑切片中快速放电神经元在刺激串后GABA非同步化释放持续时间的比较.图片修改自Jiang等[49].
6 总结
大脑皮质网络由丰富多样的神经元类型通过突触连接形成功能微环路,各种细胞类型分别表达特异性功能蛋白(如离子通道、突触相关蛋白等),它们的组合决定了各类神经元的兴奋性及其介导的微环路结构和功能,从而贡献于复杂多变的神经网络活动。综述主要从单细胞水平、突触水平和微环路水平阐述了神经元及其网络兴奋性的决定机制,及交互抑制性微环路如何动态调控网络中兴奋-抑制平衡,其中重点介绍了快速放电和低阈值放电的2类抑制性中间神经元所介导的微环路及其功能。随着基因组学、电生理学、病毒学和光遗传学等技术的日益成熟和应用[87],更多关于脑网络兴奋-抑制平衡机制的科学问题将得到深入研究。
除了梭形神经元和持续放电神经元,皮质中是否存在未被发现的新型神经元种类,不同的神经递质或调质如何特异性地调控这些神经元亚类,在整体动物生理情况下,PV和SST 2类中间神经元是如何被有效激活的,是否存在特殊的分子靶点来有效激活或调控这2类中间神经元所介导的交互抑制,从而影响网络活动功能,数字信号和模拟信号结合的信号传递模式在网络活动中又如何被精准的调控,这些问题仍亟待研究。针对这些问题的研究将为脑网络中兴奋-抑制平衡的调控提供新策略,为相关脑疾病的治疗提供新思路。
[1 ]Schüz A.Cortex:statistics and geometry of neuronal connectivity[J].J Anat,1999,194(pt1):153-157.
[2]Okun M,Lampl I.Instantaneous correlation of excitation and inhibition during ongoing and sensoryevoked activities[J].Nat Neurosci,2008,11(5):535-537.
[3 ]Cobos I,Calcagnotto ME,Vilaythong AJ,Thwin MT,Noebels JL,Baraban SC,et al.Mice lacking Dlx1 show subtype-specific loss of interneurons,reduced inhibition and epilepsy[J].Nat Neurosci,2005,8(8):1059-1068.
[4]Lewis DA,Hashimoto T,Volk DW.Cortical inhibitory neurons and schizophrenia[J].Nat Rev Neurosci,2005,6(4):312-324.
[5]Markram H,Toledo-Rodriguez M,Wang Y,Gupta A,Silberberg G,Wu C.Interneurons of the neocortical inhibitory system[J].Nat Rev Neurosci,2004,5(10):793-807.
[6 ]Petersen CC,Crochet S.Synaptic computation and sensory processing in neocortical layer 2/3[J].Neuron,2013,78(1):28-48.
[7]Harris KD,Shepherd GM.The neocortical circuit:themes and variations[J].Nat Neurosci,2015,18(2):170-181.
[8 ]Naka A,Adesnik H.Inhibitory circuits in cortical layer 5[J].Front Neural Circuits,2016,10:35.
[9]Tremblay R,Lee S,Rudy B.GABAergic interneurons in the neocortex:from cellular properties to circuits[J].Neuron,2016,91(2):260-292.
[10]Taniguchi H,Lu J,Huang ZJ.The spatial and temporal origin of chandelier cells in mouse neocortex[J].Science,2013,339(6115):70-74.
[11 ]McGarry LM,Packer AM,Fino E,Nikolenko V,Sippy T,Yuste R.Quantitative classification of somatostatin-positive neocortical interneurons identifies three interneuron subtypes[J].Front Neural Circuits,2010,4:12.
[12]Nimchinsky EA,Gilissen E,Allman JM,Perl DP,Erwin JM,Hof PR.A neuronal morphologic type unique to humans and great apes[J].Proc Natl Acad Sci USA,1999,96(9):5268-5273.
[13]Allman JM,Watson KK,Tetreault NA,Hakeem AY.Intuition and autism:a possible role for Von Economo neurons[J].Trends Cogn Sci,2005,9(8):367-373.
[14 ]Krause M,Theiss C,Brüne M.Ultrastructural alterations of Von Economo neurons in the anterior cingulate cortexin schizophrenia [J].AnatRec(Hoboken),2017,300(11):2017-2024.
[15]Seeley WW,Carlin DA,Allman JM,Macedo MN,Bush C,Miller BL,et al.Early frontotemporal dementia targets neurons unique to apes and humans[J].Ann Neurol,2006,60(6):660-667.
[16]Wang B,Yin L,Zou X,Ye M,Liu Y,He T,et al.A subtype of inhibitory interneuron with intrinsic persistent activity in human and monkey neocortex[J].Cell Rep,2015,10(9):1450-1458.
[17 ]Bean BP.The action potential in mammalian central neurons[J].Nat Rev Neurosci,2007,8(6):451-465.
[18 ]Alle H,Geiger JR.Combined analog and action potential coding in hippocampal mossy fibers[J].Science,2006,311(5765):1290-1293.
[19]Shu Y,Hasenstaub A,Duque A,Yu Y,McCormick DA.Modulation of intracortical synaptic potentials by presynaptic somatic membrane potential[J].Nature,2006,441(7094):761-765.
[20]Kole MH,Ilschner SU,Kampa BM,Williams SR,Ruben PC,Stuart GJ.Action potential generation requires a high sodium channel density in the axon initial segment[J].Nat Neurosci,2008,11(2):178-186.
[21]Palmer LM,Stuart GJ.Site of action potential initiation in layer 5 pyramidal neurons[J].J Neurosci,2006,26(6):1854-1863.
[22 ]Schneggenburger R,Meyer AC,Neher E.Released fraction and total size of a pool of immediately available transmitter quanta at a calyx synapse[J].Neuron,1999,23(2):399-409.
[23 ]Geiger JR,Jonas P.Dynamic control of presynaptic Ca2+inflow by fast-inactivating K+channels in hippocampal mossy fiber boutons[J].Neuron,2000,28(3):927-939.
[24 ]Hu W,Shu Y.Axonal bleb recording[J].Neurosci Bull,2012,28(4):342-350.
[25 ]Shu Y,Duque A,Yu Y,Haider B,McCormick DA.Properties of action-potential initiation in neocortical pyramidal cells:evidence from whole cell axon recordings[J].J Neurophysiol,2007,97(1):746-760.
[26 ]Shu Y,Yu Y,Yang J,McCormick DA.Selective control of cortical axonal spikes by a slowly inactivating K+current[J].Proc Natl Acad Sci USA,2007,104(27):11453-11458.
[27]Hu W,Tian C,Li T,Yang M,Hou H,Shu Y.Distinct contributions of Nav1.6 and Nav1.2 in action potential initiation and backpropagation[J].Nat Neurosci,2009,12(8):996-1002.
[28 ]Tian C,Wang K,Ke W,Guo H,Shu Y.Molecular identity of axonal sodium channels in human cortical pyramidal cells[J].Front Cell Neurosci,2014,8:297.
[29 ]Cruz DA,Eggan SM,Azmitia EC,Lewis DA.Serotonin1A receptors at the axon initial segment of prefrontal pyramidal neurons in schizophrenia[J].Am J Psychiatry,2004,161(4):739-742.
[30 ]Yin L,Rasch MJ,He Q,Wu S,Dou F,Shu Y.Selective modulation of axonal sodium channel subtypes by 5-HT1A receptor in cortical pyramidal neuron[J].Cereb Cortex,2017,27(1):509-521.
[31]Yang J,Ye M,Tian C,Yang M,Wang Y,Shu Y.Dopaminergicmodulationofaxonalpotassium channels and action potential waveform in pyramidal neurons of prefrontal cortex[J].J Physiol,2013,591(13):3233-3251.
[32]Xia Y,Zhao Y,Yang M,Zeng S,Shu Y.Regulation of action potential waveforms by axonal GABAA receptors in cortical pyramidal neurons[J].PLoS One,2014,9(6):e100968.
[33]Knopp A,Frahm C,Fidzinski P,Witte OW,Behr J.Loss of GABAergic neurons in the subiculum and its functional implications in temporal lobe epilepsy[J].Brain,2008,131(Pt 6):1516-1527.
[34]Li T,Tian C,Scalmani P,Frassoni C,Mantegazza M,Wang Y,et al.Action potential initiation in neocortical inhibitory interneurons[J].PLoS Biol,2014,12(9):e1001944.
[35]Mozrzymas JW.Dynamism of GABAAreceptor activation shapes the"personality"of inhibitory synapses[J].Neuropharmacology,2004,47(7):945-960.
[36 ]Kaeser PS,Regehr WG.Molecular mechanisms for synchronous,asynchronous,and spontaneous neurotransmitter release[J].Annu Rev Physiol,2014,76:333-363.
[37]Südhof TC.Neurotransmitter release:the last millisecond in the life of a synaptic vesicle[J].Neuron,2013,80(3):675-690.
[38 ]Südhof TC.Calcium control of neurotransmitter release [J].ColdSpringHarbPerspectBiol,2012,4(1):a011353.
[39 ]Koester HJ,Johnston D.Target cell-dependent normalization of transmitter release at neocortical synapses[J].Science,2005,308(5723):863-866.
[40 ]Silberberg G.Polysynaptic subcircuits in the neocortex:spatial and temporal diversity[J].Curr Opin Neurobiol,2008,18(3):332-337.
[41 ]Jackman SL,Regehr WG.The mechanisms and functions ofsynaptic facilitation [J].Neuron,2017,94(3):447-464.
[42]Rozov A,Burnashev N,Sakmann B,Neher E.Transmitter release modulation by intracellular Ca2+buffers in facilitating and depressing nerve terminals of pyramidal cells in layer 2/3 of the rat neocortex indicates a target cell-specific difference in presynaptic calcium dynamics[J].J Physiol,2001,531(Pt 3):807-826.
[43 ]Sylwestrak EL,Ghosh A.Elfn1 regulates targetspecific release probability at CA1-interneuron synapses[J].Science,2012,338(6106):536-540.
[44 ]Somogyi P,Dalezios Y,Luján R,Roberts JD,Watanabe M,Shigemoto R.High level of mGluR7 in the presynaptic active zones of select populations of GABAergic terminals innervating interneurons in the rat hippocampus[J].Eur J Neurosci,2003,17(12):2503-2520.
[45]Tomioka NH,Yasuda H,Miyamoto H,Hatayama M,Morimura N,Matsumoto Y,et al.Elfn1 recruits presynaptic mGluR7 in trans and its loss results in seizures[J].Nat Commun,2014,5:4501.
[46 ]Jackman SL,Turecek J,Belinsky JE,Regehr WG.The calcium sensor synaptotagmin 7 is required for synaptic facilitation[J].Nature,2016,529(7584):88-91.
[47]Hefft S,Jonas P.Asynchronous GABA release generates long-lasting inhibition at a hippocampal interneuron-principal neuron synapse[J].Nat Neurosci,2005,8(10):1319-1328.
[48 ]Manseau F,Marinelli S,Méndez P,Schwaller B,Prince DA,Huguenard JR,et al.Desynchronization of neocortical networks by asynchronous release of GABA at autaptic and synaptic contacts from fast-spiking interneurons[J].PLoS Biol,2010,8(9):e1000492.
[49 ]Jiang M,Zhu J,Liu YP,Yang MP,Tian CP,Jiang S,et al.Enhancement of asynchronous release from fast-spiking interneuron in human and rat epileptic neocortex[J].PLoS Biol,2012,10(5):e1001324.
[50]Jiang M,Yang M,Yin L,Zhang X,Shu Y.Developmental reduction of asynchronous GABA release from neocortical fast-spiking neurons[J].Cereb Cortex,2015,25(1):258-270.
[51 ]Freund TF,Katona I.Perisomatic inhibition[J].Neuron,2007,56(1):33-42.
[52]Wen H,Linhoff MW,McGinley MJ,Li GL,Corson GM,Mandel G,et al.Distinct roles for two synaptotagmin isoforms in synchronous and asynchronous transmitter release at zebrafish neuromuscular junction[J].Proc Natl Acad Sci USA,2010,107(31):13906-13911.
[53 ]Maximov A,Lao Y,Li H,Chen X,Rizo J,Sørensen JB,et al.Genetic analysis of synaptotagmin-7 function in synaptic vesicle exocytosis[J].Proc Natl Acad Sci USA,2008,105(10):3986-3991.
[54 ]Bacaj T,Wu D,Yang X,Morishita W,Zhou P,Xu W,et al.Synaptotagmin-1 and synaptotagmin-7 trigger synchronous and asynchronous phases of neurotransmitter release[J].Neuron,2013,80(4):947-959.
[55 ]Luo F,Südhof TC.Synaptotagmin-7-mediated asynchronous release boosts high-fidelity synchronous transmission at a central synapse[J].Neuron,2017,94(4):826-839.e3.
[56 ]Reinhold K,Lien AD,Scanziani M.Distinct recurrent versus afferent dynamics in cortical visual processing[J].Nat Neurosci,2015,18(12):1789-1797.
[57 ]Zhang S,Xu M,Kamigaki T,Hoang Do JP,Chang WC,Jenvay S,et al.Selective attention.Long-range and local circuits for top-down modulation of visual cortex processing[J].Science,2014,345(6197):660-665.
[58]Pfeffer CK,Xue M,He M,Huang ZJ,Scanziani M.Inhibition of inhibition in visual cortex:the logic of connections between molecularly distinct interneurons[J].Nat Neurosci,2013,16(8):1068-1076.
[59 ]Silberberg G,Markram H.Disynaptic inhibition between neocortical pyramidal cells mediated by Martinotti cells[J].Neuron,2007,53(5):735-746.
[60]Zhu J,Jiang M,Yang M,Hou H,Shu Y.Membrane potential-dependent modulation of recurrent inhibition in rat neocortex[J].PLoS Biol,2011,9(3):e1001032.
[61 ]Bartos M,Vida I,Jonas P.Synaptic mechanisms of synchronized gamma oscillations in inhibitory interneuron networks [J].NatRev Neurosci,2007,8(1):45-56.
[62 ]Pouille F,Scanziani M.Routing of spike series by dynamic circuits in the hippocampus[J].Nature,2004,429(6993):717-723.
[63]Nelson SB,Valakh V.Excitatory/inhibitory balance and circuit homeostasis in autism spectrum disorders[J].Neuron,2015,87(4):684-698.
[64]Dehghani N,Peyrache A,Telenczuk B,Le Van Quyen M,Halgren E,Cash SS,et al.Dynamic balance of excitation and inhibition in human and monkey neocortex[J].Sci Rep,2016,6:23176.
[65 ]Hu H,Gan J,Jonas P.Interneurons.Fast-spiking,parvalbumin+GABAergic interneurons:from cellular design to microcircuit function[J].Science,2014,345(6196):1255263.
[66 ]Adesnik H,Bruns W,Taniguchi H,Huang ZJ,Scanziani M.A neural circuit for spatial summation in visual cortex[J].Nature,2012,490(7419):226-231.
[67]Xue M,Atallah BV,Scanziani M.Equalizing excitation-inhibition ratios across visual cortical neurons[J].Nature,2014,511(7511):596-600.
[68 ]Kapfer C,Glickfeld LL,Atallah BV,Scanziani M.Supralinear increase of recurrent inhibition during sparse activity in the somatosensory cortex[J].Nat Neurosci,2007,10(6):743-753.
[69 ]Fino E,Yuste R.Dense inhibitory connectivity in neocortex[J].Neuron,2011,69(6):1188-1203.
[70 ]Rubin DB, Van Hooser SD, Miller KD.The stabilized supralinear network:a unifying circuit motif underlying multi-input integration in sensory cortex[J].Neuron,2015,85(2):402-417.
[71 ]Litwin-Kumar A,Rosenbaum R,Doiron B.Inhibitory stabilization and visual coding in cortical circuits with multiple interneuron subtypes[J].J Neurophysiol,2016,115(3):1399-1409.
[72 ]Adesnik H.Synaptic mechanisms of feature coding in the visual cortex of awake mice[J].Neuron,2017,95(5):1147-1159.e4.
[73 ]Kato HK,Asinof SK,Isaacson JS.Network-level control of frequency tuning in auditory cortex[J].Neuron,2017,95(2):412-423.e4.
[74 ]Urban-Ciecko J,Barth AL.Somatostatin-expressing neurons in cortical networks[J].Nat Rev Neurosci,2016,17(7):401-409.
[75 ]Neske GT,Connors BW.Distinct roles of SOM and VIP interneurons during cortical up states[J].Front Neural Circuits,2016,10:52.
[76 ]Gentet LJ,Kremer Y,Taniguchi H,Huang ZJ,Staiger JF,Petersen CC.Unique functional properties of somatostatin-expressing GABAergic neurons in mouse barrel cortex[J].Nat Neurosci,2012,15(4):607-612.
[77 ]Royer S,Zemelman BV,Losonczy A,Kim J,Chance F,Magee JC,et al.Control of timing,rate and bursts of hippocampal place cells by dendritic and somatic inhibition[J].Nat Neurosci,2012,15(5):769-775.
[78 ]Silberberg G,Grillner S,LeBeau FE,Maex R,Markram H.Synaptic pathways in neural microcircuits[J].Trends Neurosci,2005,28(10):541-551.
[79 ]Shu Y, Hasenstaub A, Badoual M, Bal T,McCormick DA.Barrages of synaptic activity control the gain and sensitivity of cortical neurons[J].J Neurosci,2003,23(32):10388-10401.
[80 ]Haider B,Duque A,Hasenstaub AR,McCormick DA.Neocortical network activity in vivo is generated through a dynamic balance of excitation and inhibition[J].J Neurosci,2006,26(17):4535-4545.
[81 ]Schelter B,Timmer J,Schulze-Bonhage A.Seizure Prediction in Epilepsy:from Basic Mechanisms to Clinical Applications[M].New Jersey:John Wiley&Sons,2008.
[82 ]Klausberger T,Magill PJ,Márton LF,Roberts JD,Cobden PM,Buzsáki G,et al.Brain-state-and cell-type-specific firing of hippocampal interneurons in vivo[J].Nature,2003,421(6925):844-848.
[83 ]Shu Y,Hasenstaub A,McCormick DA.Turning on and off recurrent balanced cortical activity[J].Nature,2003,423(6937):288-293.
[84 ]Hasenstaub A,Shu Y,Haider B,Kraushaar U,Duque A,McCormick DA.Inhibitory postsynaptic potentials carry synchronized frequency information in active cortical networks[J].Neuron,2005,47(3):423-435.
[85 ]Lo¨rincz ML,Gunner D,Bao Y,Connelly WM,Isaac JT,Hughes SW,et al.A distinct class of slow(~0.2-2 Hz)intrinsically bursting layer 5 pyramidal neurons determines UP/DOWN state dynamics in the neocortex[J].J Neurosci,2015,35(14):5442-5458.
[86 ]McCormick DA,Shu Y,Hasenstaub A,Sanchez-Vives M,Badoual M,Bal T.Persistent cortical activity:mechanisms of generation and effects on neuronal excitability[J].Cereb Cortex,2003,13(11):1219-1231.
[87 ]Lerner TN,Ye L,Deisseroth K.Communication in neural circuits:tools,opportunities,and challenges[J].Cell,2016,164(6):1136-1150.
猜你喜欢
浙江大学学报(医学版)(2023年6期)2023-05-11 11:09:30
中小企业管理与科技(2022年19期)2022-12-22 13:04:21
赤峰学院学报·自然科学版(2022年9期)2022-10-17 04:45:14
浙江临床医学(2021年12期)2021-11-29 14:43:18
法大研究生(2020年2期)2020-01-19 01:42:24
中西医结合心血管病电子杂志(2018年28期)2018-11-19 11:04:54
中国疼痛医学杂志(2017年2期)2017-01-10 12:16:29
中国现代医学杂志(2015年26期)2015-12-23 11:04:20
吉林大学学报(医学版)(2015年1期)2015-12-17 07:47:27
江苏农业科学(2015年1期)2015-04-17 23:53:02
