
PI3K/AKT激动剂和抑制剂对巨噬细胞炎症反应的影响*
2017-03-09 03:42杨志文赖跃兴
胃肠病学 2017年2期
王 静 徐 萍# 杨志文 徐 凯 赖跃兴
上海交通大学附属第一人民医院松江分院消化内科1(201600) 药剂科2
PI3K/AKT激动剂和抑制剂对巨噬细胞炎症反应的影响*
王 静1徐 萍1#杨志文2徐 凯1赖跃兴1
上海交通大学附属第一人民医院松江分院消化内科1(201600) 药剂科2
背景:近年发现磷脂酰肌醇3激酶/丝氨酸-苏氨酸激酶(PI3K/AKT)在重度急性胰腺炎(SAP)的发病中发挥重要作用,但机制尚未明确。目的:探讨PI3K/AKT激动剂胰岛素样生长因子-Ⅰ(IGF-Ⅰ)和抑制剂wortmannin对巨噬细胞株RAW264.7 Toll样受体4(TLR4)信号通路的影响,阐明PI3K/AKT参与调节SAP炎症反应的作用机制。方法:分别以不同浓度脂多糖(LPS)、IGF-Ⅰ、wortmannin处理RAW264.7细胞,采用CCK-8实验检测细胞活性。RAW264.7细胞分为空白对照组(不予处理)、LPS组(LPS 1 μg/mL)、IGF-Ⅰ组(IGF-Ⅰ 100 ng/mL+LPS 1 μg/mL)、wortmannin组(wortmannin 100 nmol/L+LPS 1 μg/mL)和IGF-Ⅰ+wortmannin组(wortmannin 100 nmol/L+IGF-Ⅰ 100 ng/mL+LPS 1 μg/mL),采用ELISA法检测肿瘤坏死因子-α(TNF-α)、白细胞介素-6(IL-6)蛋白表达,采用real-time PCR检测TLR4、髓样分化因子88(MyD88)、AKT、PI3K、p38丝裂原活化蛋白激酶(p38MAPK)、核因子-κB(NF-κB)mRNA表达。结果:RAW264.7细胞经不同浓度LPS、IGF-Ⅰ、wortmannin处理后,各浓度组间细胞活性无明显差异(P>0.05)。LPS组、IGF-Ⅰ组、wortmannin组、IGF-Ⅰ+wortmannin组TNF-α、IL-6表达水平均较空白对照组显著升高(P<0.05);wortmannin组TNF-α、IL-6表达水平较LPS组和IGF-Ⅰ组显著降低(P<0.05);IGF-Ⅰ+wortmannin组 TNF-α、IL-6表达水平较IGF-Ⅰ组显著降低(P<0.05)。LPS组AKT、PI3K、TLR4及其下游分子MyD88、p38MAPK、NF-κB mRNA表达均显著高于空白对照组(P<0.05);IGF-Ⅰ组上述指标较LPS组进一步升高,差异有统计学意义(P<0.05);wortmannin组上述指标较LPS组和IGF-Ⅰ组显著降低(P<0.05);IGF-Ⅰ+wortmannin组上述指标显著高于wortmannin组(P<0.05),但较IGF-Ⅰ组显著降低(P<0.05)。结论:PI3K/AKT可能通过调节巨噬细胞中的TLR4及其下游分子影响促炎细胞因子表达,从而参与SAP炎症反应的发生。
胰腺炎; 磷酸肌醇3-激酶类; 蛋白质丝氨酸苏氨酸激酶; Toll样受体4; 胰岛素样生长因子 Ⅰ; Wortmannin; 肿瘤坏死因子α; 白细胞介素6
重度急性胰腺炎(severe acute pancreatitis, SAP)是病死率较高的消化系统疾病,其发病机制尚未完全阐明。本课题组前期研究[1]发现磷脂酰肌醇3激酶/丝氨酸-苏氨酸激酶(PI3K/AKT)参与调控SAP 大鼠胰腺组织促炎细胞因子的释放,可加重胰腺组织损伤;而PI3K/AKT抑制剂wortmannin可减轻胰腺组织损伤,提高SAP大鼠生存率。然而,PI3K/AKT参与SAP发病的具体机制尚未明确。已有研究[2]证实Toll样受体4(Toll-like receptor 4, TLR4)信号通路是促炎细胞因子释放的主要通路,刺激TLR4可激活核因子-κB(NF-κB)、p38丝裂原活化蛋白激酶(p38MAPK)信号通路,导致炎性因子释放。故本研究通过探讨PI3K/AKT激动剂胰岛素样生长因子-Ⅰ (insulin-like growth factor-Ⅰ, IGF-Ⅰ)和抑制剂wortmannin对小鼠巨噬细胞TLR4信号通路的影响,旨在阐明PI3K/AKT参与调节SAP炎症反应的作用机制。
材料与方法
一、细胞株、主要试剂和仪器
小鼠巨噬细胞株RAW264.7购自中国科学院上海细胞库。CCK-8试剂盒(日本同仁公司);脂多糖(LPS)、wortmannin(美国Sigma公司);IGF-Ⅰ、肿瘤坏死因子-α(TNF-α) ELISA试剂盒(美国Abcam公司);白细胞介素-6(IL-6) ELISA试剂盒(南京建成生物工程研究所);TRIzol试剂(美国Invitrogen公司);cDNA第一链合成试剂盒(美国Promega公司); PCR扩增试剂盒[生工生物工程(上海)股份有限公司];SYBR®PrimeScriptTMreal-time RT-PCR试剂盒(日本TaKaRa公司)。
二、方法
1. 细胞培养:RAW264.7细胞在37 ℃、5% CO2条件下,以含10%胎牛血清的RPMI1640培养基培养。
2. CCK-8实验检测LPS、IGF-Ⅰ和wortmannin对RAW264.7细胞活性的影响:取对数生长期RAW264.7细胞,以1×103~5×103/孔接种于96孔板培养过夜,分别以LPS、IGF-Ⅰ、wortmannin处理细胞,每组设6个浓度组,LPS、IGF-Ⅰ、wortmannin浓度分别为0、0.1、0.3、1、3、10 μg/mL;0、1、10、30、100、300 ng/mL;0、3、10、20、100、300 nmol/L。加入100 μL无血清DMEM培养基和10 μL CCK-8试剂,37 ℃培养1 h,于酶标仪450 nm波长处测定吸光度(A)值。
3. RAW264.7细胞分组处理:取对数生长期RAW264.7细胞,以5×103/mL接种于6孔板,设置空白对照组、LPS组、IGF-Ⅰ组、wortmannin组和IGF-Ⅰ+wortmannin组。空白对照组不予处理;LPS组以LPS(1 μg/mL)处理细胞;IGF-Ⅰ组以IGF-Ⅰ(100 ng/mL)预处理1 h后加入LPS(1 μg/mL)处理细胞;wortmannin组以wortmannin(100 nmol/L)预处理30 min后加入LPS(1 μg/mL)处理细胞;IGF-Ⅰ+wortmannin组以wortmannin(100 nmol/L)预处理30 min,以IGF-Ⅰ(100 ng/mL)预处理1 h,再以LPS(1 μg/mL)处理细胞。每孔细胞悬液终体积为1 mL,培养6 h后收集细胞。
4. ELISA法检测促炎细胞因子TNF-α、IL-6表达:取空白对照组、LPS组、IGF-Ⅰ组、wortmannin组和IGF-Ⅰ+wortmannin组细胞,检测TNF-α、IL-6表达,具体步骤参照相应ELISA试剂盒说明书进行。
5. Real-time PCR检测TLR4、髓样分化因子88(MyD88)、AKT、PI3K、p38MAPK、NF-κB mRNA表达:取空白对照组、LPS组、IGF-Ⅰ组、wortmannin组和IGF-Ⅰ+wortmannin组细胞,弃上清液,PBS漂洗,加入1 mL TRIzol试剂抽提细胞总RNA。采用cDNA第一链合成试剂盒合成cDNA,以之为模板,采用ABI 7300型定量PCR仪行real-time PCR扩增,检测TLR4、MyD88、AKT、PI3K、p38MAPK、NF-κB mRNA表达。引物由生工生物工程(上海)股份有限公司合成,以GAPDH作为内参照。TLR4引物上游:5’-CTA TGA ACA AAG GGT CTA TCA G-3’,下游:5’-AAG AAC AGC AAC CAC TAA AG-3’;MyD88引物上游:5’-CAC TCG CAG TTT GTT GGA TG-3’,下游:5’-TGT AAA GGC TTC TCG GAC TC-3’;AKT引物上游:5’-GGG CAC ATC AAG ATA ACG-3’,下游:5’-TGG TCC TGG TTG TAG AAG-3’;PI3K引物上游:5’-ATG CCA GAA AGG AGA ATG-3’,下游:5’-TGT TGG ACT CAG CAA TAC-3’;p38MAPK引物上游:5’-GTG TTC ACA CCC GCA AGG TC-3’,下游:5’-CGG TCA GCT TCT GGC ACT TC-3’;NF-κB引物上游:5’-CCC GAA ACT CAA CTT CTG-3’,下游:5’-ATC TGC CCT GAT GGT AAC-3’;GAPDH引物上游:5’-ATC ACT GCC ACC CAG AAG-3’,下游:5’-TCC ACG ACG GAC ACA TTG-3’。PCR反应条件:95 ℃ 10 min;95 ℃ 15 s,60 ℃ 45 s,40个循环。以2-△△Ct法分析目的基因相对表达量。
三、统计学分析
结 果
一、不同浓度LPS、IGF-Ⅰ、wortmannin对细胞活性的影响
LPS、IGF-Ⅰ、wortmannin各浓度组间RAW264.7细胞活性无明显差异(P>0.05)(图1)。最终选择以 1 μg/mL LPS、100 ng/mL IGF-Ⅰ和100 nmol/L wortmannin进行后续实验。
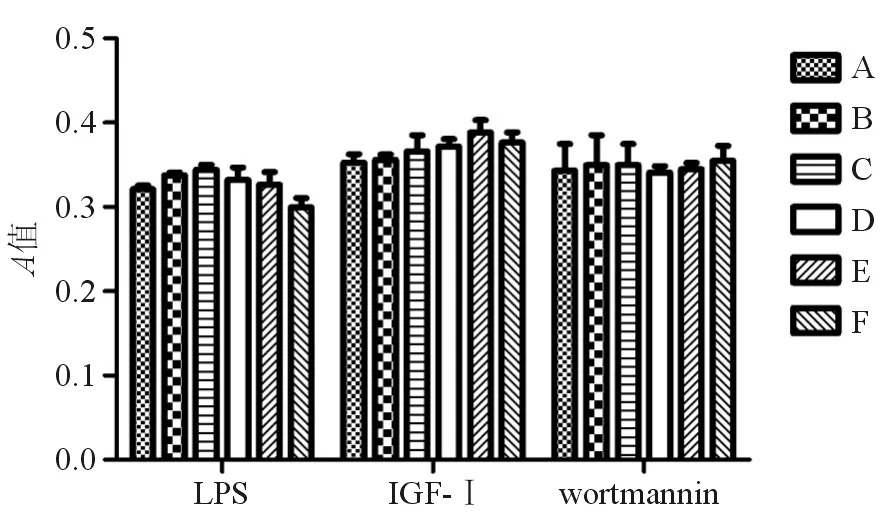
A-F:LPS浓度分别为0、0.1、0.3、1、3、10 μg/mL;IGF-Ⅰ浓度分别为0、1、10、30、100、300 ng/mL;wortmannin浓度分别为0、3、10、20、100、300 nmol/L
图1 不同浓度LPS、IGF-Ⅰ 和wortmannin对RAW264.7细胞 活性的影响
二、促炎细胞因子TNF-α、IL-6表达变化
LPS组RAW264.7细胞TNF-α、IL-6表达水平较空白对照组显著升高(P<0.05);IGF-Ⅰ组TNF-α、IL-6表达水平与空白对照组和LPS组相比均升高,与空白对照组间差异显著(P<0.05),与LPS组间差异则无统计学意义(P>0.05);Wortmannin组TNF-α、IL-6表达水平与空白对照组相比显著升高(P<0.05),与LPS组和IGF-Ⅰ组相比显著降低(P<0.05);IGF-Ⅰ+wortmannin组 TNF-α、IL-6表达水平与空白对照组相比显著升高(P<0.05),与LPS组相比有所降低,但差异无统计学意义(P>0.05),与IGF-Ⅰ组相比显著降低(P<0.05),与wortmannin组相比有所升高,但差异无统计学意义(P<0.05)(表1、图2)。
三、TLR4、MyD88、AKT、PI3K、p38MAPK、NF-κB mRNA表达
LPS组TLR4、MyD88、AKT、PI3K、p38MAPK、NF-κB mRNA表达水平均显著高于空白对照组(P<0.05)。IGF-Ⅰ 组上述指标均显著高于空白对照组和LPS组(P<0.05)。Wortmannin组上述指标显著高于空白对照组(P<0.05),与LPS组、IGF-Ⅰ组相比则显著降低(P<0.05)。IGF-Ⅰ+wortmannin组上述指标显著高于wortmannin组(P<0.05),但较IGF-Ⅰ组显著降低(P<0.05)(表2)。


组 别TNF⁃αIL⁃6空白对照组325.45±15.4653.64±3.06LPS组545.53±59.35137.80±12.30IGF⁃Ⅰ组570.29±51.24148.72±16.11wortmannin组490.20±25.97120.47±6.34IGF⁃Ⅰ+wortmannin组519.51±32.30131.92±9.58
讨 论
PI3K是生长因子超家族信号转导过程中的重要分子,可被多种细胞因子和理化因素激活。AKT主要负责由PI3K始动的生物信息传递。PI3K/AKT作为细胞内主要信号通路,在细胞代谢、细胞周期调控、细胞增殖、凋亡等多种生物学过程中发挥重要作用[3]。研究[4]表明PI3K/AKT通路可引起促炎细胞因子释放,参与SAP发病。本课题组前期研究[1]结果显示,SAP大鼠胰腺组织中磷酸化AKT表达增高,促炎细胞因子表达增加,予PI3K抑制剂wortmannin预处理后,AKT活性受抑,促炎细胞因子表达降低,胰腺组织病理学改变明显缓解,SAP大鼠生存率升高。本研究结果显示PI3K/AKT激动剂IGF-Ⅰ可促进LPS诱导巨噬细胞释放促炎细胞因子TNF-α、IL-6,而wortmannin作用于巨噬细胞后可抑制LPS引起的促炎细胞因子分泌,并可拮抗IGF-Ⅰ对促炎细胞因子的上调作用,与前期动物实验研究结果一致。
PI3K/AKT活化可导致细胞产生大量促炎细胞因子,此过程在炎症反应中发挥重要作用[5]。目前尚不明确PI3K/AKT促进促炎细胞因子产生的具体机制。TLR4信号通路是促炎细胞因子释放的主要通路, 活化的TLR4可通过MyD88等接头蛋白激活其下游NF-κB、 p38MAPK,产生大量炎性因子。研究[6-8]证实,SAP动物模型的胰腺、肠道以及肺炎症损伤均涉及TLR4信号通路激活。TLR4信号通路激活是SAP引发全身性炎症反应和脏器功能衰竭的重要病理生理学机制。本研究应用LPS刺激巨噬细胞激活TLR4信号通路,并分别以PI3K/AKT激动剂、抑制剂以及两者同时应用干预LPS引起的炎症反应,结果显示TLR4、 MyD88、p38MAPK、NF-κB表达随PI3K/AKT激动剂和抑制剂的干预而发生变化,提示PI3K/AKT与TLR4信号通路存在关联。国外一项研究[9]发现,尼古丁可减轻脓毒血症小鼠的炎症反应,此作用可能是通过影响PI3K/AKT信号通路、进而调节TLR4表达实现的,进一步提示PI3K/AKT与TLR4炎症信号通路密切相关。本研究结果显示IGF-Ⅰ可进一步激活LPS引起的炎症反应,活化TLR4信号通路,使TLR4、MyD88、p38MAPK、NF-κB表达上调,促炎细胞因子TNF-α、IL-6表达升高,而wortmannin可拮抗IGF-Ⅰ的作用;单独应用wortmannin作用于LPS刺激的巨噬细胞则可抑制LPS引起的TLR4信号通路活化,下调TLR4、MyD88、p38MAPK、NF-κB表达,并使促炎细胞因子表达降低。上述研究结果提示PI3K/AKT可能通过调节TLR4及其下游分子影响促炎细胞因子表达,从而参与SAP炎症反应的发生。

图2 各组RAW264.7细胞TNF-α、IL-6表达变化

组 别TLR4MyD88AKTPI3Kp38MAPKNF⁃κB空白对照组1.00±0.071.00±0.031.00±0.081.00±0.060.06±0.011.01±0.18LPS组7.06±0.793.26±0.084.05±0.314.89±0.150.25±0.033.92±0.27IGF⁃Ⅰ组9.11±0.365.97±0.565.13±0.346.04±0.590.35±0.015.28±0.31wortmannin组1.77±0.161.74±0.411.27±0.231.32±0.200.08±0.011.52±0.09IGF⁃Ⅰ+wortmannin组7.91±0.663.80±0.344.71±0.344.94±0.210.25±0.083.98±0.62
综上所述,本研究从细胞水平证实PI3K/AKT可通过影响TLR4信号通路调控p38MAPK、NF-κB活性,从而调节促炎细胞因子表达,参与SAP炎症反应。后期研究还需完善相关体内实验,进一步验证SAP中PI3K/AKT与TLR4信号通路的关系,为控制SAP炎症反应提供理论依据。
1 Xu P, Wang J, Yang ZW, et al. Regulatory roles of the PI3K/AKT signaling pathway in rats with severe acute pancreatitis[J]. PLoS One, 2013, 8 (11): e81767.
2 Zhao W, Ma G, Chen X. Lipopolysaccharide induced LOX-1 expression via TLR4/MyD88/ROS activated p38MAPK-NF-κB pathway[J]. Vascul Pharmacol, 2014, 63 (3): 162-172.
3 Cantley LC. The phosphoinositide 3-kinase pathway[J]. Science, 2002, 296 (5573): 1655-1657.
4 康新,王立志,王屹刚,等. 重症急性胰腺炎肺损伤时磷脂酰肌醇3激酶/蛋白激酶B信号转导通路的表达[J]. 中华医学杂志, 2010, 90 (11): 732-737.
5 Zhao M, Zhou A, Xu L, et al. The role of TLR4-mediated PTEN/PI3K/AKT/NF-κB signaling pathway in neuroin-flammation in hippocampal neurons[J]. Neuroscience, 2014, 269: 93-101.
6 Li Y, Zhou ZG, Zhang J, et al. Microcirculatory detection of Toll-like receptor 4 in rat pancreas and intestine[J]. Clin Hemorheol Microcirc, 2006, 34 (1-2): 213-219.
7 Sharif R, Dawra R, Wasiluk K, et al. Impact of toll-like receptor 4 on the severity of acute pancreatitis and pancreatitis-associated lung injury in mice[J]. Gut, 2009, 58 (6): 813-819.
8 Sawa H, Ueda T, Takeyama Y, et al. Role of toll-like receptor 4 in the pathophysiology of severe acute pancreatitis in mice[J]. Surg Today, 2007, 37 (10): 867-873.
9 Kim TH, Kim SJ, Lee SM. Stimulation of the α7 nicotinic acetylcholine receptor protects against sepsis by inhibiting Toll-like receptor via phosphoinositide 3-kinase activation[J]. J Infect Dis, 2014, 209 (10): 1668-1677.
(2016-07-01收稿;2016-07-20修回)
Effect of Agonist and Inhibitor of PI3K/AKT on Inflammatory Response in Macrophages
WANGJing1,XUPing1,YANGZhiwen2,XUKai1,LAIYuexing1.1
DepartmentofGastroenterology,2DepartmentofPharmacy,SongjiangHospitalAffiliatedtotheFirstPeople’sHospital,ShanghaiJiaotongUniversity,Shanghai(201600)
XU Ping, Email: sjzxxp@yeah.net
Pancreatitis; Phosphatidylinositol 3-Kinases; Protein-Serine-Threonine Kinases; Toll-Like Receptor 4; Insulin-Like Growth Factor Ⅰ; Wortmannin; Tumor Necrosis Factor-alpha; Interleukin-6
10.3969/j.issn.1008-7125.2017.02.004
上海市松江区卫计委医学领先专业项目(201358)
#本文通信作者,Email: sjzxxp@yeah.net
Background: Phosphoinositide 3-kinase/serine-threonine kinase (PI3K/AKT) has been found playing an important role in the pathogenesis of severe acute pancreatitis (SAP) in recent years, but the underlying mechanism has not been clarified. Aims: To investigate the role of PI3K/AKT in regulating the inflammatory response in SAP by evaluating the effect of insulin-like growth factor-Ⅰ (IGF-Ⅰ) and wortmannin, the agonist and inhibitor of PI3K/AKT on Toll-like receptor 4 (TLR4) signaling pathway in macrophage cell line RAW264.7. Methods: RAW264.7 cells were treated with different concentrations of lipopolysaccharide (LPS), IGF-Ⅰ and wortmannin, respectively, and cell viability was determined by CCK-8 assay. RAW264.7 cells were divided into blank control group (no treatment), LPS group (LPS 1 μg/mL), IGF-Ⅰ group (IGF-Ⅰ 100 ng/mL+LPS 1 μg/mL), wortmannin group (wortmannin 100 nmol/L+LPS 1 μg/mL) and IGF-Ⅰ+wortmannin group (wortmannin 100 nmol/L+IGF-Ⅰ 100 ng/mL+LPS 1 μg/mL). Protein expressions of tumor necrosis factor-α (TNF-α) and interleukin-6 (IL-6) were detected by ELISA; mRNA expressions of TLR4, myeloid differentiation factor 88 (MyD88), AKT, PI3K, p38 mitogen-activated protein kinase (p38MAPK) and nuclear factor-κB (NF-κB) were determined by real-time PCR. Results: After treated with LPS, IGF-Ⅰ and wortmannin, respectively, no differences in cell viability of RAW264.7 cells were found between different concentrations groups (P>0.05). Protein expressions of TNF-α and IL-6 in LPS, IGF-Ⅰ, wortmannin and IGF-Ⅰ+wortmannin groups were significantly higher than those in blank control group (P<0.05). Protein expressions of TNF-α and IL-6 in wortmannin group were significantly lower than those in LPS and IGF-Ⅰ groups (P<0.05), and those in IGF-Ⅰ+wortmannin group were significantly lower than those in IGF-Ⅰ group (P<0.05). In LPS group, mRNA expressions of AKT and PI3K as well as TLR4 and its downstream molecules MyD88, p38MAPK and NF-κB were significantly higher than those in blank control group (P<0.05). Expressions of all above-mentioned mRNAs in IGF-Ⅰ group were further increased and significantly higher than those in LPS group (P<0.05). Expressions of all above-mentioned mRNAs in wortmannin group were significantly lower than those in LPS and IGF-Ⅰ groups (P<0.05), and those in IGF-Ⅰ+wortmannin group were significantly higher than those in wortmannin group (P<0.05), but significantly lower than those in IGF-Ⅰ group (P<0.05). Conclusions: PI3K/AKT might regulate TLR4 signaling pathway and its downstream molecules in macrophages, thereby affects the expressions of inflammatory cytokines and being involved in the pathogenesis of inflammatory response in SAP.
猜你喜欢
中日友好医院学报(2022年4期)2022-10-15
天津医科大学学报(2021年3期)2021-07-21
世界科学技术-中医药现代化(2021年12期)2021-04-19
看世界·学术下半月(2020年7期)2020-09-10
中成药(2018年10期)2018-10-26
天然产物研究与开发(2018年6期)2018-07-09
广东农业科学(2017年10期)2018-01-25
中国医学创新(2017年22期)2017-11-15
中国医药生物技术(2015年4期)2015-12-26
医学研究杂志(2015年5期)2015-06-10
