
中国红菇属2个新记录种
2017-03-07 11:27姜旭萌李杨坤梁俊峰伍建榕
福建农林大学学报(自然科学版) 2017年1期
姜旭萌, 李杨坤, 梁俊峰, 伍建榕
(1.西南林业大学林学院,云南 昆明 650224;2.中国林业科学研究院热带林业研究所,广东 广州 510520)
中国红菇属2个新记录种
姜旭萌1,2, 李杨坤2, 梁俊峰2, 伍建榕1
(1.西南林业大学林学院,云南 昆明 650224;2.中国林业科学研究院热带林业研究所,广东 广州 510520)
报道了中国发现的红菇属2个新记录种辛德红菇(RussulathindiiK. Das & S. L. Miller)和甜汁红菇(RussulasapineaSarnari Boll.).对采自我国的这2个记录种标本进行宏观和微观形态研究,并对其重要显微特征进行绘图.同时,采用分子生物学方法,通过系统发育分析探讨其属内分类位置.
红菇属; 新记录种; 形态学; 系统发育
红菇属(RussulaPers.)隶属于担子菌门(Basidiomycota)伞菌纲(Agaricomycetes)红菇目(Russulales)红菇科(Russulaceae)[1].该属真菌多为外生菌根菌,能与多种植物共生形成菌根,常发生于夏秋两季针叶林、阔叶林和混交林中的稠树林,在森林生态系统中具有非常重要的作用[2-5].红菇属真菌具有重要的经济价值,比如在中国南部、东南亚一带广受欢迎的灰肉红菇复合群(R.griseocarnosacomplex),已经被证实具有防癌、补血等多种食疗保健功能;但也有一些红菇种类有毒,比如毒红菇[R.emetica(Schaeff.) Pers.]、亚稀褶红菇(R.subnigricans)等[6-8].
红菇属真菌分布广泛,大约有750余种[9],我国已报道大约有160余种[10-12].本文通过对我国红菇的形态及分子系统学的研究,发现2个新记录种,并进行描述报道,为进一步研究我国红菇属真菌多样性提供参考.
1 材料与方法
1.1 供试标本
研究材料采集于湖北省、四川省、西藏自治区,标本经标本干燥器50 ℃干燥后保存于中国林业科学院热带林业研究所标本馆.
1.2 方法
1.2.1 形态学研究 标本的宏观外部形态特征来自采集者野外采集新鲜子实体的记录.显微特征则在OLYMPUS BX51显微镜下进行观察;担孢子通过JSM-6510 LV扫描电镜观察.为便于显微观察研究,从干燥标本组织局部取样,置于5%(体积分数) KOH 溶液中复水;再用1%刚果红试剂染色,在明视野或相差视野下观察、记录和绘图.用梅氏试剂测定其担孢子壁是否为拟糊精质,用甲酚蓝测定担孢子壁是否变色.担子测量值来源于具有小梗的成熟担子,小梗长度不计.担孢子只测量其侧面的长度和宽度,担孢子的侧生小尖和纹饰不记入内[13];其结构大小用明美数码成像系统测量.
从研究标本的成熟担子果上,随机测量20个成熟担孢子.担孢子的长度或宽度用(a)b-c(d)表示,95%的测量数值落在b-c,a、d分别为测量数据中的最小值和最大值;担孢子的长宽比用Q表示,Qav表示担孢子长宽比的算术平均数与标准差.[n/m/p]表示测量p份标本的m个子实体上的n个担孢子.
1.2.2 分子系统学研究 DNA采用改良的CTAB法提取[14].内转录间隔区序列(ITS)用ITS5和ITS4引物进行PCR扩增[15].PCR体系:2.5 μL 10×PCR Buffer、2.5 μL dNTP(200 μmol·L-1)、ITS5(5′-GGAAGTAAAAGTCGTAACAAGG-3′)和ITS4(5′-TCCTCCGCTTATTGATATGC-3′)引物各1 μL(5 μmol·L-1)、17 μL ddH2O、DNA模板1 μL.PCR产物由北京奥科鼎盛生物科技有限公司用 Bioteke′s Purification Kit试剂盒纯化,通过ABI3730 DNA测序仪测序.新记录种序列采用上述序列测得.其它序列是在前人研究的基础上,利用Genbank数据库中的Blast软件筛选获得.对汇总后的序列用Bioedit v7.0.9进行多序列比对,并用Gblocks v0.91b进行保守区选择,最后进行人工微调[16,17].分子系统发育树通过 RAxML7.2.6和Mrbayes分别采用最大似然法和贝叶斯推断法完成.核苷酸替代模型用MrModeltest 2.3进行筛选[18].同时选用已经报道的烟色红菇[R.adusta(Pers.) Fr.]和黑红菇[R.nigricans(Bull.) Fr.]作为外类群[19].
2 形态学与系统发育分析
2.1 辛德红菇RussulathindiiK. Das & S. L. Miller, Mycosphere, 2014, 5(5): 618
子实体中型(图1A).菌盖直径3.5~8 cm,初时半球形,中间凸,后渐渐平凸,成熟时平展或向上呈浅漏斗状;表面光滑,潮湿时呈粘质或发粘;中央血红色与浅黄色混合色调,边缘部分颜色呈血红色至浅灰红色,略带浅黄褐色;成熟时中央偶带珊瑚色,干后带淡绿色;菌盖边缘有明显条纹,具槽,幼时内卷,成熟时下弯.菌褶密,乳白色,成熟时带淡黄色调,近柄处分叉,直生至微弯生,具有3~4层小菌褶,受伤不变色,干后变暗黄色.菌柄(5~8.8) cm×(1~2.3) cm,中生,初髓状后中空,近棒状,有时基部稍窄;表面具脉络状条纹,湿时粘,表面淡红色至酒红色;幼时基部白色,成熟后渐渐变为橘黄色或淡黄色至淡橙色.菌肉结实,菌盖薄,淡黄白色,遇5%(体积分数)KOH溶液不变色.味道略苦辣.气味淡香.孢子印微淡黄色.

图1 辛德红菇子实体(A)和担孢子(B);甜汁红菇子实体(C)和担孢子(D)Fig.1 Sporophore(A) and basidiospores(B) of R.thindii; sporophore(C) and basidiospores(D) of R.sapinea
担孢子(图1B)[40/2/2](6.5)6.8~8.6(9.9)μm×(6.0)6.2~7.0(7.3)μm (Qav=1.17±0.07),近球形至宽椭球形,无色;表面纹饰淀粉质,疣突高达1.2 μm,大多呈孤立,近圆柱形,之间较少或无连线;脐上区点斑非淀粉质.担子(图2A)(28~56.5) μm×(9.0~12.5) μm,尺寸变化较大,粗棒状至棒状的2~4个担子小梗向一侧肿胀,成熟的小梗长达8.0 μm.侧生囊状体(图2B)(41.5~123) μm×(12~15) μm,丰富,近圆柱形、棍棒状或纺锤形,顶端圆钝,少或无乳头状短尖,薄壁,含有丰富的晶体或纤维状内容物,遇硫酸香草醛溶液变灰黑色.褶缘囊状体(图2C)(41~75) μm×(7.5~10.5) μm,丰富,圆柱形或纺锤形,薄壁,顶端钝圆,少或无念珠状附属物;内容物丰富,遇硫酸香草醛溶液变灰黑色.子实层厚50 μm,层状菌髓由无数球状细胞(10.5~56.5) μm×(8~48.5) μm和常分支具隔的菌丝(2.5~5 μm)组成.盖表皮(图2D)由透明的菌丝和囊状体组成;菌丝栅栏状排列,末端向外延伸,钝圆,3~5.5 μm宽,常分支,具隔;盖表囊状体(23.5~50) μm×(5~8) μm,圆柱状或棍棒状,顶端钝圆;遇硫酸香草醛溶液变灰黑色.菌柄表皮菌丝宽40 μm,平行排列;柄生囊状体缺失.锁状联合缺失.
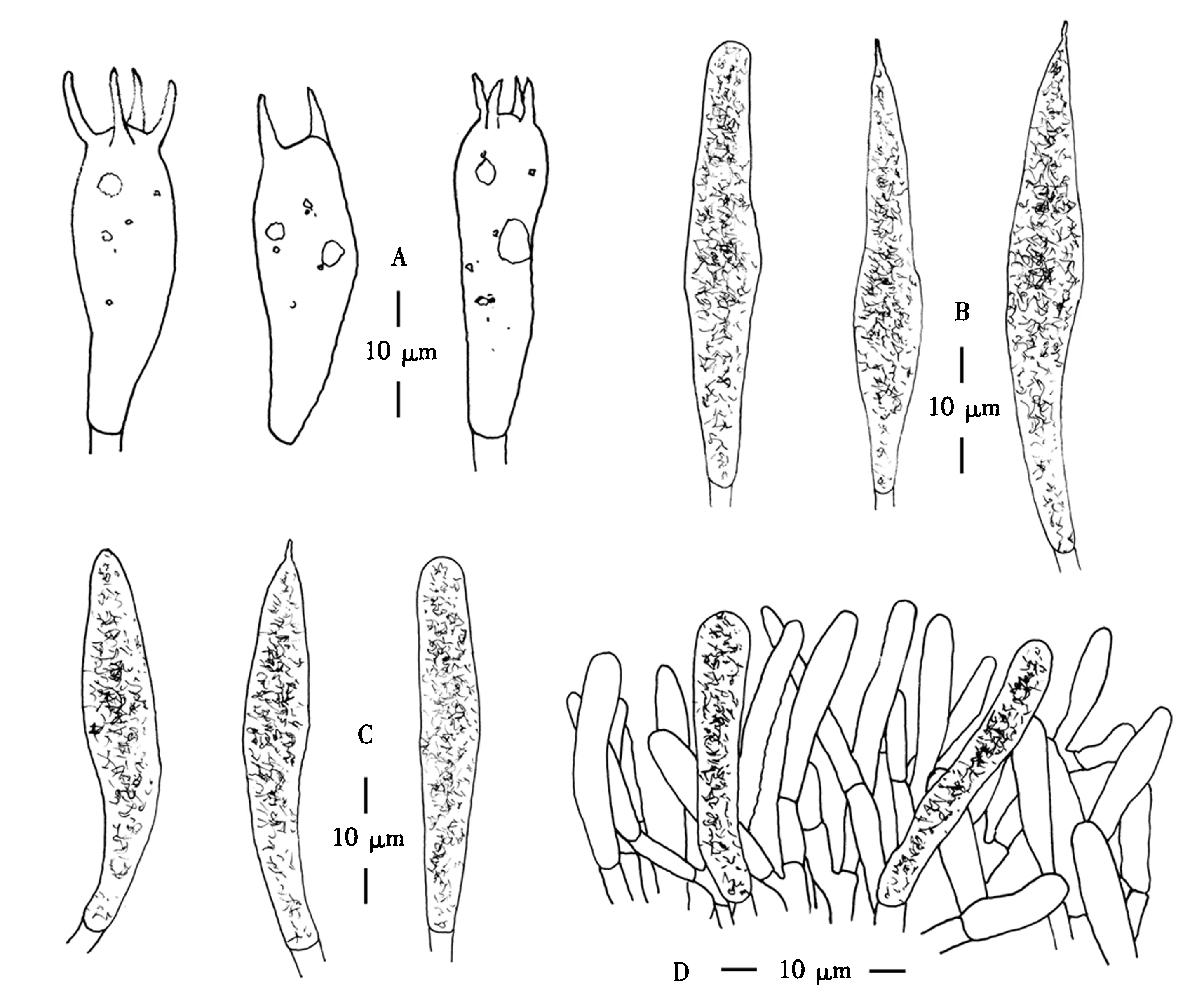
A.担子;B.侧生囊状体;C.褶缘囊状体;D.盖表菌丝和盖表囊状体.图2 辛德红菇显微结构 Fig.2 Microscopical structures of R.thindii
生长季节为夏秋两季,生境为冷杉针叶林下.
模式产地:印度锡金[20].
分布:印度、中国(湖北、西藏).
研究标本:湖北神农架林区木鱼镇神龙顶青云梯冷杉林,海拔1 800 m,2015年8月7日,李杨坤60(RITF2712);西藏林芝地区墨脱县墨脱公路冷杉林,2014年7月3日,王苗 T25044(RITF2722).
辛德红菇主要鉴别特征:菌盖红色粘质,菌褶直生略弯生,菌柄带微红至紫红色调,味道辣,淡黄色孢子印,与冷杉共生.综合这些特征,辛德红菇(R.thindii)应属于红菇属下的红菇亚属(subg.RussulaRomagnesi)辣味组(sect.SardoninaeSinger)[21].
采自我国的标本序列通过GenBank数据库中Blast序列比对,与R.thindii序列同源性达99%.利用最大似然法和贝叶斯推断法构建系统发育树(图3),结果显示采集自我国的标本与R.thindii聚在一个分支上,靴带支持率90%,后验概率0.98,并与其它辣味组红菇明显聚集在一起.

图3 系统发育分析Fig.3 Maximum likelihood analysis of R.thindii and R. sapinea
与模式标本相比,采集自西藏的标本(RITF2722)大多数担子呈粗棒状,少量棒状;而采自湖北的标本(RITF2712)担子较大,多为棒状,但其它特征均与模式标本较一致.综合形态与分子证据,认为这两份标本应该属于R.thindii.
由图3可知,辛德红菇与凯莱红菇(R.queletiiFr.)和血红菇[(R.sanguinea(Bull.) Fr.)]近缘,且辛德红菇与凯莱红菇具有相对较近的亲缘关系,聚在一支的靴带支持率为78%.辛德红菇与凯莱红菇的主要区别在于凯莱红菇菌柄较短,菌肉带有很强的水果香味,缺乏苦辣味道,孢子表面疣突多,连线呈不完整或者近完整网状[22].与血红菇的区别在于血红菇菌盖呈血红色,并具有较辣的菌肉,孢子印颜色呈奶油色,脐上区具有明显的淀粉质点[6,22].
2.2 甜汁红菇RussulasapineaSarnari Boll. Assoc. Micol. Ecol. Romana, 1994, 12(33): 21
子实体中型(图1C).菌盖直径3~6.5 cm,初时半球形,中央凸起,成熟后平展,不对称,中央下凹,表面不平,湿润时略粘,子实体干后柔软;菌盖颜色变化多样,淡粉橙色至紫丁香颜色,成熟时中央常带有橄榄色和土黄混合色;菌盖边缘幼时稍内卷,易开裂,表面有条纹,具槽.菌褶稍密,直生或近离生,等长,无小菌褶,近柄处易分叉,易碎,波状,中间较宽,乳白色,边缘淡白色,干后泛淡橙黄色.菌柄(3~7) cm×(1~1.8) cm,白色,不规则圆柱状,向基部逐渐变大,无菌环菌托,表面较有韧性,具微细皱纹,初内实,后内部松软,湿时水浸状.菌肉白色,干后较松软,老后略变锈赭色.气味水果腐烂味,味道略甜.孢子印橙黄色或赭黄色.
担孢子(图1D)[20/1/1]8.5~10.5 × 6.5~9.0 μm(Qav=1.12±0.09),椭球形或近椭球形,表面具淀粉质疣突,高约1.2 μm,疣突圆柱形,多孤立,有时相连成不规则连线或成短脊;脐上区点斑淀粉质.担子(图4A)(32.5~64) μm×(12~16) μm,棒状,具有2~4个小梗,向一侧肿胀,成熟的小梗长达10 μm.侧生囊状体(图4B)(60~93) μm×(9.0~13.5) μm,不规则的棒状或梭状,数量丰富,顶端有乳头状短尖或圆钝,薄壁,含有丰富的晶体或纤维状内容物,遇硫酸香草醛溶液变灰黑色.褶缘囊状体(图4C)(45~75) μm×(9.0~12.5) μm,近梭形,数量较少,顶端常具乳状突起或念珠状附属物,薄壁,含有丰富的晶体或纤维状内容物.盖表皮(图4D)菌丝3.0~4.5 μm,近匍匐排列,有分隔,末端向外延伸,末端细胞顶部钝圆,末端细胞(15~30) μm×(3.0~4.0) μm,近细棒状,近末端处常有分支.盖表囊状体(36~61.5) μm×(5.5~9.0) μm,棒状至细棒状,有隔,顶端圆钝,含纤维状内容物.菌柄表皮菌丝宽2.5~5.0 μm,有隔,无色透明,菌柄囊状体未见.锁状联合缺失.
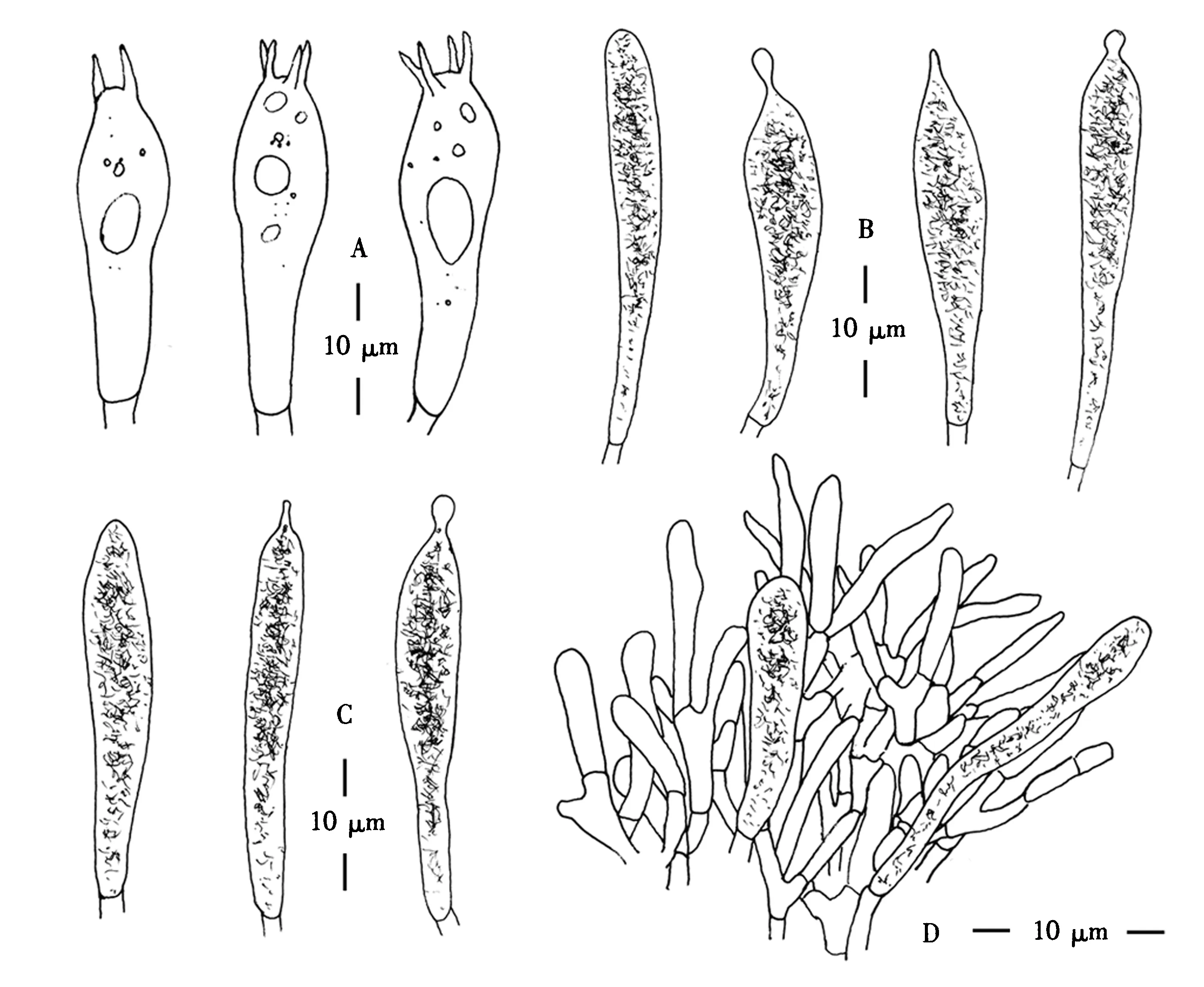
A.担子;B.侧生囊状体;C.褶缘囊状体;D.盖表菌丝和盖表囊状体.图4 甜汁红菇的显微结构Fig.4 Microscopical structures of R.sapinea
生长季节为夏秋两季,生境为云杉、落叶松针林或混交林.
模式产地:意大利[6].
分布:欧洲(挪威、芬兰等)[21]、中国(四川).
研究标本:四川省阿坝藏族羌族自治州理县米亚罗风景区,2014年6月24日,赵琦 T25439(RITF2700).
甜汁红菇的主要鉴别特征为菌盖颜色近粉红色至淡紫丁香色,菌盖中央经常橄榄色,菌褶等长,菌柄较肥厚,味道甜,孢子印赭黄色.综合这些特征,甜汁红菇应属于娇弱亚属(Sect.TenellulaRomagnesi)落叶松亚组(Subsect.LaricinaeRomagnesi)[21,22].
采自我国的标本序列通过Genbank数据库中Blast序列比对,与R.sapinea序列同源性达99%.系统发育树(图3)显示该标本与R.sapinea聚在一个分支上,靴带支持率92%,后验概率0.98,并与其它落叶松亚组明显聚在一起.
甜汁红菇和臭味红菇[R.nauseosa(Pers.) Fr.]近缘, 靴带支持率81%,后验概率0.99.甜汁红菇与臭味红菇的区别在于后者具有菌肉不变色,菌盖颜色多偏向红色,味道中和,菌肉易碎,孢子疣突更多成孤立状态[21].与美红菇(R.puellarisFr.)在外观上相似,与美红菇的区别在于美红菇具有较小的孢子,且上面纹饰连线较少[21].
3 小结
中国发现的辛德红菇和甜汁红菇属2个新记录种.采自西藏的标本(RITF2722)和湖北的标本(RITF2712)与印度报道的辛德红菇R.thindii序列同源性高达99%,西藏的标本与辛德红菇模式标本形态特征基本一致,而湖北的标本担子较模式标本略大,其它特征均一致,故将上述两标本鉴定为辛德红菇.采集自四川的标本(RITF2700)与欧洲报道的甜汁红菇R.sapinea序列同源性达99%,该标本囊状体数量较模式标本丰富,其他特征均与模式标本一致,综合形态与分子证据,将该标本鉴定为甜汁红菇.
致谢:感谢昆明理工大学王苗同学提供标本照片和采集记录,也感谢中科院微生物研究所李国杰博士为本文的修改提供建议.
[1] PERSOON C H. Descriptiones tam novorum quan notabilium fungorum[J]. Observations Mycologicae, 1796,1:1-221.
[2] BUYCK B, THOEN D, WATLING R. Ectomycorrhizal fungi of the Guinea-Congo region[J]. Proceedings of the Royal Society of Edinburgh, 1996,104: 313-333.
[3] 许旭萍,李惠珍,黄德鑫.红菇生态的研究[J].中国食用菌,2001,20(2):25-27.
[4] 张颖,徐建萍,徐远钊,等.滇中半湿润常绿阔叶林下红菇科真菌的多样性及生态学研究[J].广东农业科学,2014,41(13):152-159.
[5] 陈宇航,陈政明,林国华.远东疣柄牛肝菌仿生栽培[J].福建农林大学学报(自然科学版),2002,31(4):532-535.
[6] 王青云,石木标.中国红菇的研究现状与展望[J].中国食用菌,2004,23(4):10-12.
[7] 李国杰,李赛飞,文华安.中国红菇属物种资源经济价值[J].食用菌学报(增刊),2010:155-160.
[8] 徐惠龙,杨志坚,郑金贵.3种食用菌对高脂血症大鼠脂质代谢的影响[J].福建农林大学学报(自然科学版),2014,43(4):430-433.
[9] KIRK P M, CANNON P F, MINTER D W, et al. Dictionary of the fungi. 10th edition[M]. Oxon: CAB International, 2008.
[10] 宋斌,李泰辉,吴兴亮,等.中国红菇属种类及其分布[J].菌物研究,2007,5(1):20-42.
[11] 李国杰,文华安.中国红菇属分类研究进展[J].菌物学报,2009,28(2):303-309.
[12] 李国杰,李赛飞,赵东,等.红菇属研究进展[J].菌物学报,2015,34(5):821-848.
[13] 梁俊峰,周玲玲,仲崇禄,等.中国环柄菇属一新记录种——库勒环柄菇[J].福建农林大学学报(自然科学版),2010,39(6):622-624.
[14] 周玲玲,梁俊峰.大型真菌DNA提取方法的改进[J].广东林业科技,2011,27(1):13-16.
[15] WHITE T J, BRUNS T, LEE S, et al. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics[J]. PCR Protocols, 1990, 18: 315-322.
[16] HALL T A. BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT[J]. Nucleic Acids Symposium Series,1999,41: 95-98.
[17] CASTRESANA J. Selection of conserved blocks from multiple alignments for their use in phylogenetic analysis[J]. Molecular Biology and Evolution, 2000,17:540-552.
[18] NYLANDER J A A. MrModeltest v2. program distributed by the author[C]. Uppsala University:Evolutionary Biology Centre,2004.
[19] MILLER S L, BUYCK B. Molecular phylogeny of the genusRussulain Europe with a comparison of modern infrageneric classifications[J]. Mycological Research, 2002,106(3):259-276.
[20] DAS K, DOWIE N J, LI G J, et al. Two new species ofRussula(Russulales) from India[J]. Mycosphere, 2014,5(5):612-622.
[21] SARNARI M. Monografia illustrata del genereRussulain Europa[M]. Trento: AMB, 1998.
[22] SARNARI M. Monografia illustrata del genereRussulain Europa[M]. Trento: AMB, 2005.
(责任编辑:叶济蓉)
Two newRussulaspecies in China
JIANG Xumeng1,2, LI Yangkun2, LIANG Junfeng2, WU Jianrong1
(1.College of Forestry, Southwest Forestry University, Kunming, Yunnan 650224, China; 2.Research Institute of Tropical Forestry, Chinese Academy of Forestry, Guangzhou, Guangdong 510520, China)
Two new Chinese records,RussulathindiiK. Das & S. L. Miller andR.sapineaSarnari Boll, are reported. The detailed descriptions of micro- and macro-morphological features of the two species based on Chinese specimens are provided, and illustrations on important microscopical characteristics of both species are offered. Meanwhile, molecular biology method was used to discuss the phylogenetic position of the two species.
Russula; new record species; morphology; phylogeny
2016-06-18
2016-09-17
国家自然科学基金(31570544)和广东省自然科学基金项目(2014A030313727).
姜旭萌(1991-),女,硕士研究生.研究方向:资源微生物.通讯作者梁俊峰(1971-),男,研究员,博士生导师.研究方向:森林微生物资源遗传多样性,Email:jfliang2000@163.com
Q949.329
A
1671-5470(2017)01-0103-06
10.13323/j.cnki.j.fafu(nat.sci.).2017.01.016
猜你喜欢
中国食用菌(2022年2期)2022-11-22
现代园艺(2021年1期)2021-12-07
检验医学与临床(2021年22期)2021-11-26
作文周刊·小学三年级版(2020年40期)2020-10-15
农家之友(2020年9期)2020-04-21
渔业致富指南(2016年12期)2016-11-11
兵器装备工程学报(2015年5期)2015-12-23
科学大众(中学)(2015年9期)2015-10-12
少儿科学周刊·少年版(2015年3期)2015-07-07
华东师范大学学报(自然科学版)(2014年6期)2014-02-27
