
不同年龄香果树种子雨和种子库及其更新特征
2017-01-03 01:39郭连金徐卫红房会普肖志鹏田玉清黄厦华殷崇敏胡根秀
西北植物学报 2016年11期
郭连金, 徐卫红, 房会普, 肖志鹏, 田玉清,3,黄厦华, 殷崇敏, 胡根秀
(1 上饶师范学院,江西上饶 334000; 2 宝天曼国家级自然保护区管理局,河南南阳 474350; 3 中国科学院昆明植物研究所, 昆明 650204)
不同年龄香果树种子雨和种子库及其更新特征
郭连金1, 徐卫红1, 房会普2, 肖志鹏1, 田玉清1,3,黄厦华1, 殷崇敏1, 胡根秀1
(1 上饶师范学院,江西上饶 334000; 2 宝天曼国家级自然保护区管理局,河南南阳 474350; 3 中国科学院昆明植物研究所, 昆明 650204)
对宝天曼自然保护区香果树种子雨、土壤种子库进行了观测,并进行不同微生境播种实验,研究其种子萌发及幼苗生长动态。结果表明:(1)不同龄级香果树种子雨持续时间及其高峰期有所不同,种子雨强度存在极显著差异。(2)20~50龄级香果树的种子饱满率、土壤种子库中种子密度均显著低于其他龄级,但其不同类型种子的水平和垂直分布规律一致;大约80%的香果树种子在其萌发前消失,剩余种子中以霉烂种子密度最高(9.81粒·m-2),饱满种子密度最低(1.94粒·m-2)。(3)野外育苗实验表明,不同龄级香果树所产饱满种子的萌发率及幼苗存活率差异不显著,其种子萌发率约为16.93%,但仅有3.86%的幼苗寿命超过5个月;不同微生境对香果树幼苗存活率产生显著影响,林窗是其最适宜微生境。研究认为,人工采集香果树种子,于次年4月份在原生境下播种;清理母树冠下和冠缘的地被物,增加林缘空地地被物覆盖度,以减少香果树种子的损失,改善香果树种子萌发和幼苗生长的光照条件,以利于幼苗根系的生长,促进香果树的自然更新。
香果树; 种子质量; 种子雨;种子库;幼苗更新
种子雨是植物种子成熟后从母体降落下来在特定时间和空间上的数量[1],是研究物种生态适应及种群对策的基础[2],而存在于土壤覆盖物及土壤中的全部种子总和的土壤种子库[3]则是影响植物群落天然更新能力和方向的关键[4,5],其时空分布可以反映地上植株的过去与未来[6],两者是植被更新和恢复的物质基础[2]。幼苗是植物个体生长最为脆弱、对环境变化最为敏感的时期[7]。幼苗的定居及其数量动态决定着种群的格局和命运,是植物种群维持和实现其自然更新的一个重要阶段[8]。种子雨、种子库以及幼苗定居和数量动态是植物种群自然更新的重要组成部分[9]。植物种群自然更新如果成功,可使该种群增殖、扩展、延续,并维持其种群稳定[8],但如果失败,则使其种群衰退,甚至消失[10]。珍稀濒危植物种群都存在自然更新失败的问题[11],研究珍稀濒危植物的自然更新问题,即研究其种子雨、种子库以及幼苗定居等问题,有利于珍稀濒危植物的保护及其濒危原因的揭示,它不仅可阐明其种群消失的原因,了解物种自然更新的脆弱环节,并可为制定珍稀濒危植物保护措施以及持续利用策略提供理论依据。
香果树(Emmenopteryshenryi)属茜草科单种属植物,为落叶高大乔木,中国特有的国家Ⅱ级重点保护植物,是研究茜草科系统发育、形态演化及中国植物地理区系的重要材料[12]。有研究表明香果树具有花多果少的格局[13,14],但由于每果中种子500~800粒,其有性生殖策略以多种子取胜[15]。虽然其种子寿命不足1年,萌发率低,但产种量大,在适宜的环境下仍可萌发[13];原生境中香果树幼苗较少[16],其分布格局为聚集分布,其生长受光照和大气温湿度影响较大[17]。目前,尽管部分学者对香果树种子特征、萌发以及幼苗生长等进行了大量研究[13,15,17-22],但均为实验室或大棚实验,原生境中香果树种子雨、土壤种子库、种子萌发以及幼苗定居等方面的研究未见报道,而这正是揭示香果树种群濒危的关键。本研究以宝天曼自然保护区香果树为研究对象,通过研究不同年龄香果树种子雨的时空格局、土壤种子库分布格局、种子质量变化以及实生苗的数量变化动态等内容,阐明香果树种群自然更新的过程,寻找其更新脆弱环节及影响因素,为其种群恢复与有效经营管理提供依据。
1 研究地自然概况
研究地设在伏牛山宝天曼自然保护区(33°25′-33°33′ N,110°53′-112°E),该区位于伏牛山南麓,秦岭东段,年平均气温9~9.4 ℃,年平均降雨量893.2 mm,年蒸发量991.6 mm,相对湿度51.2%,≥10 ℃的年积温4 200~4 900 ℃,无霜期低山区227 d,土壤类型主要为黄棕壤、棕壤和暗棕壤,植被类型以温带落叶阔叶林为主[23]。香果树在研究地主要分布于落叶阔叶林中,其伴生种主要有黑桦(Betulachinensis)、栓皮栎(Quercusvariabilis)、紫荆(Cercischinensis)、小叶青冈(Cyclobalanopsismyrsinifolia)、大叶朴(Celtiskoraiensis)、山胡椒(Linderaglauca)、金钱草(Lysimachiachristinae)和三脉紫菀(Asterageratoides)等。
2 研究方法
2.1 样地设置及生境调查
在河南宝天曼自然保护区香果树分布区选择典型的4个生境(A、B、C、D)的香果树种群为研究对象,分别设置4个面积为100 m×1 000 m的样地进行群落生态学调查(表1)。调查内容包括:
① 生境:记录海拔、坡向、坡位等,其中土壤含水量采用便携式土壤含水量速测仪(TRIME-TD RZ)进行测定,用照度计(ZDH-10)测定林内地面光照,用通风干湿度温度计(DMH2)记录林内的温湿度(选择前两天无雨天气,林内光照、温湿度等的测定于每日10:00、13:00、15:00进行)。
② 群落学特征:树种组成、高度、盖度等。
③ 样地内香果树母株调查:记录其数量、胸径、树高等。
④ 母树年龄的确定:于2014年8月1日,在4个样地中分别选择10株胸径大小不等的香果树,共计40株,利用瑞典生长锥在其主干南侧离地1.3 m处进行垂直钻木,待锥筒先端钻过母树树干的髓心后,取锥筒中的木条并查数其年轮,以此作为母树的年龄,对香果树的年龄(y)和胸径(x)进行拟合,获得模型y=0.013 4x2+0.272x+17.264(R2=0.983,P=0.000,n=80),该多项式模型拟合度较高,可以作为判别香果树年龄的基本模型。
本研究选择S1(20~50 a)、S2(50~80 a)、S3(80~110 a)和S4(110~140 a)4个年龄级,每个年龄级选择12株母树(表2),即每个样地3株,其中1株用于采集种子,用于香果树的种子质量和野外萌发实验,另外2株用于种子雨和种子库研究。为避免香果树母树间种子雨的影响,本研究所标记的用于种子雨和种子库研究的母树生存环境相对开阔,其周围200 m内没有其他母树处于繁殖期。
2.2 种子雨收集
2014 年9月底,在研究种子雨和种子库的8棵香果树母树附近选择4个种子雨分布的微生境——冠下(M1)、冠缘(M2)、林窗(M3)和林缘(M4),于每个微生境中布设4个种子收集框,框口面积为100 cm×100 cm,框深20 cm[24]。由于香果树种子小,本研究采用网眼0.5 mm×0.5 mm的尼龙网制作收集框用于收集种子,收集框四周用4根1.3 m高的木棍撑起,使框底距地面1 m左右,防止小动物破坏,每天17:00收集一次框内种子,记录其数量,直至种子雨结束。待香果树母树种子雨结束后,将收集框内种子合并后进行记录。
4种微生境的选择:冠下位于母树树冠半径1/2处;冠缘位置位于母树树冠垂直投影的边缘;林窗位置在相邻乔木树冠垂直投影的中间位置(投影边缘的间距为2~5 m,林窗面积在4~20 m2之间);林缘则位于林地边缘(当香果树树冠与其一侧乔木树冠垂直投影间距均大于10 m时,可视为林缘),所选位置距香果树树冠垂直投影的距离在5 m以上,微生境中无灌木生存,仅有草本植物少量分布。所选4种微生境相互独立,互不影响。

表1 香果树种群的4个生境概况
注:林内光照、温湿度、土壤含水量等的测定于2014年7月进行,每样地连续观测3日,于每日10:00、13:00、15:00进行测定
Note: Measurement of in-forest light, temperature, humidity, soil water content, and other factors in July 2014. Data were continuously collected for three days at 10:00, 13:00 and 15:00 each day

表2 不同龄级香果树形态指标
注: S1. 20~50a; S2. 50~80a; S3. 80~100a; S4. 110~140a. 下同
Note: S1. 20~50a; S2. 50~80a; S3. 80~100a; S4. 110~140a. The same as below
2.3 香果树种子萌发实验及幼苗数量调查
2014年11月于研究种子质量及种子萌发的香果树母树上采集成熟果实,在每株母树树冠的上、中层分别选取20个果实;由于下层果实普遍较少,故在下层随机选择10个果实,每株母树共采集50个果实。自然晾干后,用游标卡尺测定种子的大小,并将所有种子进行分类,记录每类种子的数量。
从不同年龄段的香果树所产的种子中,选取饱满种子各20 000粒,在距开花母树200 m外的未结实香果树周围选择其冠下、冠缘、林窗以及林缘空地4种微生境的样方各10个,面积为200 cm×200 cm,将饱满种子均匀播撒于样方中,每样方500粒,用于次年6月、8月和10月份种子萌发及幼苗数量变化研究。
2.4 种子库收集
于2015年1月份,在与收集框相邻区域分别设置2个50 cm×50 cm的土壤种子库样方,用于1月、3月2次对香果树种子进行观测。每个土壤种子库样方分别选取枯落物和苔藓层、0~1 cm土壤层、1~5 cm土壤层等3个层位进行观察。采用网筛法分离土壤种子库中的香果树种子,并记录不同层筛选出的饱满(健康、完好的种子)、干瘪(种子已发育,扁平无活力)、虫蛀(虫啃食种子的子叶以及胚)以及霉烂种子数量。
2.5 数据处理
统计分析采用数据分析软件SPSS19.0,利用非参数Kolmogorov-Smirnov test与Lilliefors检验并校正单样本数据的正态性。采用单因素方差分析的方法确定数据间的差异性,利用LSD对数据两两间的差异做进一步显著性检验,显著水平为0.05。文中图采用软件Origin9.0制作。
3 结果与分析
3.1 香果树种子特征及组成
2014年11月,将宝天曼自然保护区采集的香果树果实以母树为一个单位收集种子,测量种子的形态及千粒重,将所得种子分为饱满、干瘪、空粒以及虫蛀等4类,计算其4类种子在总种子中所占的比率,结果见表3。由表3知,不同龄级香果树的种子特征存在显著差异(P<0.05),其中S1龄级母树产生的种子大小(长、宽和厚度)、千粒重以及饱满率均显著小于其他龄级母树,S1龄级母树所产种子大小仅为高龄级的一半左右,而千粒重也仅为0.28 g,其他龄级均在0.50 g左右。种子形态、千粒重以及饱满率在S2、S3和S4之间无差异。4个龄级香果树母树的种子干瘪率、空粒以及虫蛀率差异达显著水平。与其他龄级母树的种子相比较,S1龄级母树的种子干瘪率、空粒率较高(表3)。
3.2 不同龄级香果树种子雨特征
3.2.1 种子雨的时间动态 图1是宝天曼自然保护区不同龄级香果树的种子雨变化动态,其中y轴为不同龄级香果树母树微生境中种子雨密度的平均值。由图1知,宝天曼自然保护区香果树种子雨始于10月20日,终于12月1号,其持续时间约为40 d。不同龄级香果树的种子雨持续的时间有一定差异:随着年龄的增大,种子雨持续时间会延长,种子雨高峰期推迟。年龄最小的S1母树在10月30才开始种子飘落,而11月26日结束,种子雨持续时间最短,其种子下落量最大仅为4粒·m-2·d-1。经单因素方差分析知,4个年龄段香果树母树的种子雨密度存在极显著差异(P<0.01)。
3.2.2 种子雨的强度分布 图2为不同龄级香果树各不同微生境中种子雨的强度格局。由图2知,不同微生境中4个龄级香果树母树的种子雨强度变化趋势基本一致,由母树冠下至林缘,种子雨强度呈显著下降趋势,其中冠下和冠缘的种子雨强度差异不显著,但两者显著高于林窗的种子雨强度,林缘空地中种子雨的强度最低。经对4个龄级的香果树母树不同微生境的种子雨强度进行LSD多重比较分析知,S1母树的种子雨强度均小于其他年龄段母树种子雨强度,S2、S3和S4三个龄级不同微生境中种子雨强度之间差异未达显著水平。

表3 不同龄级香果树的种子特征
注:同列不同字母表示数据间在0.05水平差异显著
Note:The different letters marked on the same column of data show a significant difference at 0.05 level
3.3 香果树土壤种子库时空分布格局
3.3.1 土壤种子库储量及动态 图3是两次取样所得的不同龄级香果树种子库中不同类型种子的密度,经单因素方差分析知,1月和3月两次观测的香果树种子库中不同类型的种子密度之间差异显著(P<0.05);经LSD多重比较知,两次调查香果树土壤种子库中健康种子的密度与干瘪种子的密度之间差异不显著,两者显著小于虫蛀种子密度,霉烂种子密度最大,显著大于其他种子密度;经计算知,第2次调查土壤种子库总种子数量为第1次调查的42.29%,即1月到3月,近6成种子消失(图3),可见香果树种子在种子库中损耗较大。

图1 不同龄级香果树种子雨动态变化(2014年)Fig.1 Changes of the seed rain of E. henryi with different age classes

M1.冠下,位于母树树冠半径1/2处;M2.冠缘,位置位于母树树冠垂直投影的边缘;M3.林窗,位置在相邻乔木树冠垂直投影的中间位置(投影边缘的间距为2~5 m,林窗面积在4~20 m2之间);M4.林缘,位于林地边缘(当香果树树冠与其一侧乔木树冠垂直投影间距均大于10 m时,可视为林缘)。下同图2 不同龄级香果树母树周围种子雨强度格局M1. Under the tree crown,located at one half of the canopy radius; M2. Edge of the tree crown, located on the edge of vertical projection of the tree canopy;M3. Forest gap, located in the middle of the vertical projection of the adjacent tree canopies (the spacing of projection edge is between 2 m and 5 m, and the area of the forest gap is between 4 m2 and 20 m2); M4. Forest edge, located on the edge of the woodland (when the distance of the projection between the tree canopy and one side of the tree canopy is larger than 10 m, this place can be regarded as “Forest edge”). The same as belowFig.2 Seed rain intensity around the E. henryi mother trees with different age classes

同一龄级香果树种子的密度值标注有不同的字母,表示密度值之间在0.05水平差异显著图3 香果树土壤种子库不同组分在两次取样中的数量变化Different letters marked on the density value of E. henryi seed at the same age class show a clear difference at 0.05 levelFig.3 Quantity variation of different components of E. henryi seed bank between two samplings
3.3.2 土壤种子库的垂直分布特征 图4显示两次调查中不同年龄段香果树土壤种子库的垂直分布。由图4可知,香果树种子大部分聚集于枯落物和苔藓层(第1次71.54%;第2次71.20%),其中枯落物和苔藓层中的种子以霉烂种子为多(第1次7.69·m-2;第2次4.56株·m-2),健康种子很少(2.81株·m-2;1.75株·m-2);而0~1 cm的土壤层中虫蛀和霉烂的种子密度均较大,饱满种子密度最小;1~5 cm土壤层中种子仅有霉烂以及虫蛀2类种子,并以霉烂种子密度最大。
3.3.3 不同微生境种子库动态 图5为不同微生境香果树种子库中各类种子的百分比,且两次调查不同微生境土壤种子库特征存在一定差异。第1次取样种子库中以霉烂种子百分比最高,其次为虫蛀种子,饱满种子百分比最少;冠下、冠缘和林窗3种微生境土壤种子库中饱满种子百分比差异不显著,林缘空地中仅在S2和S3年龄段土壤种子库中发现饱满种子,其余两个年龄段未发现饱满种子。第二次取样香果树土壤种子库中虫蛀种子的百分比有所增加,但仍以霉烂种子为最多,饱满种子百分比仍最少;经单因素方差分析,不同龄级香果树土壤种子库中各类种子的百分比之间差异不显著(P>0.05)。
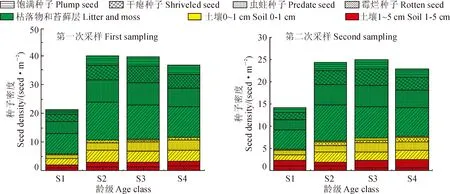
图4 不同龄级香果树土壤种子库垂直分布Fig.4 Horizontal distribution and vertical distribution of soil seed bank of E. henryi with different age classes

图5 不同微生境香果树土壤种子库特征Fig.5 Characteristics of soil seed bank of E. henryi with different microsites

表4 不同龄级香果树所产种子萌发及幼苗数量变化
注:同期同行不同字母表示微生境间在0.05水平有显著性差异
Note:The different normal letters in the same row during same stage indicate significant difference among microsites at 0.05 level
3.4 香果树种子萌发及幼苗数量
野外调查发现原生境中香果树的种子萌发较晚,4月底才出现露白,5月中下旬达到高峰,随后萌发数下降,至6月上旬结束。表4为6月、8月和10月3次调查的香果树实生苗数量变化。由表4知,香果树种子萌发率较低,为16.93%,8月份幼苗成活率仅占总幼苗数量的8.77%,10月份仅有3.86%的幼苗存活下来。不同微生境中香果树幼苗的数量均存在极显著差异(P<0.01),林窗中的香果树种子萌发及幼苗数量显著高于其他3个微生境。6月份冠下和林缘幼苗数量存在显著差异,到10月份差异不显著。经对香果树幼苗数量进行单因素方差分析,结果显示不同龄级香果树所产种子形成的幼苗数量之间无显著差异(P>0.05)。
4 讨 论
4.1 香果树的种子雨特征及其种子质量
不同植物物种的种子雨持续时间均有所不同,一些草本植物种子雨持续时间可长达5~7个月,如猪毛蒿(Artemisiascoparia)、铁杆蒿(Artemisiagmelinii)、阿尔泰狗哇花(Heteropappusaltaicus)以及灌木植物达乌里胡枝子(Lespedezadavurica)和草木樨状黄芪(Astragalusmelilotoides)[25],而乔木植物较短,如松科植物的种子雨持续时间为3个月左右[9]。宝天曼自然保护区香果树种子雨持续时间短,仅为40 d左右,这可能由其自身属性及环境所决定。宝天曼自然保护区位于河南省内乡县夏馆镇,该区10月份日最低气温已降至10 ℃以下,香果树尽管每花枝开花数量较多,但该物种具有开花不整齐的特点[13],香果树大部分果实在10月份因气温、土温的下降导致树液流动变慢而缺乏营养,导致其停止生长,未发育出种子或种子无活力,致使其每花枝最多有3个果实进入成熟期。该区分布的香果树的果实成熟期阴雨天较少,此阶段发育成熟果实的果皮迅速失水,干燥且外翻,果实沿中缝开裂,种子自然下落,但由于成熟的果实较少,导致其种子雨的持续时间较短。许多珍稀濒危植物的种子存在饱满率较低的特征,如毛红椿(Toonaciliatavar.pubescens)在12%~44%之间[26],秦岭冷杉(Abieschensiensis)为25.7%[27],大别山五针松(Pinusdabeshanensis)8%左右[28],而元宝山冷杉(Abiesyuanbaoshanensis)仅有6%[29]。香果树种子饱满率也略较低,在20%~40%之间,其余种子干瘪或被虫蛀,这可能是濒危植物的共性。
本研究发现香果树的种子雨与其自身的年龄有关,20~50龄级的香果树种子千粒重仅为0.28 g,其产种量较少,种子较小,且空粒较多,从而其种子雨持续时间最短,种子下落量最少,而较高龄级的香果树千粒重约为0.5 g,产种量较多,其种子雨持续时间较长。张希彪等[30]研究认为40 a油松林(Pinustabulaeformis)种子雨密度最大,低龄组和高龄组均较低,本研究发现高龄级香果树(110~140 a)的种子雨种子密度没表现出低于中龄级(50~110 a),这表明140 a左右的香果树还处于生殖旺盛期,未进入生理衰老期。香果树母树的年龄未对不同微生境中的种子雨强度产生显著的影响,而不同微生境之间的香果树种子雨强度存在显著差异,母树树冠下和树冠边缘较高,林窗中显著低于冠下和冠缘,而林缘空地上的种子雨强度显著低于林窗中的种子雨强度,这主要是由于距母树的距离所导致,距离越远其种子雨强度越低[31]。
4.2 香果树种子库种子数量的变化及其分布特征
结合宝天曼自然保护区香果树种子雨强度,第一次种子库调查结果显示,有41.01%的种子消失,而随后的2个月里,随着温度的上升,昆虫、病菌等的影响,剩余种子的57.71%又相继消失,种子库中种子的密度为21.69粒·m-2。有研究表明落于枯落物上的短命种子非常危险,因为它们无法与土壤接触,而被枯落物掩埋,或被苔藓挤占生存空间[32]。两次种子库调查分别有71.54%和71.20%的种子位于枯落物和苔藓层,土壤表层(0~5 cm)处的种子密度较低,而大于5 cm土壤深度无香果树种子分布。香果树种子寿命为10个月左右[33],研究发现6月中下旬未见其有新的种子萌发,这可能表明其野外条件下,种子在6月中下旬或已失去生命力,故自11月形成种子库到次年的6月之间种子的命运对其自然更新具有关键作用。
宝天曼自然保护区香果树的土壤种子库不同类型种子的密度存在显著差异,其中饱满种子和干瘪种子的密度较小,显著低于虫蛀种子的密度,而霉烂种子的密度最大。香果树母树的年龄未对其土壤种子库分布产生显著影响,不同龄级香果树的种子百分比在同一微生境中数值相近。尽管冬季的宝天曼自然保护区降雨较少,但香果树主要位于溪流边,从而其生境比较潮湿,增大了种子霉烂的可能性[17],而大部分种子位于枯落物和苔藓层,也增大了被地上动物捕食的危险。最终香果树饱满种子密度仅为1~2粒· m-2,这严重制约了其自然更新。
4.3 香果树种群更新及建议
地被物对植物的种子萌发存在显著的影响,陈迪马等[34]认为枯落物有利于云杉的自然更新,可促进其种子萌发,但也有研究表明枯落物遮挡了阳光,不利于种子萌发[35]。本研究支持后者,香果树为光敏种子[15],约71%的种子落入枯落物和苔藓层中,由于香果树的叶子较大,伴随种子一起飘落,大部分种子被覆盖,使其种子无法见光,导致其种子尽管吸胀但还是不能萌发,而位于枯落物和苔藓层表面的种子,即使萌发,由于其千粒重较小(0.2~0.5 g),大部分萌发种子的胚根未穿过枯落物和苔藓层,而耗尽营养死亡,即枯落物和苔藓层阻碍了种子雨与土壤接触,对种子萌发产生机械障碍[36]。
不同微生境对香果树幼苗成活率有着显著影响,这可能是由于母树冠下枯落物较多,光照较弱,不利于其幼苗的形态建成。由于香果树幼苗得不到充足的光照,其胚根受枯落物的阻隔无法插入土壤,从而无法从土壤中获得营养,导致幼苗大量死亡。本研究发现,林窗为其幼苗生存的最佳环境,林窗中枯落物较少,光照条件较好,有利于其幼苗的生长。有研究表明强光和弱光对香果树幼苗的生长均产生抑制作用,2 000 Lx是其最适宜生长的光照强度,而林缘空地中枯落物和苔藓很少,尽管有少量草本遮盖,但光照很强,野外调查发现其光照有时可达90 000 Lx,这可能会严重抑制香果树幼苗的生长[18],加之光照强度增加,使得幼苗蒸腾速率增大以及土壤表层缺水,导致幼苗脱水而死亡。
综上,由于香果树土壤种子库在最初的4个月里损失了约有80%的种子,故可以在种子雨前采集香果树的种子,次年4月份进行播种,以减少种子在土壤种子库中的损耗。鉴于林窗中的香果树种子萌发率及幼苗存活率较高,需加强对林窗的保护。对密度过大的林地,需进行间伐,人为制造林窗;对于冠下和冠缘两种微生境,建议清理其地被物,以增加香果树种子与土壤接触的机会,改善幼苗的光照环境;针对林缘生境,则需增加地上植被的盖度,以遮挡过强的光线,减缓土壤水分和温度的过度增减,保持空气湿度,即抑制环境因子的剧烈变化,努力改善环境因子,从而有利于香果树幼苗的存活[35]。
[1] AND H F H, SMALLWOOD J. Ecology of seed dispersal[J].AnnualReviewofEcology&Systematics, 2003,13(4):201-228.
[2] 杨允菲, 祝 玲. 松嫩平原碱化草甸朝鲜碱茅种子散布机制的分析[J]. 植物学报, 1995,37(3):222-230.
YANG Y F, ZHU L. Analysis on the mechanism of seed dispersal ofPuccinelliachinampoensison alkalized meadow in Songnen Plain of China[J].ActaBotanicaSinica, 1995,37(3):222-230.
[3] PARKER V T, KELLY V R. Ecology of Soil Seed Bank[M]. San Diego: Academic Press, 1989: 231-255.
[4] ANGELA T M, DONALD R D. Potential contributions of the seed rain and seed bank to regeneration of native forest underplantation pine in New Zealand[J].NewZealandJournalofBotany, 1999,37(1):83-93.
[5] 杨跃军, 孙向阳, 王保平. 森林土壤种子库与天然更新[J]. 应用生态学报, 2001,12(2):304-308.
YANG Y J, SUN X Y, WANG B P. Forest soil seed bank and natural regeneration[J].ChineseJournalofAppliedEcology,2001,12(12): 304-308.
[6] HALL J B, SWAINE M D. Seed stocks in Ghanainan forest soils[J].Biotropica, 1980,12(4): 256-268.
[7] PARISH J A D,BAZZAZ F A. Ontogenetic niche shifts in old-field annuals[J].Ecology, 1985,66(4): 1 296-1 302.
[8] 杨 旭, 杨志玲, 雷 虓,等. 濒危植物凹叶厚朴幼苗更新及环境解释[J]. 林业科学, 2013,49(12):36-42.
YANG X, YANG Z L, LEI X,etal. Characteristics and environmental interpretation of seedling regeneration in the endangered speciesMagnoliaofficinalissubsp.biloba[J].ScientiaSilvaeSinicae, 2013,49(12): 36-42.
[9] 高润梅, 石晓东, 郭跃东,等. 文峪河上游华北落叶松林的种子雨、种子库与幼苗更新[J]. 生态学报, 2015,35(11):3 588-3 597.
GAO R M, SHI X D, GUO Y D,etal. Seed rain,soil seed bank and regeneration ofLarixprincipis-rupprechtiistands in the Upper Reaches of Wenyuhe[J].ActaEcologicaSinica, 2015,35(11): 3 588-3 597.
[10] 李小双, 彭明春, 党承林. 植物自然更新研究进展[J]. 生态学杂志, 2007, 26(12):2 081-2 088.
LI X S, PENG M C, DANG C L. Research progress on natural regeneration of plants[J].ChineseJournalofEcology, 2007,26(9): 2 081-2 088.
[11] 张文辉, 祖元刚, 刘国彬. 十种濒危植物的种群生态学特征及致危因素分析[J]. 生态学报, 2002,22(9):1 512-1 520.
ZHANG W H, ZU Y G, LIU G B. Population ecological characteristics and analysis on endangered cause of ten endangered plant species[J].ActaEcologicaSinica, 2002,22(9): 1 512-1 520.
[12] 于永福. 中国野生植物保护工作的里程碑——《国家重点保护野生植物名录(第一批)》出台[J]. 植物杂志, 1999,5(3): 3-11.
YU Y F. Milestone in Chinese wild plant conservation—“National key protected wild plants (the first batch)” were issued[J].JournalofPlant, 1999,5(3): 3-11.
[13] 郭连金, 林国卫, 徐卫红,等. 武夷山香果树自然种群生殖构件特性研究[J]. 西北林学院学报, 2011,26(4):18-22.
GUO L J, LIN G W, XU W H,etal. Characteristics of reproductive modules ofEmmenopteryshenrysnatural population in Wuyi Mountain[J].JournalofNorthwestForestryUniversity, 2011,26(4): 18-22.
[14] 郭连金, 李 梅, 林 盛. 香果树种群开花物候、生殖构件特征及其影响因子分析[J]. 林业科学研究, 2015,28(6):788-796.
GUO L J, LI M, LIN S. Flowering phenology,reproductive module characteristics and their influencing factors of endangered plant speciesEmmenopteryshenryi[J].ForestResearch, 2015,28(6): 788-796.
[15] 李铁华, 周佑勋, 段小平,等. 香果树种子休眠和萌发的生理特性[J]. 中南林业科技大学学报, 2004,24(2):82-84.
LI T H ZHOU Y X, DUAN X P,etal. Physiological characteristics of the dormancy and light-sensitive germination ofEmmenopteryshenyiseeds[J].JournalofCentralSouthForestryUniversity, 2004,24(2): 82-84.
[16] 郭连金, 薛苹苹, 邵兴华,等. 香果树根萌苗生长特性及影响因子分析[J]. 植物科学学报, 2015,33(2):165-175.
GUO L J, XUE P P, SHAO X H,etal. Growth characteristics and influencing factors ofEmmenopteryshenryiroot sprouts[J].PlantScienceJournal, 2015,33(2): 165-175.
[17] 郭连金. 濒危植物香果树幼苗空间格局及数量动态研究[J]. 西北植物学报, 2014, 34(9):1 887-1 893.
GUO L J. Spatial distribution pattern and number dynamics ofEmmenopteryshenryiseedlings endangered in China[J].ActaBot.Boreal.-Occident.Sin., 2014,34(9): 1 887-1 893.
[18] 刘 鹏, 康华靖, 张志详,等. 香果树(Emmenopteryshenryi)幼苗生长特性和叶绿素荧光对不同光强的响应[J]. 生态学报, 2008,28(11):5 656-5 664.
LIU P, KANG H J, ZHANG Z X,etal. Responses of growth and chlorophyll florescence ofEmmenopteryshenryiseedlings to different light intensities[J].ActaEcologicaSinica, 2008,28(11): 5 656-5 664.
[19] 李中岳, 班 青. 香果树的生物学特性与繁殖方法[J]. 林业科技开发, 1995,(4):37-38.
LI Z Y, BAN Q. Biological characteristics and breeding methods ofEmmenopteryshenryi[J].ScienceandTechnologyofForestry, 1995, (4): 37-38.
[20] 郭连金. 濒危植物香果树(Emmenopteryshenryi)种群结构与动态[J]. 植物科学学报, 2009,27(5):509-514.
GUO L J. Population structure and dynamics of the endangered plantEmmenopteryshenryi[J].JournalofWuhanBotanicalResearch, 2009,27(5): 509-514.
[21] 郭连金, 李 梅. 濒危植物香果树武夷山种群分布格局的分形分析[J]. 西北植物学报, 2009,29(5):1 033-1 039.
GUO L J, LI M. Fractal analysis of the distribution pattern of an endangered plant,Emmenopteryshenryipopulation in Wuyi Mountain[J].ActaBot.Boreal.-Occident.Sin., 2009,29(5): 1 033-1 039.
[22] 甘 聃, 陈发菊, 梁宏伟,等. 珍稀濒危植物香果树种子萌发特性研究[J]. 种子, 2006, 25(5):27-29.
GAN D, CHEN F J, LIANG H W. Study on the characteristics of seed germination of the endangered plantEmmenopteryshenryi[J].Seed, 2006,25(5): 27-29, 34.
[23] 朱从波, 王万里, 刘晓静,等. 宝天曼自然保护区珍稀濒危植物研究[J]. 安徽农业科学, 2011,39(16):9 467-9 470.
ZHU C B, WANG W L, LIU X J,etal. Study on rare and endangered plants in Baotianman Nature Reserve[J].JournalofAnhuiAgri.Sci., 2011,39(16): 9 467- 9 470.
[24] 刘足根, 朱教君, 袁小兰,等. 辽东山区长白落叶松(Larixolgensis)种子雨和种子库[J]. 生态学报, 2007, 27(2):579-587.
LIU Z G, ZHU J J, YUAN X L,etal. On seed rain and soil seed bank ofLarixolgensisin montane regions of eastern Liaoning Province, China[J].ActaEcologicaSinica, 2007,27(2): 579-587.
[25] 于卫洁, 陈 宇, 焦菊英,等. 黄土丘陵沟壑区撂荒坡面种子雨特征[J]. 应用生态学报, 2015, 26(2):395-403.
YU W J, CHEN Y, JIAO J Y,etal. Characteristics of seed rain on abandoned slopes in the hilly-gullied Loess Plateau,North-West China[J].ChineseJournalofAppliedEcology, 2015,26(2): 395-403.
[26] 黄红兰. 九连山自然保护区毛红椿天然种群生态学特征[D]. 南昌: 江西农业大学, 2012.
[27] 李庆梅. 秦巴山地两种冷杉种实特性研究与秦岭冷杉濒危原因探讨[D]. 北京: 北京林业大学, 2008.
[28] 项小燕, 张小平, 段仁燕,等. 濒危植物大别山五针松母树林花粉传播规律[J]. 广西植物, 2014,34(3):333-337.
XIANG X Y, ZHANG X P, DUAN R Y,etal. Pollen dispersal patterns of the endangered plantPinusdabeshanensisin a seed production stand[J].Guihaia, 2014,34(3): 333-337.
[29] 黄仕训. 元宝山冷杉濒危原因初探[J]. 生态与农村环境学报, 1998,14(1):6-9.
HUANG S X. Preliminary study on the causes of the endangerment ofAbiesyuanbaoshanensis[J].RuralEco-Environment, 1998,14(1): 6-9.
[30] 张希彪, 王瑞娟, 上官周平. 黄土高原子午岭油松林的种子雨和土壤种子库动态[J]. 生态学报, 2009, 29(4):1 877-1 884.
ZHANG X B, WANG R J, SHANGGUAN Z P. Dynamics of seed rain and soil seed bank inPinustabulaeformisCarr. forests in eroded hilly loess regions of the Loess Plateau in China[J].ActaEcologicaSinica, 2009,29(4): 1 877-1 884.
[31] 岳红娟, 仝 川, 朱锦懋,等. 濒危植物南方红豆杉种子雨和土壤种子库特征[J]. 生态学报, 2010,30(16):4 389-4 400.
YUE H J, TONG C, ZHU J M,etal. Seed rain and soil seed bank of endangeredTaxuschinensisvar.maireiin Fujian, China[J].ActaEcologicaSinica, 2010,30(16): 4 389-4 400.
[32] DONATH T W, ECKSTEIN R L. Effects of bryophytes and grass litter on seedling emergence vary by vertical seed position and seed size[J].PlantEcology, 2010, 207(2):257-268.
[33] 陈 黎, 周 凯. 香果树种子贮藏寿命的研究[J]. 黄山学院学报, 2007, 9(5):64-66.
CHEN L, ZHOU K. Study on the storage time ofEmmenopteryshenryiOliv. seeds[J].JournalofHuangshanUniversity, 2008,9(5): 64-66.
[34] 陈迪马, 潘存德, 刘翠玲,等. 影响天山云杉天然更新与幼苗存活的微生境变量分析[J]. 新疆农业大学学报, 2005, 28(3):35-39.
CHEN D M, PAN C D, LIU C L,etal. Analysis on micro-habitat variables affecting natural regeneration and survival of seedlings inPiceaschrenkianastand[J].JournalofXinjiangAgriculturalUniversity, 2006,28(3): 35-39.
[35] 胡 蓉, 林 波, 刘 庆. 林窗与凋落物对人工云杉林早期更新的影响[J]. 林业科学, 2011,47(6):23-29.
HU R, LIN B, LIU Q. Effects of forest gaps and litter on the early regeneration ofPiceaasperataplantations[J].ScientiaSilvaeSinicae, 2011,47(6): 23-29.
[36] NAVARRO-CANO J A. Effect of grass litter on seedling recruitment of the critically endangeredCistusheterophyllusin Spain[J].FloraMorphologyDistributionFunctionalEcologyofPlants, 2008,203(8):663-668.
(编辑:潘新社)
Seed Rain, Seed Bank and Natural Regeneration ofEmmenopteryshenryiin Different Age Classes
GUO Lianjin1, XU Weihong1, FANG Huipu2, XIAO Zhipeng1,TIAN Yuqing1,3, HUANG Xiahua1, YIN Chongmin1, HU Genxiu1
(1 Shangrao Normal University, Shangrao, Jiangxi 334000, China; 2 Administrative Bureau of Baotianman National Nature Reserve,Nanyang, Henan 474350, China; 3 Kunming Institute of Botany, Chinese Academy of Sciences, Kunming 650204, China)
Emmenopteryshenryiseed rain and soil seed bank in Baotianman Nature Reserve were observed. Seeding experiments at different microhabitats were conducted, and the dynamics of seed germination and seedling growth were investigated. The results showed that: (1) there are differences in time duration and peak period ofE.henryiseed rain at different age classes. Obvious differences in seed rain intensity were also observed. (2)E.henryiseeds of 20-50 age class exhibit an obviously lower satiation rate than that of other age classes. The seed density of theE.henryiseed bank is obviously lower than that of other age classes. However, different types of seeds exhibit uniform horizontal and vertical distribution rules. Approximately 80% ofE.henryiseeds disappear before germination. Rotten seeds exhibit the highest density (9.81 particles·m-2), whereas fully grown seeds exhibit the lowest density (1.94 particles·m-2). (3) Outdoor experiment of growing seedlings shows no obvious differences in germination and seedling survival rates of fully grown seeds at different age classes.E.henryigermination rate is approximately 16.93%. However, only approximately 3.86% of seedlings exhibit more than five months of life. Different microhabitats show a clear influence on the survival rate. Forest gap is the most suitable microhabitat. This study demonstrates gathering and seeding ofE.henryiseeds artificially at the original habitat in the next April, cleaning ground covers under female crown and edge, increasing coverage of edge space to reduce loss, and improving light conditions for germination and growth to benefit the growth of the seedlings root system and promote its natural regeneration.
Emmenopteryshenryi; seed quality; seed rain; seed bank; seedling regeneration
1000-4025(2016)11-2273-10
10.7606/j.issn.1000-4025.2016.11.2273
2016-09-02;修改稿收到日期:2016-10-08
国家自然基金项目(31360145);江西省教育厅科技计划项目(GJJ08463)
郭连金(1976-),男,副教授,主要从事植物生态学与生物多样性研究。E-mail: guolianjin@163.com
Q948.1
A
猜你喜欢
湖南生态科学学报(2022年4期)2022-11-11
农业科技与信息(2022年12期)2022-08-09
山东林业科技(2021年4期)2021-08-31
落叶果树(2021年6期)2021-02-12
现代园艺(2020年17期)2020-08-31
河南农业大学学报(2020年2期)2020-05-22
山西林业科技(2019年1期)2019-05-27
现代园艺(2018年1期)2018-03-15
体育科技(2016年2期)2016-02-28
浙江大学学报(农业与生命科学版)(2015年4期)2015-12-15
